
Abstract
Salmonella serotypes linked to tomato-associated outbreaks were evaluated for survival in soil and water over a 40-day period. Salmonella enterica serotypes Anatum, Baildon, Braenderup, Montevideo, Newport, and Javiana were inoculated separately into sterile soil and water, followed by plating onto TSAYE and XLT4 at 10-day intervals. Biofilm production by Salmonella serotypes was measured on both quartz particles (soil surrogate) and glass coverslips, and was evaluated using a crystal violet dye assay. Salmonella populations in soil and water over 40 days indicated no significant differences between Salmonella serotypes tested (p > 0.05). Over a 40-day period, there was a 1.84 ± 0.22 log CFU/g and 1.56 ± 0.54 CFU/mL decrease in populations of Salmonella in soil and water, respectively. Enumeration indicated that Salmonella population fluctuated in water but decreased linearly in soil. All serotypes tested produced the “red dry and rough” morphotype on Congo Red agar. Biofilm produced by all the Salmonella serotypes tested was significantly different on quartz particles than on glass coverslips (p < 0.0001), indicating that material and surface characteristics could affect biofilm development. The ability of Salmonella serotypes to persist in soil or water and attach to abiotic surfaces through biofilm formation affirms that contact surfaces, soil, water, and sediment should be considered as possible sources of cross-contamination in the farm environment.
Introduction
F
Salmonella spp. are intracellular animal pathogens that can survive in fallow soil for a period of 6 weeks while retaining their ability to cross-contaminate the phyllosphere (Barak and Liang, 2008). Inoculation of tomato blossoms with Salmonella spp. has resulted in its presence in internal fruit tissue (Guo et al., 2001; Shi et al., 2007; Kumar et al., 2017). Since blossom-set to fruit formation in tomato can take 20–30 days, contamination of blossoms by Salmonella during this duration may result in fruit contamination. Animal feces, soil, water, dust, and insects could serve as potential vehicles of Salmonella, leading to blossom contamination (Cevallos-Cevallos et al., 2012; Kumar et al., 2017).
Persistence of Salmonella in pond water and sediments is a pertinent produce safety concern because of the risk of cross-contamination (Greene et al., 2008; Micallef et al., 2012). Foodborne pathogens such as Salmonella persist in challenging environments by forming biofilms (Kumar and Micallef, 2017). The exopolymeric substances (EPS) in Salmonella biofilms are composed of curli and cellulose and are produced by red dry and rough (rdar) morphotypes (Steenackers et al., 2012; Kumar and Micallef, 2017). Variations in the production of curli and cellulose among Salmonella isolates from produce have been observed (Solomon et al., 2005). The ability to form biofilms by Salmonella can also differ by serotype and the type of material the bacterium is attached to (Kumar and Micallef, 2017). Out of 151 Salmonella strains, only 19% were strong biofilm formers, indicating differences in density of biofilms produced by the pathogen could occur based on serotype (Agarwal et al., 2011). While studies assessing biofilm formation by Salmonella have been performed on food contact surfaces or polystyrene plates (Kumar and Micallef, 2017), the ability of serotypes to form biofilms on environmental matrices such as sediments requires better understanding. Salmonella can form biofilms on surfaces of gallstones and pebbles (Prouty et al., 2002), indicating that sediment and soil particles may also serve as a niche for Salmonella biofilms.
It was hypothesized that the survival of Salmonella in soil and water and its ability to form biofilms on abiotic surfaces could vary among Salmonella serotypes. The objective of this study was to compare populations and injury of Salmonella serotypes previously linked to tomato outbreaks in soil and water without the presence of competing microbiota over a 40-day duration. Biofilm formation on abiotic surfaces was also evaluated among the Salmonella serotypes over a 14-day period.
Materials and Methods
Bacterial strains
Stock cultures of Salmonella enterica subsp. enterica serotypes Anatum, Baildon, Braenderup, Javiana, Montevideo, and Newport were obtained from the culture collection at Virginia Polytechnic Institute and State University. Frozen stock cultures were thawed, and a loopful of culture was transferred to 25 mL of Brain Heart Infusion broth (BHIB; Becton Dickinson and Co., Sparks, MD) and incubated for 24 h at 37°C. Cultures were transferred into fresh BHIB and incubated at 37°C for 24 h in a shaking incubator three successive times before use.
