
Abstract
The monophasic variant of Salmonella Typhimurium is highly prevalent in human and in pork. However, little is known about colonization dynamics and serology in pigs. We orally inoculated 24 seven-week-old piglets with 109 CFU/pig of a porcine strain of monophasic Salmonella Typhimurium in an experimental trial. Three groups of eight piglets were orally inoculated and monitored for 21, 49, or 84 days post-inoculation until necropsied. From 3 days post-inoculation to necropsy, individual feces were sampled twice weekly and blood once weekly. At necropsy, the tonsils, mesenteric lymph nodes, and the contents of the duodenum, jejunum, ileum, and cecum were collected from each pig. We determined the number of CFU/g in all the samples and measured also Salmonella antibodies in OD% in all blood samples. At different times during the trial, we tested by MLVA (Multilocus Variable Number Tandem Repeat Analysis) the genomic stability of the strain after passing through the intestinal tract. Salmonella was continuously excreted by pigs, ranging from 1.4 to 5.8 log10 CFU/g. At necropsy, Salmonella was present in all samples, but the tonsils were particularly infected. Salmonella antibodies were detected in five pigs 7 days post-inoculation. At 49 days post-inoculation, all the pigs were seropositive. We observed new MLVA types for 3.3% of the isolates tested over the trial. Our study allowed us to show the serovar's ability to persist in pigs after infection up to 84 days post-inoculation. We demonstrated that Salmonella seroconversion appeared earlier than in naturally infected pigs and that the strain's genome can evolve after passing through the digestive tract of pigs.
Introduction
S
Longitudinal studies on naturally infected pig herds have been conducted to determine the dynamics of Salmonella spp. infection over time (Lynch et al., 2017). However, due to the multiple factors that may be involved in Salmonella pig infection, results vary greatly between herds and studies (Lynch et al., 2017). Experimental trials to assess the infection in pigs have been reported for Salmonella Typhimurium, but animals were monitored for 28 or 32 days p.i. (post-inoculation) (Boyen et al., 2009; Ivanek et al., 2012). No study described the colonization and serology of pigs by the monophasic Salmonella Typhimurium serovar over an extended period of time.
Under experimental conditions, bacteria can genetically adapt to environmental changes, and virulence may evolve. Genetic variations have previously been described for Campylobacter after passage in pigs (Leblanc-Maridor et al., 2011) and for Salmonella in mice (Koskiniemi et al., 2013).
The present work was designed to study the fecal excretion and colonization of pigs after inoculation with a monophasic Salmonella Typhimurium strain in an experimental trial. We also looked at the seroconversion to Salmonella and followed the genetic stability of the strain after passing through the gastrointestinal tract.
Materials and Methods
Strain
We used a S.1,4[5],12:i:- strain that we isolated from pig feces on a French farm in 2014 (S14CH63). To enumerate the strain in samples during the trial, it was made resistant to Rifampicin after several passages on plate count agar (BioMerieux, France) supplemented with Rifampicin (100 mg/L; Sigma, France).
To check the similarity of the S14CH63-Rif strain with the S14CH63 strain, the two strains were serotyped, by classical serotyping and polymerase chain reaction (PCR), as recommended by EFSA (2010) with a Bugarel's adapted protocol (Bugarel et al., 2012), typed by pulsed-field gel electrophoresis (Ribot et al., 2006) and Multilocus Variable Number Tandem Repeat Analysis (MLVA; Profile A: 3-14-10-N-211) (Larsson et al., 2009) and antimicrobial susceptibility tested to 16 drugs.
Experimental design
The in vivo protocol was approved by the Ethics Committee in Animal Research N°16 of the French Ministry of National Education, Higher Education and Research (License APAFIS 2697-2015110409457994). The trial was conducted on 32 secondary specific pathogen free piglets born at the ANSES Ploufragan protected animal facilities. At 4 weeks of age, the piglets were weaned and isolated in biosecurity level 3 experiment rooms. The 24 piglets to be inoculated were distributed in 3 rooms (Groups G1, G2, and G3) in 2 pens (P1–P2, P3–P4, and P5–P6, respectively) with 4 piglets per pen. The three groups were monitored, respectively, for 21, 49, and 84 days p.i. until necropsy. Eight control pigs were placed in another room. Biosecurity measures were carried out by the animal facility staff. Pigs were monitored daily, rectal temperature being measured and clinical manifestations recorded. Food consumption and body weight were reported weekly.
