
Abstract
Escherichia coli O157:H7 is a human pathogen associated with gastrointestinal disease and hemolytic uremic syndrome. Direct contact with ruminants and their feces at agritourism or livestock interaction events is a known source of transmission. Footbath use is a pathogen reduction strategy that may decrease the transmission of E. coli O157:H7 at these interactions. The efficacy of chemical disinfectants in footbaths is not well reported. Our objective was to determine the susceptibility of E. coli O157:H7 toward commonly used disinfectants in vitro and within contaminated footbaths. The minimum inhibitory concentration and the minimum bactericidal concentration (MIC and MBC) and the time-to-kill were determined in vitro for seven E. coli O157:H7 strains using five disinfectant compounds (didecyldimethylammonium chloride [DDAC], glutaraldehyde, hydrogen peroxide, phenol, and sodium hypochlorite). Time–kill assays were performed within simulated footbaths at 22°C, 37°C, and 42°C with and without organic contamination using three commercial disinfectants with similar active ingredients (0.26% Clorox® Bleach, 0.034% Virex® II 256, and 1% Virkon™ S). The MBCs of disinfectants toward E. coli O157:H7 were 3.2, 625, 40, 5000, and 320 ppm of DDAC, glutaraldehyde, hydrogen peroxide, phenol, and sodium hypochlorite, respectively. At 2 × MIC, E. coli O157:H7 reached a 3 log10(colony-forming unit [CFU]/mL) reduction on contact with glutaraldehyde, by 20 min with DDAC and sodium hypochlorite, and by 4 h with phenol and hydrogen peroxide. In simulated footbaths, the commercial disinfectants reduced concentrations by 3 log10(CFU/mL) on contact in the absence of organic contamination, but viable E. coli O157:H7 was recovered from organically contaminated Clorox Bleach and Virex II 256 footbaths. No E. coli O157:H7 was recovered from the Virkon S footbaths after 10 min. This study highlights the ability for organic contamination to compromise the efficacy of disinfectants in footbaths and the importance of choosing an appropriate footbath disinfectant to retain the efficacy.
Introduction
E
Of major concern is the risk of infection to children who visit agritourism events with ruminants. Outbreaks of E. coli O157:H7 have been associated with agritourism events in many states across the United States (CDC, 2006; Saulo, 2013; Laughlin et al., 2016). Risk factors for direct transmission include exposed skin, poor hand hygiene, food and drinks in animal habitats, and fomites including strollers, bottles, pacifiers, or toys in animal habitats (CDC, 2018).
Biosafety and pathogen control reduction strategies are necessary to prevent E. coli O157:H7 infections at agritourism and livestock interaction events. Previous work has shown that hand hygiene was only practiced by 37% of observed visitors at petting zoos (Erdozain et al., 2012), suggesting that mitigation strategy alone is insufficient to protect against infection. Footbaths, which are disinfectant-filled mats or pans placed at the entrances and exits of animal habitats, may be used as a biosafety measure to reduce the risk of pathogen transmission at these events.
Footbaths are frequently used on farms and in the food industry to clean shoes and boots when leaving a soiled animal habitat. Several studies indicate that footbath use may decrease viable bacterial load on shoes and floors, although the efficacy is dependent on the type of disinfectant, the presence and amount of contamination, temperature, and exposure to sunlight (Morley et al., 2005; Dunowska et al., 2006; Allen et al., 2010; Hartmann et al., 2013; Hornig et al., 2016). General guidelines recommend that footbaths should be cleaned and refilled with freshly prepared disinfectant solution daily, and the footbath should be used when entering and exiting an animal habitat (Gernat, 2004). Recommended disinfectants include phenols, iodophors, hypochlorites, quaternary ammonium compounds (QACs), oxidizing agents, compound cresol, and formals (Jeffrey, 1997; Gernat, 2004).
Previous studies have shown that the efficacy of chlorine-, hydrogen peroxide-, and QAC-based disinfectants can be reduced in the presence of organic matter (Rodgers et al., 2003; Gehan et al., 2009; Shen et al., 2012). Therefore, we aimed to determine the survival of E. coli O157:H7 in vitro and within a simulated footbath with manure contamination to provide owners and operators of livestock interactions recommendations for their use.
