
Abstract
Carbapenems are classified as critically important antibiotics since they are employed when resistant Gram-negative bacterial infections fail to respond to other antibiotic therapies. Carbapenem-resistant bacteria (CRB) were traditionally understood to be rare in the U.S. food-producing animals. Recently, using quantitative polymerase chain reaction (qPCR), our group detected bla KPC-2 in all 72 metagenomic DNA (mgDNA) samples prepared from the feces of 36 lots of beef cattle “raised without antibiotics” (RWA) and 36 lots raised “conventionally” (CONV). Since a small internal fragment of the bla KPC-2 gene was targeted by the qPCR detection method, we sought to determine if functional bla KPC-2-like sequences are present in beef cattle feces. Full-length bla KPC-2 sequences were amplified from 18 mgDNA samples (9 CONV and 9 RWA), cloned into pCR4Blunt-TOPO vectors, and transformed into Escherichia coli TOP10 cells. All 14 of the samples with bla KPC-2 cloned in the same orientation as the Plac promoter had carbapenemase activity and imipenem minimum inhibitory concentrations ≥32 μg/mL. We conclude that the bla KPC-2 genes detected in our previous study were functional, which indicates that CRB were present in those fecal samples. Identification of functional Klebsiella pneumoniae carbapenemases in fecal samples from both CONV and RWA cattle strongly suggests that CRB are more common in U.S. beef cattle feces than previously believed. Critically, more research using similar qPCR methods to determine the levels of carbapenem-resistant genes in human feces, feces from other food animal species, wildlife, companion animals, and the environment are required to accurately assess public health implications.
Introduction
C
Klebsiella pneumoniae carbapenemases (KPC) are class A serine β-lactamases (encoded by the bla KPC gene) that can hydrolyze a wide variety of β-lactam antimicrobials, including carbapenems, cephalosporins, cephamycins, and penicillins (Queenan and Bush, 2007; Papp-Wallace et al., 2010; Munoz-Price et al., 2013). Since identification in 1996, CRB-harboring bla KPC have been isolated from human clinical samples worldwide (Munoz-Price et al., 2013). We recently reported the results of a study that used multiple methods to compare the occurrence of antimicrobial resistance in feces of beef cattle “raised without antibiotics” (RWA) and raised “conventionally” (CONV) (Vikram et al., 2017). The bla KPC-2 gene was detected in all 72 samples of metagenomic DNA (mgDNA) tested by quantitative polymerase chain reaction (qPCR). These 72 samples (36 CONV and 36 RWA) each represented a “lot” (group of animals produced and marketed together) and were equally distributed over a calendar year. Since in the previous study, a small internal fragment of the bla KPC-2 gene was targeted (Fig. 1) for qPCR, this report describes the additional experiments necessary to confirm whether functional KPC carbapenemases were present in those beef cattle fecal samples.
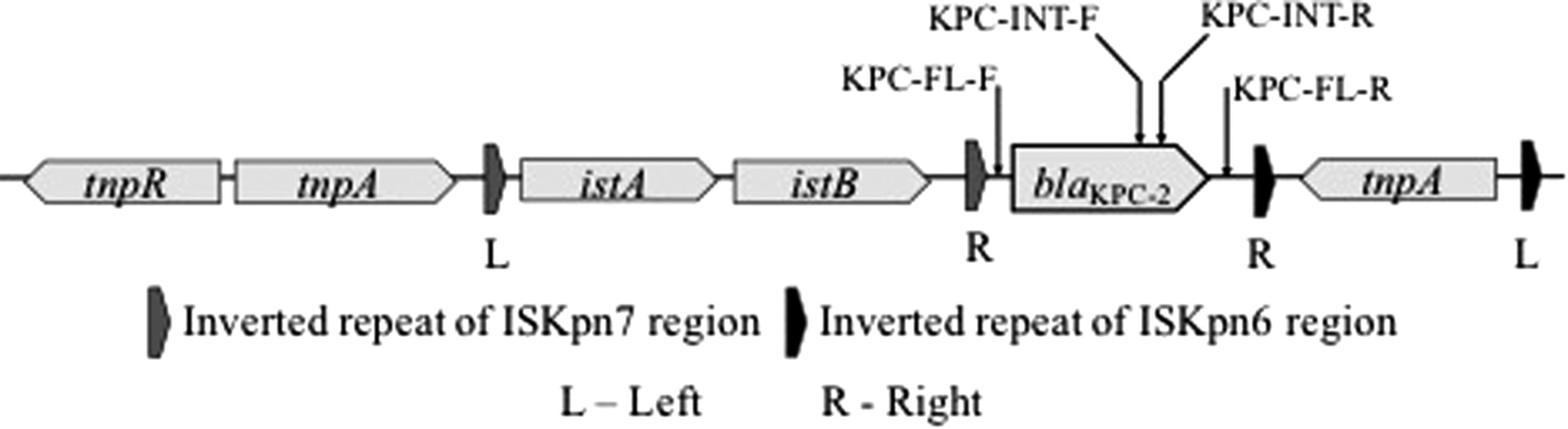
Genetic organization of canonical Tn4401 carrying bla KPC-2. Oligonucleotide primer annealing locations are indicated by black arrows. KPC, Klebsiella pneumoniae carbapenemases
Methods
Institutional approval
All procedures and protocols used in this study were reviewed and approved by the U.S. Meat Animal Research Center Institutional Biosafety Committee (USMARC IBC Protocol #3.0). All fecal samples were obtained from eviscerated colons produced during the normal and routine operations at a commercial beef processing plant operating in compliance with all applicable regulations and inspected by the U.S. Department of Agriculture, Food Safety and Inspection Service. The samples were collected monthly at the processing plant from February 2014 to January 2015. For each month 10 samples per lot for six lots (three RWA, three CONV) representing six production sites were collected. Samples were transported to the laboratory on ice packs and held at −20°C until DNA isolation.