At day 0, the 24 piglets were orally inoculated with 10 mL of TS (Tryptone Salt Broth; BioMerieux) containing 109 CFU (colony-forming unit) of S14CH63-Rif Salmonella strain using a cannula attached to a luer-lock syringe. Control pigs received 10 mL of TS.
Sampling and necropsy
One week before inoculation, we checked that pigs were negative to Salmonella in feces using the NF U47-102 method (AFNOR, 2008).
Samples were 1:10 diluted in buffered peptone water (BPW; BioMerieux). After 18 h at 37°C, selective enrichment was performed on Muller-Kauffmann tetrathionate-novobiocin broth and modified semisolid rappaport-vassiliadis agar (Biokar Diagnostics, France). After incubation, enrichments were streaked on xylose lysine deoxycholate (XLD; Biokar Diagnostics) and Rapid'Salmonella (Bio-Rad, France). Sample was negative if no typical colonies were observed.
Individual feces were sampled directly from the animal's rectum twice weekly from day 3 p.i. to necropsy, and individual blood was obtained once weekly from the jugular vein from day 3 p.i. for pigs in G1 and G2 and from day 7 p.i. for pigs in G3. Upon arrival to the laboratory, feces were immediately analyzed for Salmonella detection and enumeration and blood centrifuged at 3500 rpm for 5 min to recover the serum. The sera were stored at −20°C until analysis.
All pigs of G1, G2, and G3 were necropsied at 21, 49, and 84 days p.i., respectively. Two control pigs were slaughtered at 21 and 49 days p.i. and four at 84 days p.i. Pigs were euthanized after intravenous inoculation of tiletamine and zolazepam (Zoletil 100; Virbac, France). At each pig's necropsy, organs and tissues from the abdominal and thoracic cavities were observed, and mesenteric lymph nodes (MLN), tonsils, and intestinal contents from the duodenum, jejunum, ileum, and cecum were collected.
Enumeration and detection of Salmonella
Samples were first 1:10 diluted in BPW and then serial diluted in TS until dilution 10−4. One milliliter of the 10−1 dilution was seeded on three plates of XLD agar supplemented with rifampicin (XLD-Rif), and 100 μL of the following dilutions were plated on XLD-Rif.
The 1:10 diluted samples and the plates were incubated at 37°C for 18 h.
If typical black colonies of Salmonella were observed on XLD-Rif, these were enumerated, and we expressed the result in CFU/g of sample. If not (i.e., under the limit of 10 CFU/g), we continued the detection as described previously. Typical colonies on XLD-Rif were kept in Peptone Glycerol Broth at −70°C until MLVA typing. We checked also the absence of Salmonella in the samples collected from control pigs throughout the experiment.
Serological response to Salmonella infection
For serology, we used the IDEXX Swine Salmonella Ab Test® (IDEXX, France). Samples with OD% values equal to or greater than 15% were considered positive as recommended by the manufacturer.
Genetic stability after in vivo passage
To evaluate the genetic stability of the strain after in vivo passage, we selected 368 isolates from the 8 pigs monitored for 84 days p.i. On day 17 and on day 49, we considered, respectively, 108 and 120 isolates from feces. On day 84, 140 isolates were selected: 20 isolates for each organ (tonsils and MLN), 20 for each part of the intestine (duodenum, jejunum, ileum, and cecum), and 20 for feces.