Materials and Methods
Bacterial strains
E. coli O157:H7 isolates were previously obtained from goats at a processing plant (Jacob et al., 2013). Seven genetically distinct strains were selected to determine the minimum inhibitory concentration and the minimum bactericidal concentration (MIC and MBC) toward disinfectant compounds. Three strains (171F, 202H, and 208H) were selected for further in vitro time–kill analyses. Three additional strains (217C, 229F, and 230C) were made resistant to 50 μg/mL of nalidixic acid (Nalr) and used in simulated footbath time–kill analyses. The MIC of these Nalr strains was determined to be stable after induced resistance.
Determination of MIC and MBC
The MIC and MBC of five laboratory-grade disinfectants (didecyldimethylammonium chloride [DDAC; Sigma-Aldrich, St. Louis, MO], 25% glutaraldehyde [Fisher Chemical, Fair Lawn, NJ], 30% hydrogen peroxide [Fisher Chemical], phenol [Sigma-Aldrich], and 4% sodium hypochlorite [Sigma-Aldrich]) toward seven strains of E. coli O157:H7 were determined using microbroth dilution according to the Clinical and Laboratory Standards Institute guidelines (CLSI, 2015). Phenol and DDAC were dissolved in sterile deionized water to produce 8% and 1% solutions, respectively. The E. coli O157:H7 strains were streaked onto blood agar (Remel, Lenexa, KS) and incubated overnight at 37°C.
One colony was selected and inoculated into 2 mL of Mueller–Hinton Broth (MHB; Becton Dickinson, Sparks, MD) or M9 minimal media, prepared as previously described (Studier, 2005). These media were chosen to represent a high and low organic load. The MHB and M9 cultures were incubated at 37°C. Bacterial concentrations were periodically determined using the DensiCHEK plus (BioMerieux™), until a 0.5 McFarland Standard was achieved (∼1.5 h for MHB and 3 h for M9). The bacterial inoculum was added to twofold serial dilutions of each disinfectant. An uninoculated well with disinfectant served as a negative control. The MHB plates were incubated for 24 h at 37°C. M9 plates were incubated for 48 h at 37°C because of slower growth.
After incubation, turbidity was assessed visually, and the lowest concentration with no visible turbidity was determined to be the MIC. Twenty microliters of aliquots from the MIC well and one dilution higher were spotted three times on tryptic soy agar (TSA; Becton Dickinson). The TSA plates were incubated overnight at 37°C, and the growth was observed. The disinfectant concentration which reduced bacterial concentrations by 3 log10(colony-forming unit [CFU]/mL) or greater was determined to be the MBC. These assays were performed in triplicate.
In vitro time–kill assays
A colony from three E. coli O157:H7 strains was individually inoculated into 2 mL of MHB and incubated at 37°C until the cultures reached 0.5 McFarland. Forty microliters of each enrichment was diluted in 4 mL of MHB. Disinfectants were diluted in 1 mL of MHB to achieve concentrations of 1 × , 2 × , and 4 × the median MIC as determined for each disinfectant. One milliliter of the diluted culture was added to 1 mL of MHB with disinfectant so that the final disinfectant concentrations were 0.5 × , 1 × , and 2 × MIC, and the final bacterial concentration was ∼7.5 × 105 CFU/mL. The cultures were incubated at 37°C.
One hundred microliters of aliquots was removed at 0 min, 10 min, 20 min, 30 min, 1 h, 4 h, and 24 h. Aliquots were serially diluted 10-fold in sterile phosphate-buffered saline (PBS; Fisher BioReagents, Fair Lawn, NJ). Using the Miles and Misra method, 20 μL of each dilution was spotted onto TSA in triplicate and incubated overnight at 37°C. Colonies were counted to determine bacterial concentrations at each time point.
Manure source for simulated footbaths
Approximately 20 g of fresh goat feces was collected and determined to be E. coli O157:H7 negative before use within a simulated footbath. Briefly, the feces were homogenized. One gram was added to 9 mL of Gram-negative broth (Becton Dickinson) supplemented with 0.5 mg/L cefixime, 10 mg/L cefsulodin, and 8 mg/L vancomycin and incubated for 6 h at 37°C. Immunomagnetic bead separation was performed using 20 μL of anti-E. coli O157 dynabeads (Applied Biosystems, Thermo Fisher Scientific, Foster City, CA) according to the manufacturer's instructions. Beads were washed three times in 100 μL of sterile PBS, and a 50 μL aliquot was spread on sorbitol MacConkey agar (Criterion; Hardy Diagnostics, Santa Maria, CA) supplemented with 0.5 μg/mL cefixime and 2.5 μg/mL tellurite (CT-SMAC) and incubated overnight at 37°C. The plates were free of gray colonies and feces confirmed to be negative before use.
