
Abstract
Background:
Diseases caused by livestock-associated methicillin-resistant Staphylococcus aureus (MRSA) are an important global public health concern, and MRSA is increasingly being isolated in bovine milk. However, information on the genotype and antimicrobial resistance of MRSA in bovine milk in Xinjiang is limited. The objective of this study was to determine the antimicrobial-susceptible phenotypes and genotypes of the circulating MRSA clone isolated in bovine mastits milk samples in Xinjiang, China.
Methods:
Fifty six MRSA isolates collected from milk of bovine mastitis were investigated by multilocus sequence typing (MLST), staphylococcal protein A (spa) typing, and a minimum inhibitory concentration test with 21 antimicrobial agents.
Results:
Antibiotic resistance results showed that 47.4% of the isolates were resistant to 16 or more antibiotics. Twelve MLST types were defined in this study, and ST398 (n = 7) and ST2393 (n = 2) were found to be the most prevalent types. Seven spa types (t034, t269, t4030, t114, t35, t189, and t7589) were identified, of which t034 (n = 7), t189 (n = 3), and t4030 (n = 3) were predominant. Here, 3 MRSA ST188 is reported among human MRSA isolates in China, and this is the first time that it is reported in bovine MRSA strains.
Conclusions:
The antimicrobial susceptibility of MRSA in this area exhibited multidrug resistance, and clonal complexes CC398 and CC188, which have been reported among human MRSA isolates, do occur in Xinjiang dairy cows. This study provides a foundation for further MRSA monitoring.
Introduction
D
The first MRSA isolates from bovine mastitis were reported in 1972 (Devriese et al., 1972), and the investigation of MRSA and its characteristics has increased globally. After humans, cattle are the second-largest reservoir of MRSA (Sakwinska et al., 2011). A recent study of an MRSA variant found in both humans and cows suggested that the host range of MRSA has expanded (García-Álvarez et al., 2011). Studies on MRSA ST398 and ST5 from pigs and chicken have also demonstrated such a host shift (Loo et al., 2007; Lowder et al., 2009). The inability to control dairy cow mastitis caused by S. aureus is largely attributed to the fact that little is known about the genotype and the antimicrobial resistance phenotype of S. aureus, especially MRSA isolates from bovine milk (Gindonis et al., 2013).
Gene transfer between human and animal pathogens has been demonstrated by numerous genetic analyses, including those of data from human and bovine MRSA isolates (Juhasz-Kaszanyitzky et al., 2007). Recent studies also identified several S. aureus sequence types (STs) that are associated with zoonotic transmission or transmission from a recent common ancestor. These bacteria include the poultry-associated ST5 (ST5) and livestock-associated MRSA (Smith et al., 2005a) clone ST398 (Lowder et al., 2009; Price et al., 2012). MRSA ST398 was developed in humans as a methicillin-sensitive S. aureus (MSSA) strain and was then transferred from humans to livestock (Weese et al., 2010), first to pigs and then to cattle, wherein it acquired the mecA gene and become MRSA (Price et al., 2012).
Numerous methods have been developed to analyze the genetic characteristics and phylogeny of MRSA. Multilocus sequence typing (MLST) has increasingly become the gold standard for the evolutionary study of microbes, and it has a high discriminatory capability (Enright et al., 2002; Stefani et al., 2012). This typing method differentiates bacteria isolates by comparing nucleotide variations in housekeeping genes and enables an in-depth investigation of their epidemiology. Another widely used method is staphylococcal protein A (spa) typing, which analyzes bacteria sequence repeats at the spa × region and it also has excellent discriminatory power. Spa typing not only provides unambiguous typing results, but is also much easier to perform and less expensive than MLST or pulsed-field gel electrophoresis (Strommenger et al., 2006; O'Hara et al., 2016).