Soil and water preparation
Soil was obtained from a tomato farm on the Eastern Shore of Virginia. The soil was passed through a no. 20 sieve (USA Standard Testing Sieve) to obtain a uniform grain size. Ten-gram soil portions were autoclaved (121°C for 12 min) to inactivate existing microbiota. The autoclaved soil was tested for the absence of viable bacteria by performing aerobic plate counts. Autoclaved deionized distilled (121°C for 12 min) water was used to study survival patterns of Salmonella serotypes in water.
Inoculum preparation
A loopful of each Salmonella serotype (Salmonella Anatum, Baildon, Braenderup, Javiana, Montevideo, and Newport) was used to inoculate separate tubes (25 mL) of tryptic soy broth (TSB). The broth was incubated at 37°C for 24 h in a shaking incubator at 100 rpm. After incubation, the culture was centrifuged at 1400 g for 7 min. The supernatant was decanted. The pellet was washed twice with sterile distilled deionized water and re-suspended in 5 mL of sterile distilled deionized water.
Inoculation of soil and estimation of Salmonella population
To ensure even spatial distribution of Salmonella in soil, 10 g portions of the autoclaved soil inoculated with 1 mL deionized distilled water suspension of Salmonella was added to a sterile sample collection container (Thermo Fisher Scientific, Waltham, MA) to obtain a final 50 g sample of inoculated soil. The soil samples were thoroughly mixed with a sterile stirring rod. Each 50 g sample of soil contained a single Salmonella serotype. The initial concentration of pathogen in soil was around 9 log CFU/g. The containers were incubated at 25°C over a period of 40 days, and 1 g of soil was sampled every 10 days from each container. Populations of Salmonella spp. in soil were estimated through spread plating on XLT4 and TSAYE, respectively (Tryptic Soy Agar supplemented with 0.6% yeast extract).
Inoculation of water and estimation of Salmonella population
Forty-five-milliliter portions of sterile distilled deionized water were inoculated individually with 5 mL of each Salmonella serotype and mixed well. The Salmonella in water suspensions were incubated at 25°C for 40 days, and 1 mL of the suspension was sampled at day 0 and every 10 days. Populations of Salmonella spp. were estimated through spread plating on XLT4 and TSAYE.
Curli and cellulose expression
Salmonella serotypes Anatum, Baildon, Braenderup, Javiana, Montevideo, and Newport were grown in TSB at 37°C for 18 h. A loopful of inoculum of each serotypes was streaked on separate Luria Bertani plates without salt, supplemented with Congo Red (40 μg/mL) and Coomassie brilliant blue (20 μg/mL; CR plates). Colony morphologies were assessed on CR plates after incubation at 28°C for 48 h for individual colonies with red dry and rough morphotype, which is indicative of curli and cellulose expression.
Growth of biofilm glass coverslips
Glass coverslips were placed in the bottom of a sterile polystyrene Petri dish and covered with 30 mL of TSB. Each Salmonella serotype was inoculated into plates containing coverslips using 1:100 dilutions in fresh TSB of overnight cultures to compare differences in attachment and biofilm formation between Salmonella serotypes. Plates were rocked slowly (30 rpm) overnight at 37°C. Every 24 h, the medium was removed, and the coverslips were thoroughly washed twice with 1 × phosphate-buffered saline (PBS) to remove easily detached cells. Coverslips were sampled to estimate biofilm formation on days 2, 6, 10, and 14.
Growth of biofilm quartz particles
Quartz particles were used as a surrogate for soil particles to facilitate crystal violet (CV) staining due to uniformity in color. Twenty grams of quartz (50–70 mesh; Sigma–Aldrich, St. Louis, MO) was placed in a sterile Petri dish containing 20 mL of TSB broth. Each individual Salmonella serotype was inoculated into plates containing quartz particles using 1:100 dilutions of overnight cultures to compare differences in attachment and biofilm formation between Salmonella serotype. The Petri dishes were placed in a shaking incubator at 30 rpm and allowed to incubate overnight. Every 24 h, the medium was removed, and the quartz particles were washed twice in 1 × PBS. Two hundred and fifty milligrams of the quartz particles were removed for biofilm estimation on days 2, 4, 6, 10, and 14.