bla KPC-2 cloning
mgDNA isolation was described previously (Vikram et al., 2017). Briefly, each mgDNA sample consisted of mgDNA isolated from feces obtained from 10 colons from different animals within a lot. Cattle within a lot were usually housed together at a feeding operation in the same pen or groups of pens. To design oligonucleotide primer pairs KPC-INT-F/KPC-INT-R and KPC-FL-F/KPC-FL-R 21 bla KPC-2 sequences from multiple species were downloaded in Geneious 9.1.6 (Biomatters Ltd., Newark NJ) and aligned. Primers KPC-INT-F and KPC-INT-R were designed in the conserved region within the bla KPC-2 gene, whereas KPC-FL-F and KPC-FL-R were constructed to anneal to the conserved spacer region between the Tn3 repeat region and the 3′ end of the bla KPC-2, and the 5′ end of the bla KPC-2 gene and tnpA (Fig. 1). Oligonucleotide primers KPC-FL-F and KPC-FL-R (Table 1) were used to amplify full-length bla KPC-2 from 18 mgDNA samples (9 CONV and 9 RWA). These samples were selected from RWA and CONV lots and were positive for bla KPC-2 gene by qPCR. A Bio-Rad T100 thermal cycler (Bio-Rad Laboratories, Inc., Hercules, CA) was used with 0.2 μM (final concentration) of primers, and Platinum Pfx DNA polymerase (Invitrogen Life Technologies, Carlsbad, CA). The cycling conditions were 94°C for 3 min, followed by 27 cycles of 94°C for 30 s, 55°C for 30 s, and 68°C for 90 s with final extension on 68°C for 10 min. For each sample, a single band of the expected size (∼1.1 kbp) was observed when PCR products were separated by electrophoresis on a 1.2% agarose gel. The band was excised and purified using the QIAquick Gel Extraction Kit (Qiagen Inc. Valencia, CA). The purified DNA was inserted into pCR4Blunt-TOPO cloning vector (Invitrogen Life Technologies) and transformed into Escherichia coli TOP10 cells (Invitrogen Life Technologies) plated on Luria-Bertani agar plates supplemented with 50 μg/mL kanamycin according to the manufacturer's instructions. E. coli transformants with reduced carbapenem susceptibility were identified by streaking on LB agar media supplemented with 1.25 μg/mL of imipenem. For each transformation event, between 5 and 10 colonies with reduced carbapenem susceptibility were screened for the presence of cloned fragments by PCR using M13 primers specific to sequences that flank the pCR4Blunt-TOPO cloning site. Then transformants with cloned sequences were screened for the presence or absence of bla KPC-2 by PCR with the oligonucleotide primers KPC-INT-F and KPC-INT-R (Table 1). A Bio-Rad T100 thermal cycler was used with 0.2 μM (final concentration) of primers, 0.3 μL Bullseye Hot Start Taq DNA polymerase (MidSci, St. Louis, MO), and dNTPs (0.4 μL). Thermal cycling conditions were 96°C for 3 min followed by 34 cycles of 96°C for 30 s, 55°C for 30 s, and 72°C for 1 min with final extension on 72°C for 5 min. The transformants were stored as glycerol stocks and for ease of reading are referred to as “strains” hereafter.