Data from different countries have revealed a variety of MRSA genotypes (Smith et al., 2005b; van Wamel et al., 2006; Thurlow et al., 2013), suggesting that these MRSA strains have typical molecular characteristics. MRSA from swine, swine farm workers, and pet animals were also reported from China (Enright et al., 2000; Wyllie et al., 2011). MRSA ST97, ST126, ST133, and ST398 were isolated from bovine milk samples in many countries. (Aires-de-Sousa et al., 2007; Smyth et al., 2009; Hata et al., 2010; Vanderhaeghen et al., 2010). However, the genetic characterization of MRSA and the epidemiology and antimicrobial resistance profile of MRSA from dairy cows in North Xinjiang have not been studied.
This study aims to evaluate the occurrence of MRSA from Xinjiang, China, subclinical mastitis milk samples, and to characterize the antimicrobial resistance and genotype of MRSA isolates using MLST/Based Upon Related Sequence Type (BURST) and spa typing/Based Upon Repeated Pattern (BURP).
Materials and Methods
Ethics
Milk samples were obtained from dairy cows with naturally occurring clinical mastitis under the ethical approval granted by the College of Animal Science and Technology, Xinjiang Agricultural University. The milk samples were collected by professional veterinarians with the permission of the dairy farm owner. Conventional milking and sample collection methods were used, and no pain-causing procedures were involved. This study did not involve endangered or protected species.
Sampling and isolation of MRSA
Milk samples were taken from cattle with subclinical mastitis (somatic cell count ≥200,000 cells/mL) (Hiitiö et al., 2017). Milk samples were collected in 2014–2015 from five different farms 100 cows per farm in four different geographically diverse regions of North Xinjiang: Urumqi, Changji, Shihezi, and Yili. For milk collection, the udders of all the sampled cattles were cleaned with water and the teat surfaces were disinfected with 70% ethanol. The first three streams of milk were discarded. The milk samples were collected in 15 mL sterile plastic tubes. The collected samples were kept on ice, transported to the laboratory, and processed for bacterial isolation.
Samples were inoculated on agar plates (trypticase soy agar supplemented with 10% NaCl and phenol red mannitol) and incubated at 37°C for 15 h. Yellow colonies were assumed to be S. aureus (Li et al., 2015). Further identification of these staphylococcal colonies was confirmed using conventional methods, including Gram staining, colony morphology, catalase testing, and coagulase testing. In total, 56 MRSA were isolated from 572 milk samples from five different farms. In all, 572 individual quarter milk samples were obtained from 500 cows. S. aureus ATCC29213 and ATCC43300 served as the quality control strains.
Genetic confirmation of MRSA
DNA from S. aureus was prepared using a Column Bacterial DNA kit (Tiandz, Inc., Beijing, China). In brief, 1 mL of the culture was incubated overnight and then pelleted by centrifugation at 10,000 g for 2 min. The cell pellet was resuspended in 600 μL of lysostaphin (50 μg/mL) (Tiandz, Inc.) and the mixture was then incubated at 37°C for 1 h. Total DNA was extracted according to the manufacturer's protocol. In addition, all S. aureus isolates were confirmed using a polymerase chain reaction (PCR) assay targeting an S. aureus-specific region of the thermonuclease gene (nuc), as described previously (Lovseth et al., 2004; Fournier et al., 2008). In addition, isolates were confirmed as MRSA by using penicillin-binding protein 2a gene (mecA) detection.
Antibiotic susceptibility testing
Antibiotic susceptibility testing was performed by the micro broth dilution method according to Clinical and Laboratory Standards Institute guidelines (CLSI, 2015). All S. aureus isolates were subjected to antimicrobial susceptibility testing for 21 antimicrobial agents: amoxicillin, ampicillin, cefuroxime, ceftiofur, cefoxitin, penicillin, gentamicin, streptomycin, levofloxacin, enrofloxacin, ciprofloxacin, clarithromycin, norfloxacin, doxycycline, sulfafurazole, sulfadiazine sodium, rifampin, erythromycin, tetracycline, cefuroxime, and clindymycin, broth microdilution testing was performed in accordance with the guidelines in CLSI (2010) document. Minimum inhibitory concentration (MIC) values were determined visually, after 24 and 48 h incubation, as the lowest concentration of drug that caused complete inhibition. The S. aureus reference strains ATCC29213 and ATCC43300 served as quality control strains in each MIC test.