CV staining glass coverslips
CV staining of sessile cells was adapted from the procedure used by Prouty et al. (2002). Coverslips were removed from the Petri dish and washed with a continuous spray of 1 × PBS until nutrient medium and easily detached cells were washed off. Biofilms were fixed by drying coverslips at 60°C for 1 h. Biofilms were stained by immersing coverslips in 0.1% CV (gentian violet in isopropanol-methanol-1 × PBS [1:1:18]) for 15 min at room temperature. The coverslip was then washed thoroughly with 1 × PBS until the PBS ran clear and then crushed and put into an Eppendorf tube containing 1 mL 33% acetic acid to extract the dye. The optical density at 570 nm (OD570) of the acetic acid solution was assessed to determine the amount of dye retained by attached bacterial cells.
CV staining quartz particles
Quartz particles were washed with a steady spray of 1 × PBS until all broth and debris was removed. Two hundred and fifty milligrams of the quartz particles were transferred to a 1.5 mL polypropylene Eppendorf tube. Quantification of biofilm was performed by staining heat-fixed quartz particles and measuring the OD570 of bound dye, as in the previous paragraph described.
Statistical analysis
The experiments to study and compare survival patterns of Salmonella serotypes in soil and water and to evaluate biofilm formation on abiotic surfaces (glass coverslips and quartz particles) were randomized blocked designs with factorial treatment arrangements. All experiments were replicated three times. Salmonella populations were measured as log CFU/mL or g, and biofilm formation was measured by determining OD570 of bound dye. Differences in Salmonella population and biofilm formation over time and by serotype were compared using PROC GLM (General Linear Model) SAS v9.1 (SAS Institute, Inc., Cary, NC). The significant difference between the means was established by least significant difference tests. Variables included for analysis were Salmonella serotype, matrix, and day. Comparison of least square means of OD values was used to analyze cell attachment on glass and quartz over time. Significant differences were defined at p < 0.05, and highly significant differences were defined as p < 0.0001.
Results
Survival of Salmonella serotypes in soil
No significant difference was observed in cell population between serotypes during survival in soil over a 40 -day period at 25°C (p > 0.05). All Salmonella serotypes survived in soil for 40 days. The initial mean population for Salmonella serotypes on TSAYE was 9.27 ± 0.06 log CFU/g soil and 9.25 ± 0.16 log CFU/g soil on XLT4 (Table 1). A decrease in Salmonella population to 6.60 ± 0.39 log CFU/g of soil was observed through enumeration on TSAYE after 20 days of survival. Salmonella populations stabilized after decreasing during the first 20 days and increased by 1.1 log CFU/g by day 30 when enumerated on TSAYE. Significant differences in cells enumerated on TSAYE and XLT4 occurred on days 30 and 40, indicating sub-lethal injury (p < 0.05). At the end of the 40-day period, the mean population of Salmonella cells averaged 7.43 ± 0.54 log CFU/g on TSAYE and 5.15 ± 0.96 log CFU/g soil on XLT4.
Soil obtained from a tomato farm on the Eastern Shore of Virginia, passed through a #20 sieve, and autoclaved at 121°C for 12 min before use. Distilled deionized water was autoclaved at 121°C for 12 min before use.
Average Salmonella population in soil designated with the same lower case letters (a, b, c, d) are not significantly different. Average Salmonella population in water designated with the same upper case letters (A, B, C, D) are not significantly different.
Survival of Salmonella serotypes in water
All Salmonella serotypes survived in water for the sampling duration of 40 days. Higher populations of Salmonella were recovered from water after 40 days than from soil (p < 0.0001). No significant differences were observed between populations of the six Salmonella serotypes in water (p > 0.05). Initial mean populations of Salmonella cells in water on day 0 were 9.27 ± 0.08 log CFU/mL (TSAYE) and 9.27 ± 0.07 log CFU/mL (XLT4; Table 1). Salmonella populations enumerated on TSAYE and XLT4 were significantly different on days 10 and 30 (p < 0.05), indicating sub-lethal injury. The mean population of the Salmonella serotypes after 40 days was 7.70 ± 0.7 log CFU/mL water and 7.31 ± 0.81 log CFU/mL water on TSAYE and XLT4, respectively (Table 1).
Morphology on Congo Red agar
All six Salmonella serotypes—Anatum, Baildon, Braenderup, Javiana, Montevideo, and Newport—produced rdar morphotype after 48 h of incubation on Congo Red agar at 28°C, indicating the presence of curli fimbriae and cellulose production as extracellular matrix components.