Footbath time–kill assays
A rubber footbath (Wearwell™, Smyrna, TN) was cut into 9 × 12 cm pieces and laid into empty 9 × 12 cm plastic boxes to simulate a footbath. One gram of pine wood shavings (Tractor Supply Company, Brentwood, TN) was added to half of the footbaths, and all footbaths were autoclaved to sterilize. Two grams of goat feces were added to each footbath with pine wood shavings.
One colony from each of three strains of Nalr E. coli O157:H7 strains were individually inoculated into 17 mL of MHB and incubated at 37°C to 0.5 McFarland. Footbaths were prepared either by adding 100 mL of sterile deionized water to empty footbaths or footbaths with pine shavings and manure or by adding 100 mL of sterile deionized water with disinfectant to empty footbaths or footbaths with pine shavings and manure. The footbaths with disinfectant had one of three commercially available disinfectants at the manufacturer's recommended concentration (0.26% Clorox® Bleach [Clorox Professional Products Company, Oakland, CA], 0.034% Virex® II 256 [Diversy, Charlotte, NC], and 1% Virkon™ S [The Chemours Company, Wilmington, DE]). The footbaths were incubated at 22°C, 37°C, and 42°C for 1 h to equilibrate the temperature of the water in the footbaths. Temperatures were chosen to represent footbath conditions on mild, warm, and hot days when exposed to direct sunlight.
One milliliter of bacterial culture was added to each footbath. After manually mixing, 100 μL aliquots was taken at 0 min, 15 min, 30 min, 1 h, 4 h, and 24 h, added to 900 μL of Dey–Engley Neutralizing Broth (D/E broth; Criterion; Hardy Diagnostics), and incubated at room temperature for 15 min to neutralize the disinfectant compound and prevent further killing of the bacteria. The samples were serially diluted 10-fold, plated onto CT-SMAC supplemented with 50 μg/mL of nalidixic acid (CTN-SMAC), and incubated overnight at 37°C. Gray colonies were counted to determine the concentration of E. coli O157:H7 at each time point. If no colonies were present, a 10 μL aliquot of the D/E enrichment was spotted on CTN-SMAC to determine if the bacterial concentration was below the limit of detection.
Study analysis
The median, mean, and standard deviation were calculated for the MIC and MBC measures. The median MIC was used for time–kill assays because MICs are measured in intervals. For the time–kill assays, the concentrations (CFU/mL) of each strain were averaged at each time point to produce a mean CFU/mL value and transformed to log10(CFU/mL). The bactericidal potential of each disinfectant was evaluated by its ability to reduce the concentration of E. coli O157:H7 by 3 log10(CFU/mL), or a 99.9% reduction, as per CLSI guidelines (CLSI, 2015). In some cases where bacterial concentrations decreased by 3 log10(CFU/mL), but bacteria were still recovered, the bacteria were considered substantially reduced but viable. For the footbath time–kill assays, in cases where bacterial concentrations were reduced below the limit of detection (1.67 × 102 CFU/mL), but viable bacteria were still recovered from the enriched aliquots, the bacterial concentration was considered to be the lowest concentration at which CFUs could be detected.
Results
MICs and MBCs
Strains behaved similarly in their MICs and MBCs toward the five laboratory-grade disinfectant compounds. The median and mean MIC and MBC values for each disinfectant are presented in Table 1. In the MHB, DDAC and hydrogen peroxide had the lowest median MIC and MBC (3.2 and 40 ppm, respectively), while phenol had the highest (MIC 2500 ppm and MBC 5000 ppm). The median MIC and MBC of glutaraldehyde, hydrogen peroxide, and sodium hypochlorite was 625, 40, and 320 ppm, respectively. When the assay was repeated using M9, the median MIC and MBC of DDAC, glutaraldehyde, and sodium hypochlorite was 1.6, 5, and 1.25 ppm, respectively. The median MIC of hydrogen peroxide and phenol was 10 and 1250 ppm, while their MBCs were 20 and 2500 ppm, respectively.