The 368 isolates were typed by MLVA. The 5-loci MLVA allele profiles were determined according to the previously published harmonized method (Larsson et al., 2009). DNA was extracted with an InstaGene matrix according to the manufacturer's instructions (Bio-Rad). STTR9, STTR5, STTR6, STTR10, and STTR3 were amplified within a multiplex PCR assay using the QIAGEN Multiplex PCR Kit (Qiagen, France). Amplified fragments were separated by capillary electrophoresis with the 3100-Avant Genetic Analyzer (Applied Biosystems, France). The GENEFLO™ 625 Fluorescent DNA Ladder ROX (Chimerx) was used as a labeled size standard marker. Sequencing data were imported, and the size of the PCR products and number of repetitions in each locus were determined with BioNumerics v7.5 software (Applied Maths, Belgium).
Statistical analysis
Statistical analysis was performed using R software (R version 3.2.4).
The excretion level was compared between groups for each sampling time postinoculation with a Kruskal–Wallis test (p < 0.05). Post hoc pairwise comparisons were then performed using the Holm test to adjust the p-values of these comparisons given the number of tests conducted (p < 0.05). The correlation between the amounts of Salmonella in feces between successive sampling times was assessed with a Spearman correlation test (p < 0.05). The amount of Salmonella shed by each pig on the whole trial period was estimated by calculating the area under the curve of excretion (AUC). A Kruskal–Wallis test was used to compare the AUC between groups (p < 0.05). We compared the colonization levels of each organ at the three dates of necropsy using the Kruskal–Wallis test (p < 0.05).
Results
Clinical monitoring
No clinical signs were observed in control pigs. Inoculated pigs had elevated rectal temperature (greater than 40°C), lethargy, and diarrhea the day after inoculation. Then, no clinical signs were noted throughout the trial in G1 and G2. In G3, animals had a short episode of diarrhea at 42 days p.i., due to an unknown etiology. The pigs' growth and feed consumption were normal throughout the trial. At 21 days, no significant difference was observed in the average body weight gain of pigs in G1, G2, G3, and Control groups (889, 949, 721, and 787 g/day, respectively) (p = 0.112).
Salmonella excretion kinetics in feces
The control pigs remained negative for Salmonella in feces throughout the trial. All the inoculated pigs were continuously excreting Salmonella from day 3 p.i. until day 84 p.i., with variations in the amount of Salmonella (Fig. 1). The highest level of excretion in feces was on day 3 p.i. (5.8 ± 1.6 log10 CFU/g) and the lowest on day 52 p.i. (1.4 ± 0.7 log10 CFU/g) (Table 1). The amount of Salmonella in feces decreased significantly (p < 0.01) dropping by 2.4 log10 CFU/g on day 7. The excretion curves of each pig from day 0 to 21 were similar, with the peak on day 3 p.i. (Fig. 2). No significant difference was found between individual excretion of pigs in G1 and G2 (p > 0.05). In contrast, there was significant difference in individual excretion of pigs in G3 (p < 0.01). There was a strong positive (rho ≥0.70) significant correlation (p < 0.05) of the amount of Salmonella in feces between days 7–10, 35–38, 46–49, 49–52, 52–56, 56–60, and 66–70.