MLST and eBURST
All 56 S. aureus isolates (isolated from Shihezi, Yili, Changji, and Urumqi 4 different area in xinjiang) were randomly subjected to MLST. MLST was performed as described previously by sequencing the fragments of seven housekeeping genes (arcC, aroE, glpF, gmk, pta, tpi, and yqiL), and the sequences of PCR products were compared with those in an MLST database (
Spa typing
The polymorphic X region of the protein A gene (spa) was amplified and sequenced for spa typing. The spa types were assigned using an online spa database (
Phylogenetic analysis
The reconstruction of evolutionary relationships was performed using MGEA 4. MLST data were used to construct trees using Splits Tree software. For comparison, a Splits Tree analysis of the concatenated MLST sequences of all STs was constructed.
Results
Bacterial identification by nuc PCR and detection of mecA by PCR
The 56 S. aureus isolates were confirmed by the presence of the nuc gene by PCR. We determined the presence of mecA in the S. aureus isolates and identified MRSA in the S. aureus isolates.
Antimicrobial susceptibility testing
The antibiotic susceptibility of the analyzed isolates is shown in Figure 1. The overall resistance rates were high with cefuroxime (100%), ceftiofur (89.5%), cefoxitin (84.2%), sulfadiazine sodium (84.2%), and clindamycin (84.2%), while the overall resistance rates were low with doxycycline (31.6%) and gentamicin (36.8%). These isolates were generally resistant to penicillins and lincosamides.
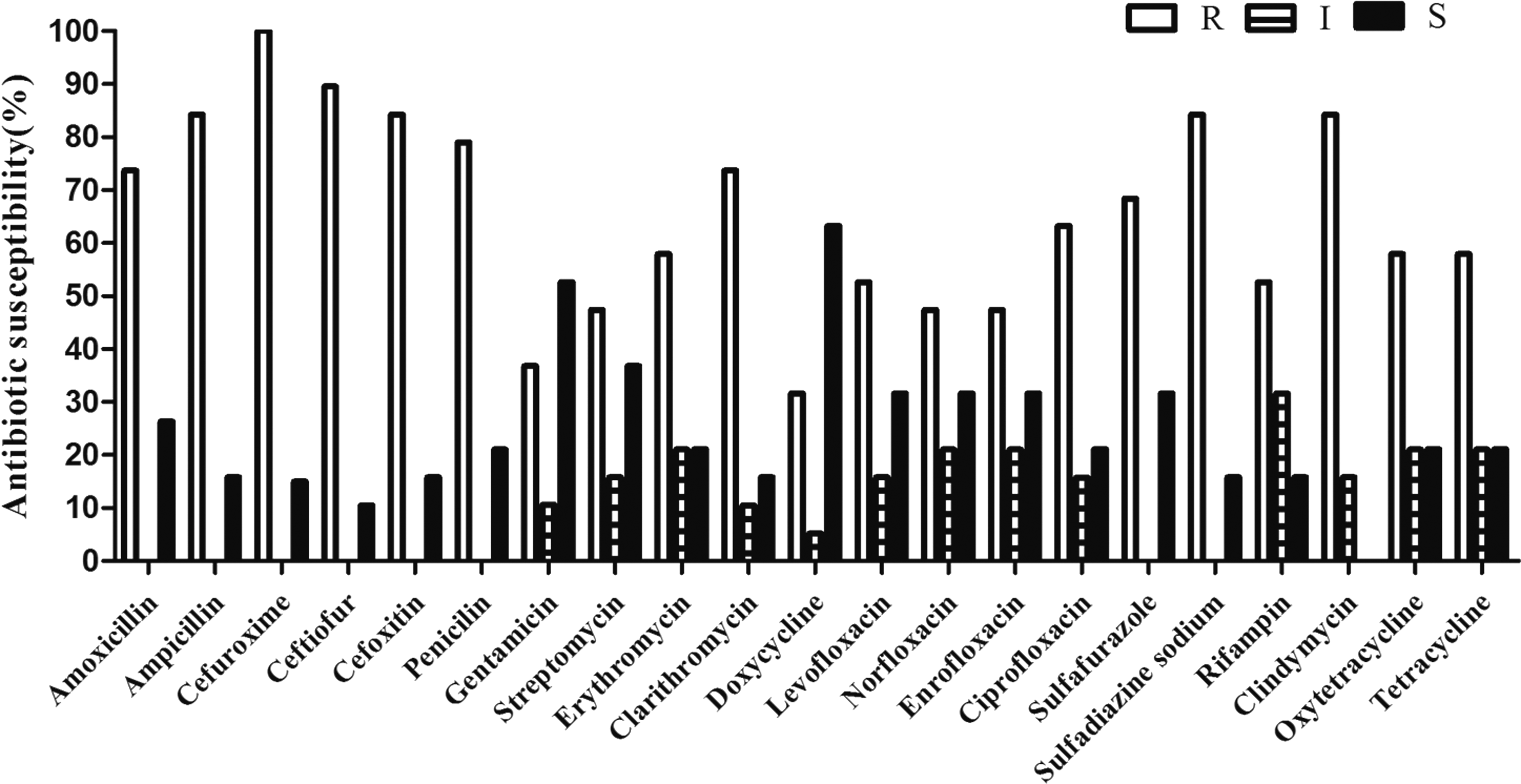
Antimicrobial susceptibility profiles of MRSA isolates from Xinjiang. MRSA, methicillin-resistant Staphylococcus aureus.