Median and Mean Minimum Inhibitory Concentrations (ppm) and Minimum Bactericidal Concentrations of Laboratory-Grade Disinfectant Compounds for Escherichia coli O157:H7 Grown in Mueller–Hinton Broth and M9 Minimal Media
Seven strains of E. coli O157:H7 at ∼5.9 log10(CFU/mL) were exposed to twofold dilutions of each disinfectant compound in MHB or M9. Cultures were incubated for 24 h in MHB and 48 h in M9, and MICs and MBCs were observed. The assay was repeated in triplicate, and the median MIC and MBC of each disinfectant toward all strains is reported.
The lowest concentration at which no visible growth was observed was determined to be the MIC.
Concentrations at which no visible growth was observed were spotted in triplicate on tryptic-soy agar, and the lowest concentration at which a 3 log10(CFU/mL) reduction was observed was determined to be the MBC.
CFU, colony-forming unit; MBC, minimum bactericidal concentration; MHB, Mueller–Hinton Broth; MIC, minimum inhibitory concentration; SD, standard deviation.
In vitro time–kill assays
As expected, 0.5 × MIC of all disinfectants did not reduce E. coli O157:H7 concentrations by 3 log10(CFU/mL) (Fig. 1). At 1 × and 2 × MICs, the concentrations of E. coli O157:H7 were either static or reduced by 3 log10(CFU/mL) within 24 h for all disinfectants.
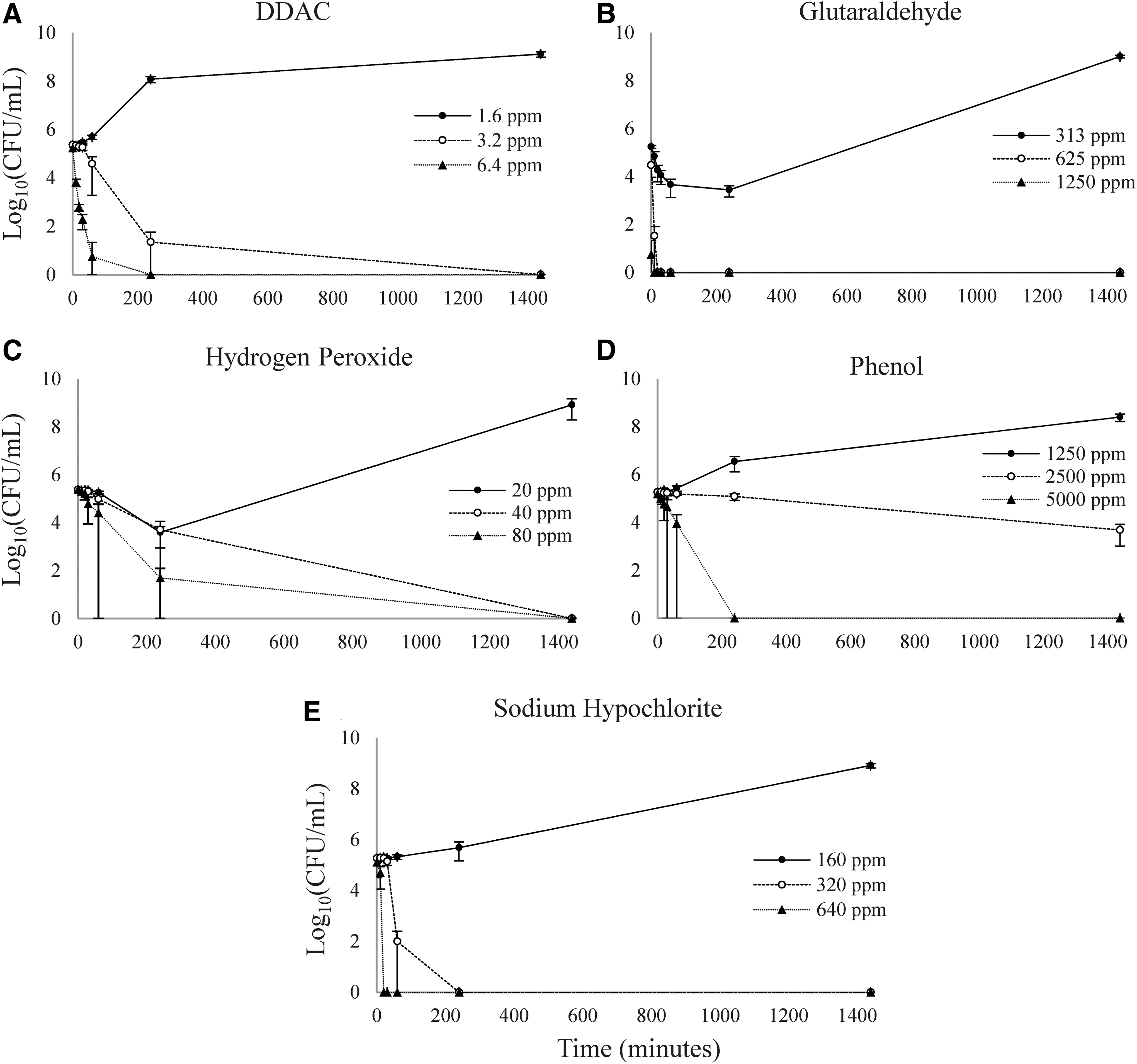
Viable Escherichia coli O157:H7 concentrations over time when exposed to five disinfectant compounds at different concentrations. E. coli O157:H7 was exposed to 0.5 × (filled circle, solid line), 1.0 × (open circle, dashed line), and 2 × MIC (filled triangle, dotted line) of didecyldimethylammonium chloride
At 1.6 ppm of DDAC, E. coli O157:H7 reached a final concentration of 9.1 log10(CFU/mL) by 24 h, whereas at 3.2 and 6.4 ppm, the concentrations were reduced by 3 log10(CFU/mL) by 4 h and 20 min, respectively.
