
Abstract
This study was intended to investigate the effect of contact with concentrations close to the minimum inhibitory concentration (MIC) (0.5, 1, and 1.5 MIC; MIC of planktonic cells was determined using a microdilution broth method) of sodium hypochlorite (SHY) or benzalkonium chloride (BZK) during the process of formation of biofilm (24 h), upon the architecture and viability of the biofilms formed by four L. monocytogenes isolates of molecular serotype 1/2a: S2-1 (BZK-susceptible strain; MICBZK = 3.0 ppm), S2-2 (BZK-resistant strain qacH positive; MICBZK = 13 ppm), CDL 69 (BZK-resistant strain bcrABC positive; MICBZK = 10 ppm), and S2BAC (BZK-resistant laboratory mutant of S2-1, with multidrug resistance phenotype; MICBZK = 9 ppm). Images were examined through confocal laser scanning microscopy after staining with SYTO 9 and Propidium Iodide. Biovolume values in the observation field (14,161 μm2) in the absence of biocides ranged from 103,928.3 ± 6,730.2 μm3 (S2BAC) to 276,030.9 ± 42,291.9 μm3 (S2-1). Exposure to SHY at 0.5 MIC reduced (p < 0.05) the biovolume of biofilms formed by S2-1 and S2BAC and did not modify (p > 0.05) the biovolume of biofilms by S2-2 and CDL 69. Exposure to sub-MICs of BZK decreased (p < 0.05; S2-1) or enhanced (p < 0.05; S2-2, CDL 69 and S2BAC) biofilm development. Exposure to biocides at 1 or 1.5 MIC inhibited biofilm formation. This study provides clear evidence that BZK at sub-MICs can enhance the biofilm-forming ability of BZK-resistant L. monocytogenes strains. Because biofilms contribute to the persistence of bacteria throughout the food chain and represent a major source of food contamination, our findings suggest the importance of avoiding sub-MICs of disinfectants in food-handling environments.
Introduction
L
Cells in biofilms display increased resistance to environmental challenges, and as a consequence are difficult to eradicate in the food industry. Several compounds are commonly used as antimicrobials for sanitizing surfaces in food contact surfaces (Buzón-Durán et al., 2017). Chlorine-based disinfectants, such as sodium hypochlorite (SHY), are oxidizing compounds showing a broad-spectrum bactericidal activity, high efficacy, and low cost (Waghmare and Annapure, 2015). Quaternary ammonium compounds, such as benzalkonium chloride (BZK), are cationic surfactants that act by disrupting lipid membrane bilayers, being effective against a number of pathogenic microorganisms, especially Gram-positive bacteria (Henriques and Fraqueza, 2017).
An inappropriate use of biocides (e.g., incorrect doses, unsuitable storage, or insufficient cleaning before disinfection), as well as the use of disinfectants at points where access by such chemicals is restricted and bacterial cells may find shelter (e.g., the inner surfaces of pipes), may cause exposure of bacteria to subminimum inhibitory concentrations (MICs) (Capita et al., 2014). Previous studies assessing biofilm structures through confocal microscopy have shown that exposure to sub-MICs of some food-grade biocides enhances biofilm production by Salmonella, Escherichia coli, or methicillin-resistant Staphylococcus aureus (Capita et al., 2014, 2017; Buzón-Durán et al., 2017). However, it would seem that reports quantitatively assessing the effect of sub-MICs of SHY and BZK on structural parameters (e.g., biovolume, surface coverage, maximum thickness, or roughness) and cellular viability of L. monocytogenes biofilms are lacking. Thus, this study was intended to evaluate, through confocal laser scanning microscopy (CLSM), the effects upon the architecture and viability of the biofilms formed by four L. monocytogenes strains, arising from the use at low concentrations (0.5, 1, and 1.5 MIC) of two disinfectants commonly employed in food-processing premises (SHY and BZK).
Materials and Methods
Bacterial strains
Four L. monocytogenes strains of molecular serotype 1/2a were tested. Three strains were isolated from a pork processing plant (Ortiz et al., 2014, 2016; López-Alonso et al., 2015): S2-1 (BZK-susceptible strain), S2-2 (BZK-resistant strain harboring the qacH gene of the Tn6188 transposon), and S2BAC (BZK-resistant laboratory mutant of S2-1, with a multidrug resistance (MDR) phenotype due to the overexpression of endogenous efflux pumps after in vitro exposure to quaternary ammonium compounds). A BZK-resistant L. monocytogenes strain isolated from smoked salmon and harboring the bcrABC resistance cassette was also tested (strain CDL 69; Müller et al., 2013). Strains were maintained in Tryptic Soy Broth (TSB; Oxoid Ltd., Hampshire, England) supplemented with 20% (v/v) glycerol at −80°C. Before experiments, frozen cells were subcultured twice in TSB at 37°C. Working cultures were kept at 4°C on plates of Tryptic Soy Agar (TSA; Oxoid).
Disinfectants
Two biocides were tested: SHY (10% active chlorine; Sigma-Aldrich, Steinheim, Germany) and BZK (Fluka, Deisenhofen, Germany). Sterile solutions were prepared in distilled water immediately before each experiment.
Determination of MICs
Five colonies of each strain were taken from TSA plates, and subcultured twice in 10 mL of TSB at 37°C for 18 h. The concentration of bacterial cells after incubation was estimated in two ways: by enumerating viable cells in Plate Count Agar (PCA; Oxoid) in duplicate, using 0.1% (wt/vol) peptone water (Oxoid) for decimal dilutions, and counting visible colonies after 48 h of incubation at 37°C, and by measuring the optical densities (ODs) using a wide band filter (band area 420–580 nm; OD420–580) in an automated turbidimetric-based system Bioscreen C MBR (Oy Growth Curves Ab Ltd., Helsinki, Finland). These experiments had shown that such cultures contain, as average, 9.48 ± 0.15 log10 colony-forming units (CFU)/mL, and have an OD of 0.666 ± 0.015.