Biofilm formation on glass coverslips
All the serovars tested, except Salmonella Anatum, demonstrated an initial increase in biofilm production on glass coverslip over time, followed by a decrease (Fig. 1). Salmonella Montevideo (OD570 0.74 ± 0.19) and Salmonella Javiana (OD570 0.41 ± 0.19) produced maximum biofilms on glass coverslips on day 6 (Fig. 1). Salmonella Baildon (OD570 0.50 ± 0.07), Salmonella Braenderup (OD570 0.78 ± 0.15), and Salmonella Newport (OD570 0.80 ± 0.23) demonstrated maximum biofilm production on coverslips on day 10, as indicated by CV dye retention. The biofilm formed by Salmonella Anatum linearly increased, resulting in maximum biofilm formation on day 14 (OD570 0.44 ± 0.26). On the glass coverslip surface, days 6, 10, and 14 were significant (p < 0.0001) in relation to changes in biofilm density. Glass coverslips had a mean residual OD570 of 0.09, resulting from bound CV dye, which was subtracted from the OD570 values obtained.
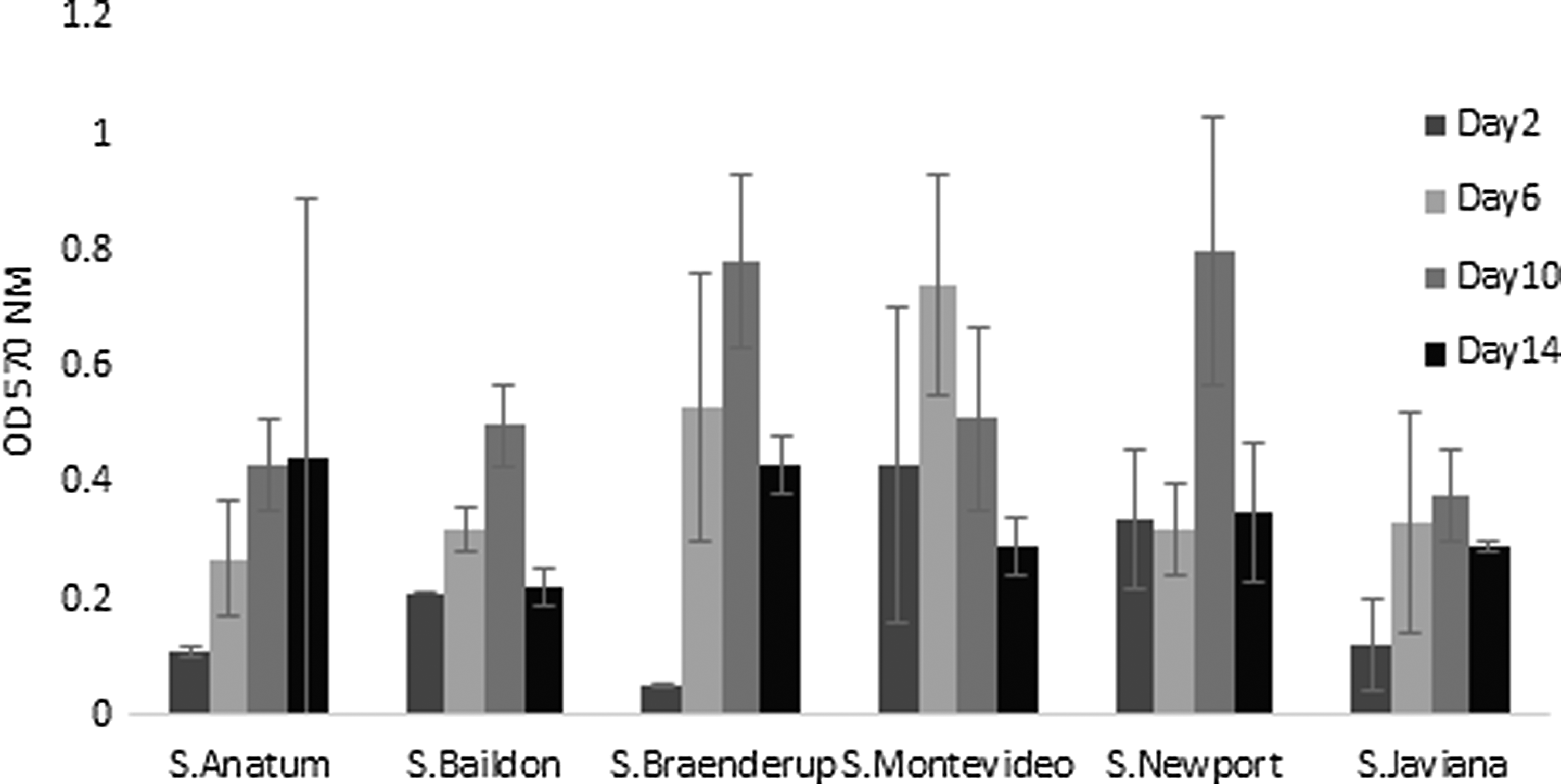
Quantification of biofilm density produced by Salmonella serovars on glass coverslips immersed in TSB using a modified CV staining and destaining procedure. OD of released dye was measured at 570 nm after destaining bound CV from coverslips using 33% acetic acid. OD, optical density; CV, Crystal violet; TSB, tryptic soy broth.