At 313 ppm of glutaraldehyde, concentrations initially decreased for the first hour but then increased to 9.0 log10(CFU/mL) by 24 h. At 625 ppm of glutaraldehyde, E. coli O157:H7 concentrations were reduced by 3 log10(CFU/mL) within 10 min, and at 1250 ppm, the bacteria were eliminated on contact.
At 20 ppm of hydrogen peroxide, concentrations decreased for the first 4 h but then increased to a final concentration of 8.9 log10(CFU/mL). A 3 log10(CFU/mL) reduction was achieved by 24 h at 40 ppm and by 4 h at 80 ppm of hydrogen peroxide.
When exposed to phenol, E. coli O157:H7 increased in concentration at 1250 ppm [final concentration of 8.4 log10(CFU/mL)], remained stable at 2500 ppm [final concentration of 3.7 log10(CFU/mL)], and achieved a 3 log10(CFU/mL) reduction by 4 h at 5000 ppm.
At 160 ppm of sodium hypochlorite, E. coli O157:H7 reached a final concentration of 8.9 log10(CFU/mL). A 3 log10(CFU/mL) reduction was reached by 1 h of exposure at 320 ppm and by 20 min at 640 ppm sodium hypochlorite.
Footbath time–kill assays
The concentration of E. coli O157:H7 strains incubated in a footbath assay without manure or disinfectant decreased to 4.1, 3.5, and 2.5 log10(CFU/mL) at 4 h for 22°C, 37°C, and 42°C, respectively, and below the detection limit by 24 h (Fig. 2A). E. coli O157:H7 concentrations in the manure-contaminated footbaths held at 37°C and 42°C increased by ∼1.5 log10(CFU/mL) over 24 h, whereas concentrations at 22°C increased by ∼0.5 log10(CFU/mL). When each disinfectant was added to footbaths without organic contamination, the E. coli O157:H7 concentrations were reduced to undetectable levels on contact with no recovery upon enrichment, regardless of the type of disinfectant or the incubation temperature of the footbath (data not shown).