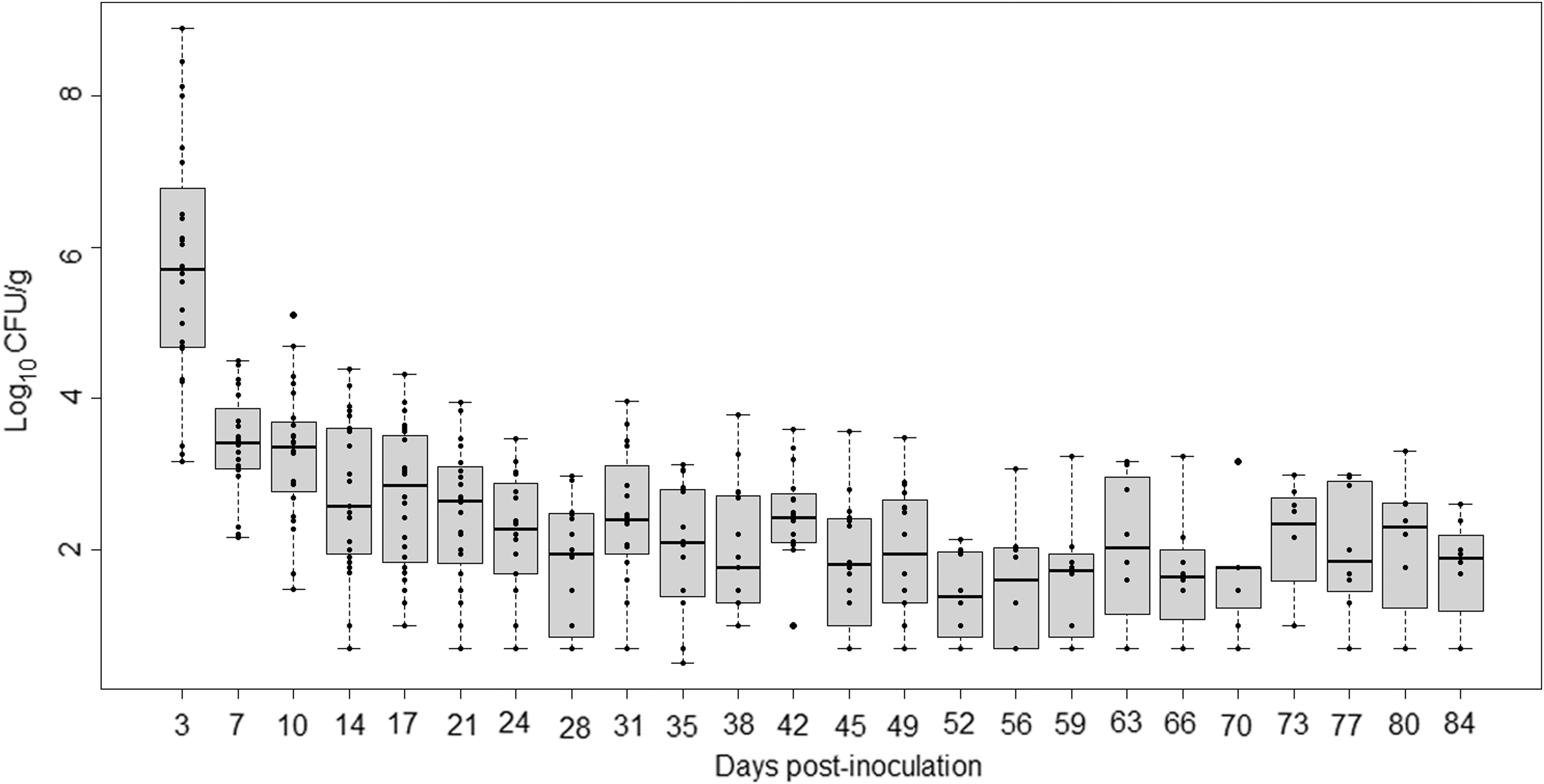
Monophasic Salmonella Typhimurium excretion kinetics for all pigs over 84 days (in log10 CFU/g). Each point represents the numeration data for each pig on each sampling day post-inoculation. Each box-plot shows the distribution of the data within a day, for each day post-inoculation. CFU, colony-forming unit.