KPC, Klebsiella pneumoniae carbapenemases
Imipenem minimum inhibitory concentrations
Imipenem minimum inhibitory concentrations (MICs) were determined using the broth dilution method. Briefly, each strain was restreaked on Mueller Hinton agar supplemented with 1.25 μg/mL imipenem and grown overnight at 37°C. Four colonies from each plate were resuspended in Mueller Hinton Broth (MHB) and OD625 was adjusted to 0.08–0.1 (equivalent to 0.5 McFarland solution). Fifty microliters of the OD625-adjusted culture was added to 96-well plates containing twofold dilutions of imipenem in MHB for final concentrations ranging from 0.25 to 256 μg/mL. Plates were incubated at 35°C for 16 to 18 h. Imipenem MIC interpretive criteria were: susceptible ≤1 μg/mL, reduced susceptibility = 2 μg/mL, resistant ≥4 μg/mL following Clinical and Laboratory Standards Institute (CLSI) guidance for imipenem breakpoints for Enterobacteriaceae (CLSI, 2016).
Carbapenemase activity
Carbapenemase activity was detected using the Neo-Rapid CARB Kit 98024 (Rosco Diagnostica, Denmark), according to the manufacturer's instructions. Briefly, 10 μL loop of a colony grown overnight was suspended in 200 μL 0.9% sodium chloride solution (pH 8.5) in a glass tube. Ten microliters of 2.14% (wt/vol) Triton X-100 solution was added, and the tube was vortexed for 30 s and incubated for 30 min at room temperature. Indicator tablets were added to each tube then vortexed briefly to dissolve the tablets. Tubes were incubated at 37°C and inspected at 15, 30, and 60 min for any color change compared with negative controls. Change in color (from red to yellow or orange) was considered as positive carbapenemase activity.
Analysis of cloned sequences conferring reduced imipenem susceptibility
Plasmids were isolated using the PureLink Quick Plasmid Miniprep Kit (Invitrogen Life Technologies) according to the manufacturer's instructions. The cloned region was Sanger sequenced using M13 primer pair and fluorescently labeled terminator dye on ABI 3730xI DNA Analyzers (Applied Biosystems, Carlsbad, CA) at GeneWiz (South Plainfield, NJ) facility. Forward and reverse reads were merged using BBmerge in BBtool plugin (
Results
For each of the 18 mgDNA samples, one bla
KPC-2 putative positive strain with reduced imipenem susceptibility was selected without any specific criteria (named A1 to A18) and characterized. Of the 18 bla
KPC-2 putative positive strains characterized, 14 had carbapenemase activity and imipenem MIC ≥32 μg/mL (Table 2). The remaining four strains had imipenem MICs = 2 μg/mL and lacked carbapenemase activity. Of the 18 bla
KPC-2-positive strains, 17 were sequenced with average Phred quality score (Q) of >30, where Q refers to the accuracy of nucleotide identification during automated sequencing (Ewing et al., 1998). Repeated sequencing for strain A13 in both directions resulted in poor-quality data. For each strain, the cloned sequences were highly similar to each other and the three bla
KPC-2 canonical examples (Fig. 2 and Supplementary Fig. S1; Supplementary Data are available online at

Generalized reversible time model tree for cloned bla KPC-2 fragments with three published sequences. The sequences were aligned on Geneious 9.1.6 using parameters 93% similarity, Gap open penalty 12 and Gap extension penalty 3 and tree was generated using general time-reversible model by MrBayes in Geneious 9.1.6. The numbers over branches denote the number of base substitutions per site.
MIC, minimum inhibitory concentration; n/a, not applicable since A13 was not sequenced with high quality.
For each sample, reduced imipenem susceptibility strains negative for bla KPC-2 by PCR with the oligonucleotide primers, KPC-INT-F and KPC-INT-R, were identified. Thirty-three of these strains were characterized to determine if these strains harbored non-bla KPC-2 carbapenemases. For the one strain (D3) with a 64 μg/mL imipenem MIC and carbapenemase activity, the cloned sequence was bla KPC-2 (Table 1). The other 32 strains had 2 μg/mL imipenem MICs and lacked carbapenemase activity. Sequence analysis showed these were similar to Bacteroides intestinalis hypothetical protein (Accession no. WP_061437718), Alistipes finegoldii gene encoding a tetraacyl disaccharide 4′ kinase LpxK protein (Accession no. CCZ76077), uncultured bacterium extrachromosomal DNA RGI00888 and Ruminococcus sp. (CDA20468) RecX protein.