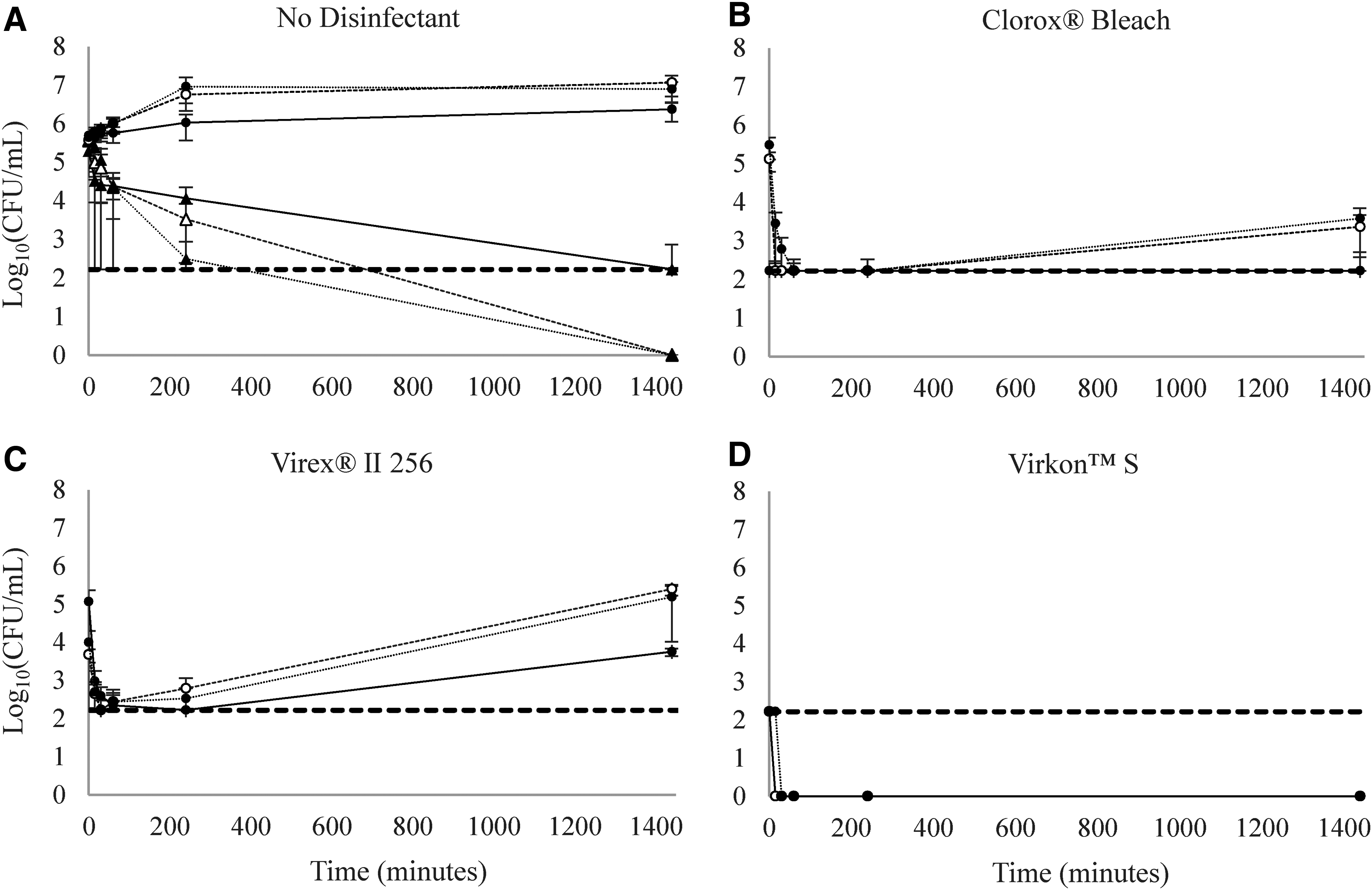
Viable Escherichia coli O157:H7 concentrations over time in simulated footbaths. Footbaths with no disinfectant
In the organically contaminated footbaths with Clorox Bleach at 22°C, E. coli O157:H7 was reduced by 3 log10(CFU/mL) on contact, although viable organisms were recovered upon enrichment (Fig. 2B). At 37°C and 42°C, E. coli O157:H7 concentrations reached a 3 log10(CFU/mL) reduction by 15 and 30 min, respectively; yet upon enrichment, the footbaths were still positive for viable organisms. By 24 h of exposure, E. coli O157:H7 reached 3.4 and 3.6 log10(CFU/mL) at 37°C and 42°C, respectively (Fig. 2B). Concentrations remained below the limit of detection at 22°C, with recovery upon enrichment at 24 h.
At all temperatures in the Virex II 256 footbaths with organic contamination, E. coli O157:H7 concentrations were reduced by at least 3 log10(CFU/mL) within 15 min; however, viable organisms were recovered after enrichment (Fig. 2C). E. coli O157:H7 concentrations began increasing by 24 h, reaching 3.8, 5.4, and 5.2 log10(CFU/mL) at 22°C, 37°C, and 42°C, respectively.