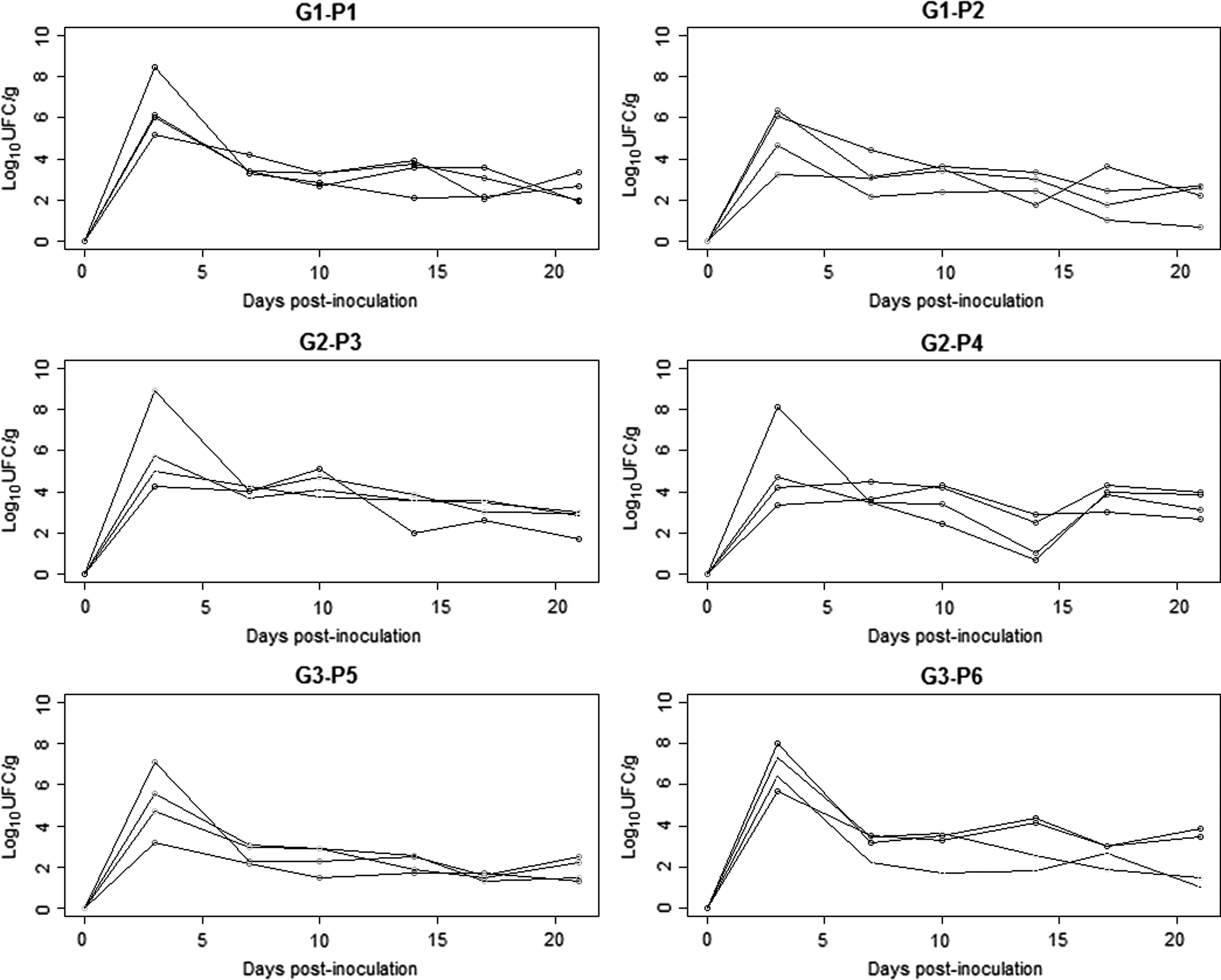
Individual curves for excretion of monophasic Salmonella Typhimurium in feces (in log10 CFU/g) per pen over 21 days (n = 24 pigs). G1, Group 1 (monitored for 21 days); G2, Group 2 (monitored for 49 days); G3, Group 3 (monitored for 84 days); P1 to P6, Pen 1 to Pen 6. CFU, colony-forming unit.
n = number of pigs monitored for each sampling day (D0 to D84).
Only 16 pigs in Groups 1 and 2 were sampled on D3 for serology.
—, no sample; CFU, colony-forming unit; SD, standard deviation.
Colonization levels in organs and intestinal contents
The average colonization levels of Salmonella per organ for the three groups range from 0.6 ± 0.9 log10 CFU/g (duodenum) to 5.0 ± 1.4 log10 CFU/g (tonsils) (Table 2). Except for tonsils, samples taken on day 21 p.i. (G1) were more contaminated than those obtained on days 49 and 84 p.i. The colonization levels in tonsils, mesenteric lymphatic nodes, jejunum, ileum, and cecum all significantly differed (p < 0.01) between the three necropsy days. All the ileum, cecum, and tonsils were contaminated, whatever the group. Tonsils were the most contaminated organs, particularly on day 49 p.i. (G2) (6.0 ± 0.3 log10 CFU/g). In contrast, duodenum and MLN showed low colonization levels at the three necropsies, 0.6 ± 0.9 log10 CFU/g and 0.8 ± 0.9 log10 CFU/g, respectively. These levels were significantly lower than those of the ileum and tonsils whatever the necropsy date (p < 0.05).