In our study, all 56 MRSA strains were resistant to cefuroxime, and 48 strains (85.7%) were resistant to cefoxitin, ampicillin, sulfadiazine sodium, and clindamycin. Seventeen strains (30.4%) were resistant to ceftiofur and 45 strains (80.4%) were resistant to penicillin. Amoxicillin and clarithromycin resistance was observed in 42 strains (75%). Twelve strains (21.4%) were resistant to ciprofloxacin. Resistance to more than 10 different antimicrobials was observed in these bovine MRSA isolates, and most of these MRSA isolates were multiresistant. According to our multi-resistance analysis of these 56 MRSA isolates, about 47.4% of MRSA isolates were resistant to more than 10 kinds of antibiotics, and 47.4% of MRSA isolates were resistant to more than 16 different antibiotics.
MLST and spa typing of MRSA isolates
MLST typing (Table 1) revealed that 56 representative isolates could be grouped into 12 STs, with ST398 as the most dominant (41.0%). The other types included ST2393 (10.7%), ST1079 (3.6%), ST2199 (5.4%), ST1490 (3.6%), ST188 (5.4%), ST2154 (3.6%), ST97 (7.1%), ST124 (5.4%), ST617 (5.4%), ST454 (3.6%), and ST2140 (5.4%).
A, Urumqi; B, Changji; C, Shihezi; CC, clonal complexes; D, Yili; MLST, multilocus sequence typing; spa, staphylococcal protein A.
The 56 isolates were grouped into seven spa types: t034, t267, t4030, t114, t359, t189, and t7589. Most of the S. aureus isolates studied here were spa type t034 (n = 21), followed by t189 (n = 9) and t4030 (n = 9). Spa types t267 (n = 6), t359 (n = 6), t114 (n = 3), and t7589 (n = 2) were also detected (Table 1).