At all temperatures in the organically contaminated footbaths with Virkon S, E. coli O157:H7 concentrations were reduced by at least 3 log10(CFU/mL) on contact (Fig. 2D). Viable E. coli O157:H7 were recovered upon enrichment at 0 min at 22°C and 37°C, and at 15 min at 42°C footbaths, but no E. coli O157:H7 was recovered at any other time point.
Discussion and Conclusions
Results from our study support high disinfectant efficacy at killing E. coli O157:H7 in vitro. Five classes of disinfectant compounds reduced the E. coli O157:H7 concentration by 3 log10(CFU/mL) within 4 h of exposure at 2 × MIC, well below the expected concentration in commercial products. However, the efficacy was variable in footbaths amended with manure and pine shavings. Incubation time, temperature, and the disinfectant influenced the concentration of E. coli O157:H7. These results suggest that E. coli O157:H7 may survive in used footbaths at livestock interactions, causing the footbath to be a potential source of contamination.
Previous studies have shown that E. coli O157:H7 has an MIC50 of 2 μg/mL and an MIC90 of 4 μg/mL when treated with DDAC, one of the most common QACs used in food processing facilities and clinical environments (Bloomfield, 2002; Tezel et al., 2007; Beier et al., 2012; Melin et al., 2014). We report similar inhibitory concentrations, with MICs for DDAC reported at 3.2 ppm (3.2 μg/mL) in MHB and 1.6 ppm (1.6 μg/mL) in M9 (Table 1).
However, our results suggest that E. coli O157:H7 may survive within footbaths containing manure and filled with Virex II 256, which uses DDAC as an active ingredient. A 3 log10(CFU/mL) reduction was achieved within 15 min, but viable E. coli O157:H7 were recovered after 24 h (Fig. 2C). This reflects previous studies, which demonstrated little to no decrease in the concentration of bacteria recovered from the sole of boots when boots are dipped in footbaths containing QAC-based disinfectants (Morley et al., 2005; Hornig et al., 2016). Taking this into consideration with our findings, these disinfectants should not be recommended for use in footbaths.
Glutaraldehyde has been demonstrated to quickly and effectively eliminate E. coli O157:H7 (Sagripanti et al., 1997). At 1 × and 2 × MICs, glutaraldehyde rapidly killed the E. coli O157:H7 in vitro, eliminating the pathogen within minutes (Fig. 1B). The MIC of glutaraldehyde was appreciably lower in M9 versus MHB. This could have been due to an interaction of the disinfectant with organic compounds, antioxidants, reducing agents, or other constituents in the media, as a previous work has demonstrated that glutaraldehyde irreversibly binds to and denatures proteins and nucleic acids (Maris, 1995).
We suspect that the efficacy of a glutaraldehyde-based disinfectant would be decreased in a footbath contaminated with organic matter, although we did not evaluate a commercial product containing glutaraldehyde as an active ingredient because these are not as commonly used as QAC-, chlorine-, and peroxygen-based disinfectants. Similarly, we did not evaluate phenol products as previous studies suggested that they tend to be ineffective at reducing bacterial load on boots (Hartmann et al., 2013; Hornig et al., 2016). At 1.0 × MIC of phenol, the concentration of E. coli O157:H7 remained fairly stable, indicating the static activity.
Hydrogen peroxide has been effective in reducing pathogen concentrations (Materon, 2003; Fouladkhah and Avens, 2010). When exposed to 0.5 × and 1 × MICs of hydrogen peroxide, the bacterial concentrations were similarly reduced for 4 h; however, after 4 h at 0.5 × MIC, the concentration began to increase (Fig. 2C). We suspect that the hydrogen peroxide was neutralized to subinhibitory concentrations, although further work is needed to confirm this hypothesis. When exposed to the peroxygen-based disinfectant Virkon S in the organically contaminated footbaths, E. coli O157:H7 concentration was reduced by 3 log10(CFU/mL) on contact and viable organisms were not recovered (Fig. 2D).
This disinfectant was the most efficacious at preventing E. coli O157:H7 survival within footbaths containing manure. Peroxygen-based disinfectants consistently rank as one of the most effective footbath disinfectants, reducing concentrations on boot soles by 1.3–1.4 log CFU/cm2 (Morley et al., 2005; Dunowska et al., 2006; Hornig et al., 2016). As evidenced by our results, Virkon S is a practical disinfectant that can be used to prevent the survival of E. coli O157:H7 within footbaths.