CFU, colony-forming unit; MLN, mesenteric lymph nodes; n, number of pigs; SD, standard deviation.
Kinetics of individual seroconversion
Seroconversion (period of time during which antibodies develop and become detectable) occurred at different times for pigs. Antibody responses began by day 7 p.i. and OD% varied in mean from 7.2 ± 8.3 on day 7 to 73.0 ± 10.9 on day 84 p.i. (Table 1). All the G2 and G3 pigs had seroconverted by day 49 p.i. In G1, only one pig had seroconverted by day 7 p.i. and maintained a high OD% until necropsy (day 21 p.i.). In G2, two pigs out of eight had seroconverted by day 7 p.i., then, seven pigs by day 42 p.i., and the whole group by day 49 p.i. (day of necropsy). In G3, only two pigs had seroconverted by day 7 p.i. All the pigs in G3 had seroconverted by day 42 p.i., and antibody levels stayed high until necropsy (day 84 p.i.).
Genetic stability of strains after in vivo passage
During in vivo passage, three MLVA profiles were identified (Table 3). Among the 368 isolates, 351 belonged to MLVA Profile A like the inoculated strain (3-14-10-N-211), 11 to MLVA Profile B (3-14-11-N-211), and 1 to MLVA Profile C (3-14-9-N-211). Five isolates could not be typed.
MLVA, Multilocus Variable Number Tandem Repeat Analysis; NT, nontypable.
The difference between the three profiles lies in locus STTR6; this 6-bp locus was repeated 10, 9, or 11 times, respectively, for profiles A, B, and C (difference checked by sequencing the locus).
Profile A concerned 99.0% of strains on day 17 p.i., 95.8% of strains on day 42 p.i., and finally 92.9% of strains on day 84 p.i. Presence of profile B increased during the trial, 1.0%, 1.7%, and 5.7% of the strains on day 17, 42, and 84 p.i., respectively, and was found for: one isolate of feces of pig 5672, four isolates of feces (n = 2) and jejunum (n = 2) of pig 5671, four isolates from feces of pig 5658, and two isolates from cecum and ileum of this pig. The isolate with profile C was from feces of pig 5660. Other isolates from those same pigs (5672, 5671, 5658, 5660) had profile A in same or other types of samples.
Discussion
The main aim of this work was to describe the dynamics of colonization of pigs after inoculation with a monophasic variant of a Salmonella Typhimurium strain in experimental conditions. We inoculated pigs with a dose of 109 CFU because such high dose assures the uniformity of colonization (Boyen et al., 2009).
On day after inoculation, the rectal temperature of pigs briefly increased. This was already described in pigs experimentally infected with Salmonella Typhimurium (Uthe et al., 2007; Knetter et al., 2015).
We observed continuous excretion of Salmonella during the 84 days which peaked at day 3, and shedding patterns varied by individual pig. Most of studies reported intermittent excretion for Salmonella Typhimurium in experimental trials (Boyen et al., 2009; Ivanek et al., 2012) and in naturally contaminated pigs infected with Salmonella Typhimurium (Beloeil et al., 2003; Tessier, 2015) or monophasic Salmonella Typhimurium (Fernandes et al., 2016). Continuous shedding over a long time period may be explained by the high challenge dose used, as already reported, (Osterberg and Wallgren, 2008) or by the strain tested, a monophasic S.4,[5],12;i;-. A recent study showed that pigs exposed to an environment contaminated with a low dose of monophasic Salmonella Typhimurium can excrete up to 104 CFU/g (Lynch et al., 2017).