BURST analysis uses gene sequences to identify genetically related groups of bacteria. Our MLST typing results were further analyzed using eBURST software. The evolutionary relationships between STs are shown in Figure 2. ST2393 and ST1079 were evolutionarily distant from the other 10 STs.
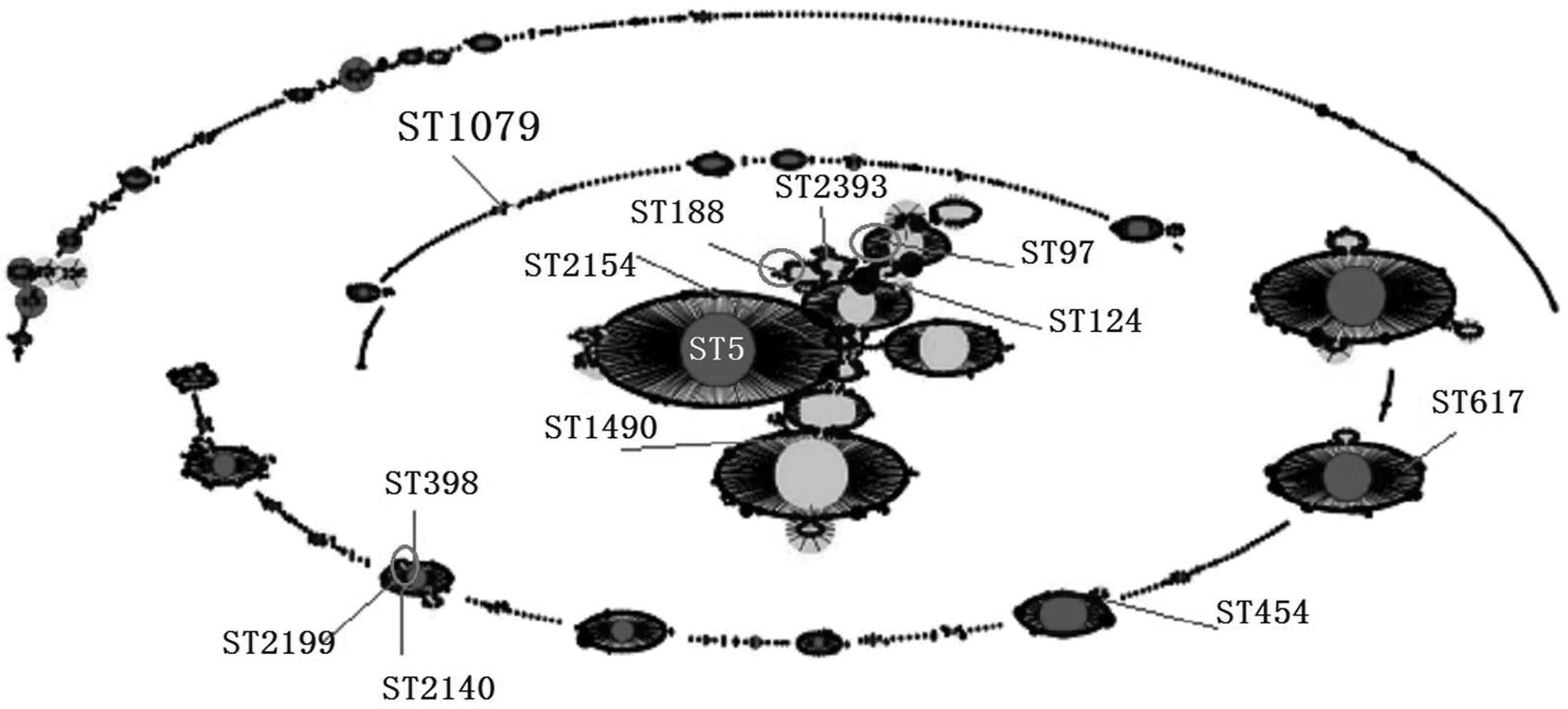
Population snapshot analyses by eBURST of MRSA isolates in this study based on the S. aureus MLST database (
The neighbor-joining method was used to analyze the evolution of different STs. The 56 MRSA isolates were divided into 7 CCs: CC398 (n = 29), CC45 (n = 5), CC8 (n = 2), CC97 (n = 7), CC188 (n = 9), and CC1 (n = 2) (Table 1). CC398 isolates were the most common. The evolutionary distance between CC groups are shown in Figure 3.