The presence of organic matter, high temperatures, and light exposure have been shown to decrease the activity of chlorine-based disinfectants (Rodgers et al., 2003; Shen et al., 2012). We found that the MIC of sodium hypochlorite toward E. coli O157:H7 was appreciably lower in M9 minimal media than in the MHB (Table 1). It is suspected that the concentration of active disinfectant was decreased by protein in the MHB. This hypothesis is supported by our simulated footbath, where Clorox Bleach solution in footbaths with manure led to a 3 log10(CFU/mL) reduction by 30 min at each temperature, but viable E. coli O157:H7 was still recovered upon enrichment (Fig. 2B). Because it took longer for the E. coli O157:H7 concentration to be reduced at 42°C than at 22°C or 37°C, and E. coli O157:H7 was recovered upon enrichment, we suspect that the increased temperature decreased the activity of the disinfectant and the organic matter decreased the active concentration of the disinfectant available for biocidal activity.
In our study, footbath environments were simulated in plastic boxes. Although we do not expect this to alter the results of our study, these surfaces would not be present in an actual rubber footbath. Additionally, footbaths and in vitro cultures were incubated without the presence of light, which could affect the efficacy of the disinfectants. If deactivated by light, our results would overestimate the efficacy of manure-amended footbaths. Light is a reasonable consideration for footbath use at livestock interactions.
We chose to report prepared disinfectant concentrations as recommended by the manufacturer, and we did not directly measure actual disinfectant concentrations throughout the assays. We expect that active disinfectant concentrations in footbaths will decrease over time, particularly at higher temperatures and in the presence of organic matter, which may negatively impact the efficacy. This should have also occurred in our simulations, so it is not expected to radically impact our results.
The amount of organic matter in the footbaths at agritourism events will vary depending on the frequency of cleaning and the amount of traffic the footbath receives. If footbaths contain a greater concentration of organic matter than what was used in this study, disinfectant efficacy could be further reduced. We did not directly measure the total organic content or chemical oxygen demand of the footbath water amended with manure in this study; however, enhancements for future studies would determine the maximum organic content at which disinfectants are still active in footbaths. Certainly, the practicality of maintaining low levels of organic contamination with multiple users is challenging and should be considered.
We evaluated disinfectants in footbaths at moderate (22°C), warm (37°C), and hot (42°C) temperatures, which would be expected for footbaths exposed to direct sunlight in summer months. Previous studies have shown that QAC- and chlorine-based disinfectants remain viable for longer periods at lower temperatures (4°C), although the disinfectants are not as active against microorganisms (Gelinas et al., 1984; Piskin and Turkun, 1995; Jang et al., 2014). Considering the limited effect the temperature had on the survival of E. coli O157:H7 in footbaths in this study, we would expect to similarly find E. coli O157:H7 survival at lower temperatures, but further work is needed to confirm this hypothesis.
This study and others have demonstrated that, while footbaths may decrease viable bacterial concentrations, they should not be used as a sole prevention method for complete elimination of potential pathogens (Morley et al., 2005; Dunowska et al., 2006; Hartmann et al., 2013; Hornig et al., 2016). Additionally, E. coli O157:H7 was shown to have survived in Clorox Bleach- and Virex II 256-filled footbaths that were contaminated with organic matter after 24 h, suggesting that they may actually be a source of contamination to future footbath users.
Of the three evaluated disinfectants, Virkon S was the most effective at preventing E. coli O157:H7 survival within footbaths. If footbaths are to be used as a biosafety measure, Virkon S or similar peroxygen-based disinfectants would be the most appropriate of the disinfectants we evaluated. Additional recommendations should include that footbaths be cleaned regularly or as soon as it is apparent that organic matter is accumulating. The practicality of this recommendation is of concern. If Virkon S or a similar peroxygen-based disinfectant is used, it may be acceptable to clean the footbath every 24 h to retain the efficacy.
Footnotes
Acknowledgments
This study was funded by the North Carolina State University of Veterinary Medicine (CVM) intramural funds. We thank Dr. Derek Foster for his assistance with this project.
Disclosure Statement
No competing financial interests exist.