Biofilm formation on quartz particles
Different densities of biofilm were formed over time on quartz particles in comparison to glass coverslips by all serotypes tested, indicating that characteristics of surface and material played a significant role in biofilm development (p < 0.0001). Maximum biofilm production by Salmonella Anatum (OD570 0.78 ± 0.16), Salmonella Braenderup (OD570 0.62 ± 0.07), Salmonella Montevideo (OD570 0.71 ± 0.01), Salmonella Newport (OD570 0.67 ± 0.07), and Salmonella Javiana (OD570 0.74 ± 0.05) on quartz particles occurred on day 6 (p < 0.0001; Fig. 2). The density of biofilm material on quartz particles then decreased to day 14. Salmonella Baildon (OD570 0.66 ± 0.03) produced maximum biofilm density on day 2, following which the density of biofilm material on quartz particles declined until day 10 (OD570 0.24 ± 0.01) and increased on day 14 (OD570 0.33 ± 0.02). Quartz particles (control) had a mean residual OD570 from bound dye of 0.46, which was subtracted from the OD570 values obtained.
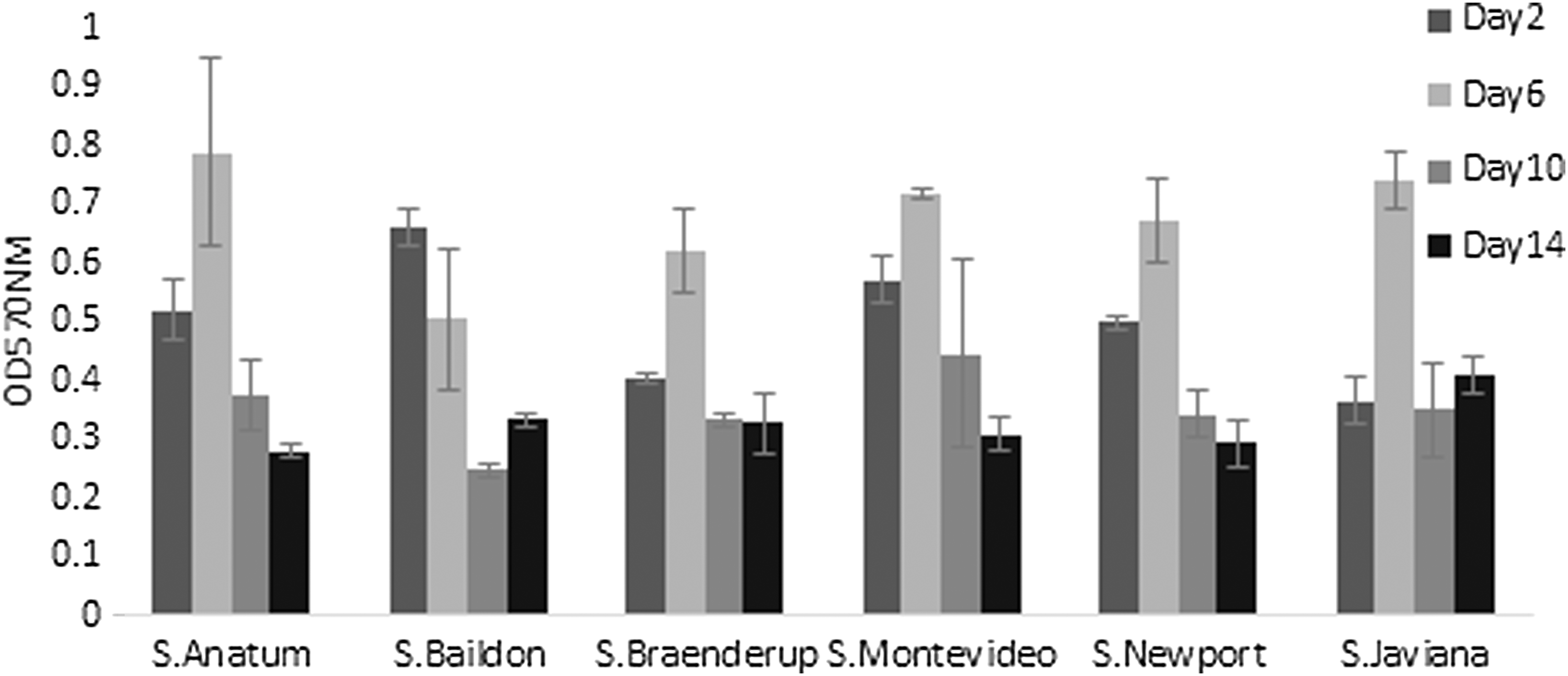
Quantification of biofilm density produced by Salmonella serovars on quartz particles immersed in TSB using a modified CV staining and destaining procedure. OD of released dye was measured at 570 nm after de-staining bound CV from quartz particles using 33% acetic acid.