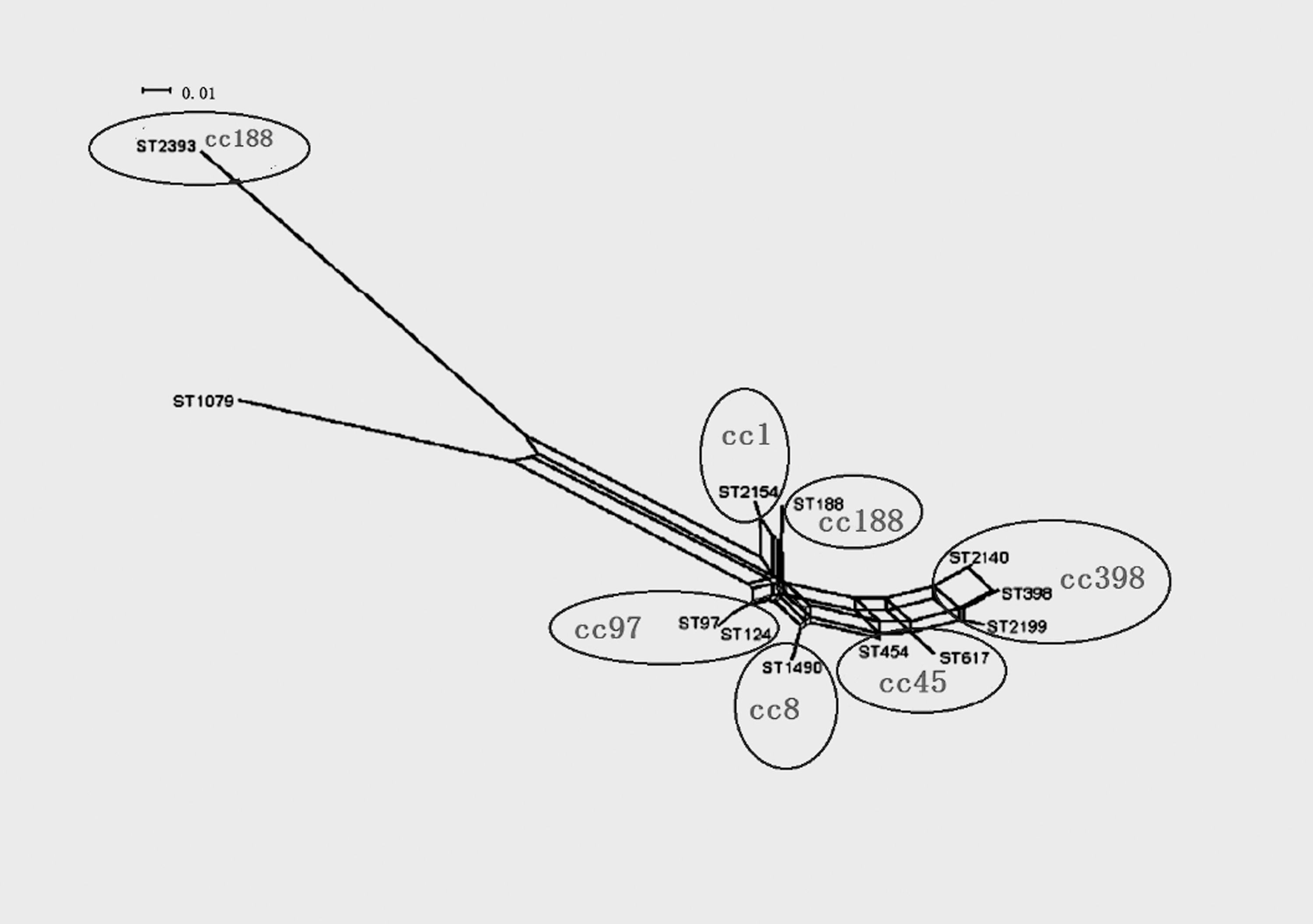
Network graph visualizing relationships between concatenated MLST sequences of all STs mentioned in this study, generated by the Neighbor-net algorithm of Splits Tree 4. Each CC is composed of STs that cluster with ≥70% bootstrap confidence. ST188 and ST2393 belonged to CC188, ST398, ST2140, and ST2199 belonged to CC398. ST97 and ST124 belonged to CC97. ST1490 belonged to CC8. ST617 belonged to CC45. ST2154 belonged to CC1. MLST, multilocus sequence typing.