Discussion
In this study, we have used a functional metagenomics approach to unambiguously demonstrate that a functional bla KPC-2 gene is present in the feces of market weight U.S. beef cattle. Furthermore, the successful bla KPC-2 cloning from all 18 samples with primers specific to transposon sequences (KPC-FL-F and KPC-FL-R) demonstrates that the detected bla KPC-2 genes may be mobilized. These results are significant because bla KPC-2 was not previously reported from food animals despite its widespread distribution in the United States. Interpreting these results is challenging since we are aware of only three analogous published studies that examined feces or fecal impacted environments for bla KPC-2 by qPCR, although with different primers and amplification conditions. Two studies of a municipal wastewater treatment plant in Tianjin, China reported that bla KPC-2 was detected in influent, all treatment phases, final effluent, and dewatered sludge (Yang et al., 2016, 2017). Another study reported that bla KPC-2 was detected in wastewater effluents from three of seven Tunisian hospitals (Nasri et al., 2017).
Previous studies have suggested that CRB are rare in U.S. food animals. Webb et al. (2016) recently examined 159 fecal samples from dairy cattle at U.S. farms and concluded that CRB are “rare” in U.S. dairy cattle. However, the selective culture methods used may have failed to detect poorly cultivated species. Moreover, none of the bacteria with reduced susceptibility to carbapenems isolated by Webb et al. harbored bla KPC-2. Also, Mollenkopf et al. (2017) used selective culture methods to isolate CRB from environmental samples obtained at a U.S. swine production operation. Multiple genera of CRB (Citrobacter, Enterobacter, Escherichia, Klebsiella, Morganella, Proteus, and Providencia) were isolated by Mollenkopf et al. and all CRB isolates harbored bla IMP-27.
In contrast, results from our studies (Vikram et al., 2017; and this study) led us to conclude that bla KPC-2 is likely more common in U.S. beef cattle than previously thought. This conclusion is supported by: (1) qPCR detection of bla KPC-2 in feces from 72 different lots of cattle distributed over one calendar year (Vikram et al., 2017); (2) successful cloning of bla KPC-2 from all 18 attempts using fecal mgDNA from different lots (Table 2); and (3) carbapenemase activity for all 14 strains with a bla KPC-2 cloned in the proper orientation for transcription from the Plac promoter (Table 2).
Additionally, we conclude that bla KPC-2 is present in U.S. cattle feces regardless of antimicrobial exposure. Carbapenems, a sub-class of β-lactam antimicrobials, are not used in U.S. agriculture. However, Mollenkopf et al. (2017) more frequently isolated CRB in barns with greater use of the cephalosporin β-lactam ceftiofur leading to speculation that ceftiofur use may contribute to increased occurrence of carbapenemase genes. Conversely, using qPCR we found that fecal bla KPC-2 levels did not differ between beef cattle produced CONV and RWA (Vikram et al., 2017) suggesting that factors other than antimicrobial use contribute to occurrence of bla KPC-2 and possibly other CRB.
This study, together with our previous study of AMR in beef cattle feces (Vikram et al., 2017), the Webb et al. (2016) and Mollenkopf et al. (2017) studies demonstrate that diverse CRB are present in U.S. food animal production environments. However, conclusions regarding public health impacts or the influence of antimicrobial use practices during U.S. food animal production are premature. We note that Agga et al. (2015) more frequently detected carbapenem resistance genes in U.S. municipal wastewater treatment plant effluents than in cattle feedlot runoff catchment ponds and swine waste lagoons. Additional research is required to provide an ecological understanding of carbapenemase resistance in human feces, feces from food animal species, companion animals, wildlife, and the environment. These surveys must simultaneously incorporate multiple culture methods (as this may facilitate the isolation and analysis of organisms harboring bla KPC-2) and qPCR for multiple carbapenemase genes. Metagenomic sequencing will not be sufficient. Using qPCR we detected bla KPC-2 in 72 mgDNA samples of beef cattle feces, but metagenomic sequencing did not detect bla KPC-2 (Vikram et al., 2017). Also, longitudinal cohort studies specific to each commodity are required to accurately gauge the impact of specific antimicrobial uses (i.e., ceftiofur) in food animal production on carbapenemase gene and CRB occurrence.
Footnotes
Acknowledgments
The authors thank Julie Dyer and Frank Reno for technical support. They thank Jody Gallagher for administrative assistance.
Names are necessary to report factually on available data; however, the USDA neither guarantees nor warrants the standard of the product, and the use of the name by the USDA implies no approval of the product to the exclusion of others that may also be suitable. USDA is an equal opportunity provider and employer.
Funding
This work was supported by the Untied States Department of Agriculture, Agricultural Research Service, National Program 108—Food Safety (Project No. 3040-42000-018). The funding agency had no role in study design, data collection, and interpretation, or the decision to submit the work for publication.
Disclosure Statement
No competing financial interests exist.
Author Contributions
A.V. and J.W.S. designed the research. A.V. performed the experiments. A.V. and J.W.S. analyzed data. A.V. and J.W.S. wrote the article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.