Discussion
The results from this study indicate that no significant difference in survival in soil and water was observed between Salmonella serotypes tested. All the Salmonella serotypes tested were capable of forming biofilms on quartz particles, indicating a possible mechanism for long-term persistence in soil and sediment.
The conditions tested in this study did not account for stressors that foodborne pathogens might encounter in the field environment such as sunlight, temperature fluctuations, and competing organisms. Stressors such as sunlight (Zhang et al., 2015) and desiccation (Marsh et al., 1998) can cause reduced detection of foodborne pathogens on culture-based media due to transition into nonculturable states. A dual plating technique (Kumar et al., 2016) was used to enumerate Salmonella populations over 40 days to account for cell injury over time. While the population of Salmonella changed with time (p < 0.0001) in both soil and water, cell injury was detected on days 30 and 40 in soil, indicating that Salmonella cells were stressed as time progressed. In water, Salmonella cells demonstrated injury followed by recovery on both days 10 and 30, indicating that the population fluctuated over the 40-day period, with the highest decrease in population on day 10 (p < 0.05).
Salmonella have been known to survive in marine environments and persist in the presence of adverse conditions such as salinity, starvation stress, and temperature variations (Winfield and Groisman, 2003). Long-term persistence of the pathogen has been previously observed in irrigation ponds (Greene et al., 2008; Luo et al., 2015), indicating the pathogen's ability to adapt to stressors in hydrological niches and highlighting the importance of assessing irrigation water quality regularly. In the autoclaved soil samples used in this experiment, Salmonella survived over a period of 40 days, with a decrease in population over time. Similar durations of survival (21–30 days) of Salmonella have been reported in soil amended with compost, manure, and fecal slurries (Arthurson et al., 2010; Hruby et al., 2018).
Biofilms formed by bacteria allow persistence in the environment (Kumar and Micallef, 2017). All six tomato-associated Salmonella serotypes produced the rdar morphotype commonly associated with environmental persistence (White et al., 2006). A CV dye binding assay was used to account for EPS formation by Salmonella serotypes on the abiotic surfaces (Prouty et al., 2002; Peeters et al., 2008). All six serotypes also produced biofilm on quartz particles and glass coverslips, indicating an ability to attach to abiotic surfaces, though there were differences between serotypes for biofilm production over time.
Conclusion
Accidental or unintentional introduction of Salmonella into agricultural matrixes could occur through wildlife and domesticated animal or human feces, untreated biological amendments, and contaminated irrigation water. This study indicates that Salmonella serotypes Newport, Javiana, Montevideo, Baildon, Braenderup, and Anatum can survive in soil and water for extended periods and attach to surfaces and particulates through biofilms. The ability of Salmonella serotypes to persist in soil or water and attach to abiotic surfaces through biofilm formation affirms that contact surfaces, soil, water, and sediment should be considered as possible sources of cross-contamination in the farm environment.
Footnotes
Acknowledgments
The authors would like to thank Dr. Hengjian Wang for assistance with statistical analysis. Funding for this work was provided in part by the Virginia Agricultural Experiment Station and the Hatch Program of the National Institute of Food and Agriculture, U.S. Department of Agriculture.
Disclosure Statement
No competing financial interests exist.