Discussion
Recently, due to the high use of antibiotics in food animals, such as dairy cows with mastitis, an MRSA strain with zoonotic potential emerged and became a risk to human health. In this study, we observed relatively high multidrug resistance rates among the MRSA isolates analyzed. Moreover, limited information is available on genetic characterization of MRSA of bovine mastitis milk in xinjiang, china. In this study, 56 mecA-positive MRSA from 572 milk samples were isolated and analyzed. Compared with some clinical studies on S. aureus from bovine mastitis in the United States and European countries (Erskine et al., 2002; Oliver and Murinda, 2012) the multi-resistance level in our study was relatively high and 47.3% isolates were resistant to 16 or more antimicrobials. The prevalence of ampicillin (≥0.5 μg/mL), ceftiofur (≥8 μg/mL), cefoxitin (≥8 μg/mL), sulfadiazine sodium (≥512 μg/mL), and clindamycin (≥4 μg/mL) resistance is extremely high. This result suggests uncontrolled or common use of these antimicrobials for treatment of bovine clinical infections in dairy cow farms. This result therefore emphasizes the importance of antimicrobial susceptibility testing and regular surveillance of bovine MRSA in dairy cow farms. Our antimicrobial resistance data were much higher than those obtained from the United States, European countries, and other regions of China (Erskine et al., 2002; Oliver and Murinda, 2012; Li et al., 2015).
Regular surveillance of the MRSA in livestock and humans is important (McAdam et al., 2012). Differences in epidemiological characteristics and dependable molecular methods may be useful for understanding MRSA dissemination and for designing effective strategies to prevent MRSA infection. According to a study by Wang et al. (2011), ST97, ST965, ST9, ST6, and ST50 have been identified from bovine in five provinces of China, including some areas in Xinjiang. Studies performed on isolates from Shanxi Province have identified ST71, ST2683, ST97, and ST2738 (Li et al., 2015). The present study identified 12 different STs. With the exception of ST97, none of these STs were similar to those reported by the other animal studies in China.
This study is the first report on the ST398 MRSA isolate in dairy cow milk in China. This finding will help us to understand the genetic characteristics of MRSA in China. We also found that the MRSA ST398 isolate occurs at a relatively high frequency. This genotype was first identified in a clone from a pig and was later recovered from other food animals and from humans (Witte et al., 2007). MRSA ST398 was also found in individuals who had close contact with animals (Zhao et al., 2012; Yan et al., 2014). A recent study of hospital samples in China indicated that ST398 was the most prevalent clone in healthy adults (Yan et al., 2015).
MRSA ST398 is thought to be transmitted from pigs to humans and cows. Cows are regarded as the most likely transmission path, with humans being the vector of transmission. Many European countries have identified ST398 in bovine mastitis (Fessler et al., 2010; Huber et al., 2010). ST130 MRSA was recently shown to have spread from cows to humans, resulting in clinical infections (Harrison et al., 2013).