In our study, pigs in the first and second group had similar excretion patterns but not in the G3: excretion level of one pig was much higher compared with the other pigs. The carrier status of pigs with Salmonella Typhimurium has been widely described by many authors, and the existence of low, persistent (Knetter et al., 2015), and super shedders (Berriman et al., 2013) has been reported.
This study was also to establish the degree of contamination in organs and intestinal contents of pigs over time. Indeed, the tonsils, lymphatic nodes, and gastrointestinal tract are known to be the most important colonization sites for Salmonella Typhimurium and other serovars (Van Parys et al., 2011). At necropsy, 100% of the tonsils, ileum, and cecum were contaminated with whatever in all groups, tonsils being the most contaminated organ (5.0 log10 CFU/g), with a level higher than that obtained in another study for pigs challenged with 107 CFU of Salmonella Typhimurium (Boyen et al., 2009). Tonsils are one of the primary sites of Salmonella colonization in pigs that became subclinical carriers (Fedorka-Cray et al., 1995). The bacteria can subsequently spread to the retropharyngeal lymph nodes. Lymph nodes are a predominant colonization site of Salmonella Typhimurium in pigs and are used for diagnosis (Osterberg et al., 2010; Van Parys et al., 2013). However, a low colonization of lymph nodes was observed with our strain.
We found a high rate of colonization in jejunum, ileum, and cecum compared to duodenum. Our results are consistent with studies which established that Salmonella Typhimurium preferentially colonizes ileum, cecum, and colon, presumably due to low pH and fewer biliary salts than in the duodenum and jejunum (Uribe et al., 2016). These authors also reported that jejunum, ileum, and colon respond differently to Salmonella infection.
Our study showed that seroconversion occurred at different moments during the trial, from 7 days after infection for some pigs and for all the pigs in two groups after 49 days. However, in a recent study most of the pigs had seroconverted by 28 days after exposure of pigs to an environment contaminated with monophasic Salmonella Typhimurium (Lynch et al., 2017). Other authors with Salmonella Typhimurium reported 100% seroconversion 14 days after inoculation (Osterberg and Wallgren, 2008) or 77.8% 40 days after inoculation (Van Parys et al., 2013). A recent longitudinal survey carried out on 360 pigs from 3 farms allowed us to observe that pigs seroconverted on average at 137 ± 2.2 days of age and that by slaughter age, 75.07% of the pigs had seroconverted (Cevallos-Almeida, personal communication).
Our colonization and serological results indicate that the monophasic variant of Salmonella Typhimurium causes close reactions in pigs than those observed by other authors after inoculation with Salmonella Typhimurium. These two serovars are genetically close and have strong similarities in terms of pathogenicity island and virulence (Vieira-Pinto et al., 2012).
After in vivo passage in pigs, two new MLVA profiles appeared for 3.3% of the typed isolates. It results from insertion or deletion of one repeated unit in STTR6 locus, and this occurred in a short time frame. Other studies described genetic variation or mutation of human isolates of Salmonella Typhimurium after in vitro assay or in vivo passage in mice (Dimovski et al., 2014). Some studies reported multiple or single deletions in STTR5, STTR9, and STTR10 Salmonella MLVA loci (Dimovski et al., 2014; Vignaud et al., 2016). Dimovski et al. (2014) recommend that isolates with different profiles for one of the rapidly changing STTR5, STTR6, and STTR10 loci be grouped together into a same cluster. Thus, surveillance of outbreaks using this methodology has to use this clustering approach.
Conclusions
Our study highlighted a continuous and persistent excretion in pigs experimentally infected by monophasic variant of Salmonella Typhimurium, the tonsils being the most infected organs. Seroconversion was achieved by 49 days post-inoculation. In vivo passage resulted in low genetic variation of isolates at a single rapidly changing locus. Our findings are important for clarifying colonization in pigs by the monophasic variant of Salmonella Typhimurium.
Footnotes
Acknowledgments
The authors acknowledge funding by SENESCYT-ECUADOR (Secretaría de Educación Superior Ciencia y Tecnología Del Ecuador; Programa de Becas para Docentes Universitarios;
Disclosure Statement
No competing financial interests exist.