Previous comparison of ST398 spa types revealed that the most common were spa types t011 and t034. Researchers in Germany and Switzerland identified an ST398/t034 strain (Monecke et al., 2007). Eight MRSA ST398/t034 strains were also identified in our study (Table 1). We also identified MRSA ST398 isolates with other spa types: t4030-ST398 and t189-ST398.
A genetic variation study of MRSA from bovine milk in Japan identified MRSA ST188 and t189 (Hata et al., 2010). This type was also reported to occur at a relatively high frequency in hospitals in China (Chen et al., 2012; Yu et al., 2012). ST188 MRSA isolates were also detected in bovine milk samples in our study. The presence of human-related MRSA isolates in mastitic milk samples may indicate that these isolates are transmitted between humans and animals.
We compared our data with those of a recently reported study about community residents and healthcare workers at Sun Yat-Sen University, Guangzhou, China. ST398 and ST188 were also detected in the human population, and ST188 was the most prevalent type among the tested samples. Previous studies determined that the five most prevalent human clinical MRSA types in China are CC5, CC8, CC188, ST398, and CC59 (Chen et al., 2015). These findings indicate that ST188 and ST398 MRSA strains should be investigated further.
The ST97 and ST188 genotypes are typical bovine-associated genotypes (Hasman et al., 2010; Hata et al., 2010) and the most common STs in bovine mastitis (Zadoks et al., 2011). These genotypes have been isolated from bovine milk in Brazil, Chile, Norway, Canada, Italy, Netherland, Spain, the United Kingdom, and the United States (Hata et al., 2010). The present findings show that ST97 has become the dominant lineage in Japan, the Netherlands (Ikawaty et al., 2009), Brazil (Rabello et al., 2007), Chile (Smith et al., 2005a), and the United States (Smith et al., 2005b). In this study, both ST97 and ST188 lineages were identified in our bovine samples. ST97 also has been reported from mastitic milk samples from Shanxi province in China (Li et al., 2015). CC97 was also reported in North and South America, Asia, and Africa (Diep et al., 2003; Ruimy et al., 2009; Schuenck et al., 2009; Udo et al., 2011).
Livestock might serve as an important reservoir for the spreading of new human pathogenic MRSA. According to a recent report from Denmark, MRSA complex CC97 was isolated from human infections, providing evidence of a livestock-to-human jump. MLST typing has greatly improved the ease of comparison across herds, countries, and host species. This method was used to show that ST1, ST8, ST9, and ST398 are associated with bovine mastitis and human infections (van den Borne et al., 2010; Vanderhaeghen et al., 2010).
In this study the occurrence of identical and predominant MRSA genotypes (spa and ST) differ in isolates from four divergent geographical regions and displayed some geographic characteristics. MRSA isolates from Urumqi and Shihezi displayed more molecular genotypes. In addition, bovine-specific genotype ST97 and human-related genotype ST188 distributed in these two areas and ST97 was exclusively found in farms located in Urumqi, ST188 was exclusively found in farms located in Shihezi. The genotypes diversity in these two regions might be the result of large breed scale and management practices of these dairy cow farms.
Conclusions
The inappropriate use of antibiotics in the treatment of mastitis may have created a selective pressure on S. aureus that lead to the emergence of antibiotic-resistant bacteria, especially for foodborne pathogens MRSA. Our antibiotic resistance data showed that MRSA isolates investigated in this study displayed multidrug resistance, and it is, therefore, essential to implement more effective and comprehensive measures to monitor and disseminate antibiotic resistance in S. aureus and MRSA isolates from dairy cow milk.
A genotype analysis identified 12 MLST types and 7 spa types, among them ST398, ST2393, t034, and t189 were predominant. MRSA isolate ST188, which has been previously reported in humans in China, has, for the first time, been reported in cows.
Footnotes
Disclosure Statement
All authors disclose potential sources of conflict of interest relevant to the present article.