Three decimal dilutions were performed in TSB. Individual wells of 100-well polystyrene microplates (Oy Growth Curves Ab) were filled with 20 μL of the chemical solutions, a range of concentrations being used for each disinfectant (from 250 to 5000 ppm, at intervals of 250 ppm, for SHY, and from 1 to 20 ppm, at intervals of 1 ppm, for BZK), and 180 μL of the third dilution of this bacterial culture. This gave a final concentration in the well of ∼6.5 log CFU/mL. The inoculum concentration was confirmed by plating. The microplates were incubated at 37°C in a Bioscreen C MBR (Oy Growth Curves Ab). Positive controls (200 μL of inoculum at 6.5 log CFU/mL) were included in each experiment, as were negative controls, which consisted of 180 μL of TSB and 20 μL of chemical solution. The experiments were replicated five times on separate days. The MIC was established as the lowest biocide concentration necessary to prevent growth after 48 h of incubation. An OD420–580 of 0.200 was considered the cutoff for bacterial growth (Alonso-Calleja et al., 2015).
Biofilm formation and analysis
An analysis of the structure of the biofilm was undertaken using the method previously described by Capita et al. (2014), with some modifications. Briefly, five colonies of each strain were taken from TSA plates, and subcultured twice in 10 mL of TSB at 37°C for 18 h. Appropriate (two-fold) dilutions in TSB were made to obtain a concentration of ∼7.5 log CFU/mL. A volume of 250 μL was added to the wells of Nunc™ MicroWell™ 96-Well Optical-Bottom Plates with Polymer Base (Thermo Fisher Scientific, New Hampshire, England), having high optical quality, low fluorescent background, and overall flatness, which allowed high-resolution imaging.
After 1 h of adhesion at 37°C without aeration, the wells were rinsed with 150 mM of NaCl to eliminate any nonadherent bacteria before being refilled with 250 μL of TSB or TSB with disinfectant (SHY or BZK at concentrations of 0.5, 1, and 1.5 MIC were used). After incubation for 24 h at 37°C, the wells were rinsed with 150 mM of NaCl. For staining with fluorescent dyes, a volume of 2 μL of a 1:1 mixture of SYTO 9 (stock 3.34 mM in dimethyl sulfoxide [DMSO]) and Propidium Iodide (PI; stock 20 mM in DMSO) fluorescent dyes from the BacLight Viability Kit (Invitrogen, Carlsbad, CA) was added to 1000 μL of TSB, and 250 μL of this solution was put into each well. The plate was then incubated in the dark at 37°C for 20 min to enable fluorescent labeling of the bacteria. To determine the number of cells adhering to the polystyrene before biofilm formation, one well in each experiment was stained immediately after adhesion and rinsing.
CLSM image acquisition was performed using a Nikon Eclipse TE 2000-U confocal laser scanning microscope with EZ-C13.60 software (Nikon Instruments, Inc., New York). The biofilms were observed with a forty magnification (40 × ) lens. Fluorescence was detected by excitation at 488 nm (argon laser), and emissions were collected with a 590/50 (SYTO 9) or a 650LP (PI) bandpass filter. Three stacks of horizontal plane images (512 × 512 pixels corresponding to 119 × 119 μm) with a z-step of 1 μm, were acquired for each biofilm from three different areas in the well. Three independent experiments were performed for each strain and chemical treatment on different days. Thus, in total, 126 CLSM images were obtained. Image analysis was carried out as previously described (Rodríguez-Melcón et al., 2018) using the IMARIS 9.1 software package (Bitplane, Zurich, Switzerland) for reconstructions of three-dimensional projections of the structure of the biofilms.
Statistical analysis
The structural parameters of biofilms were compared for statistical significance using one-way analysis of variance techniques. Mean separations were obtained using Duncan's multiple range test. A correlation analysis was performed to determine the relationship between the different structural parameters of the biofilms. Data were processed using the Statistica® 8.0 software package (Statsoft Ltd., Tulsa, OK).
Results
Effect of disinfectants used at 0.5 MIC
Using a microdilution broth method, the MIC value of SHY for L. monocytogenes planktonic cells was 3500 ppm (350 ppm of free chlorine). MICs for BZK were 3 ppm (S2-1), 13 ppm (S2-2), 10 ppm (CDL 69), or 9 ppm (S2BAC). To determine whether 0.5 MIC of SHY or BZK would influence biofilm production, the structural parameters (biovolume, surface coverage, maximum thickness, and roughness) and cellular viability of biofilms formed by L. monocytogenes cells in the absence of biocides (control) or in the presence of SHY at 1750 ppm and BZK at 1.5 ppm (S2-1), 6.5 ppm (S2-2), 5 ppm (CDL 69), or 4.5 ppm (S2BAC) was studied by CLSM.
When biofilms were grown in the absence of biocides, L. monocytogenes cells produced dense and homogenous structures that covered more than 90% of the surface available. The highest (p < 0.05) biovolume in the observation field (14,161 μm2) was observed for biofilms formed by strain S2-1 (276,030.9 ± 42,291.9 μm3). Strain S2-2 showed a biovolume of 224,576.0 ± 5,362.3 μm3, which was higher than those observed for CLD 69 (129.028,1 ± 58,348.9 μm3) and for S2BAC (103,928.3 ± 6,730.2 μm3) (Fig. 1). Figure 2 shows the percentage of surface coverage, maximum thickness, and roughness for L. monocytogenes biofilms.
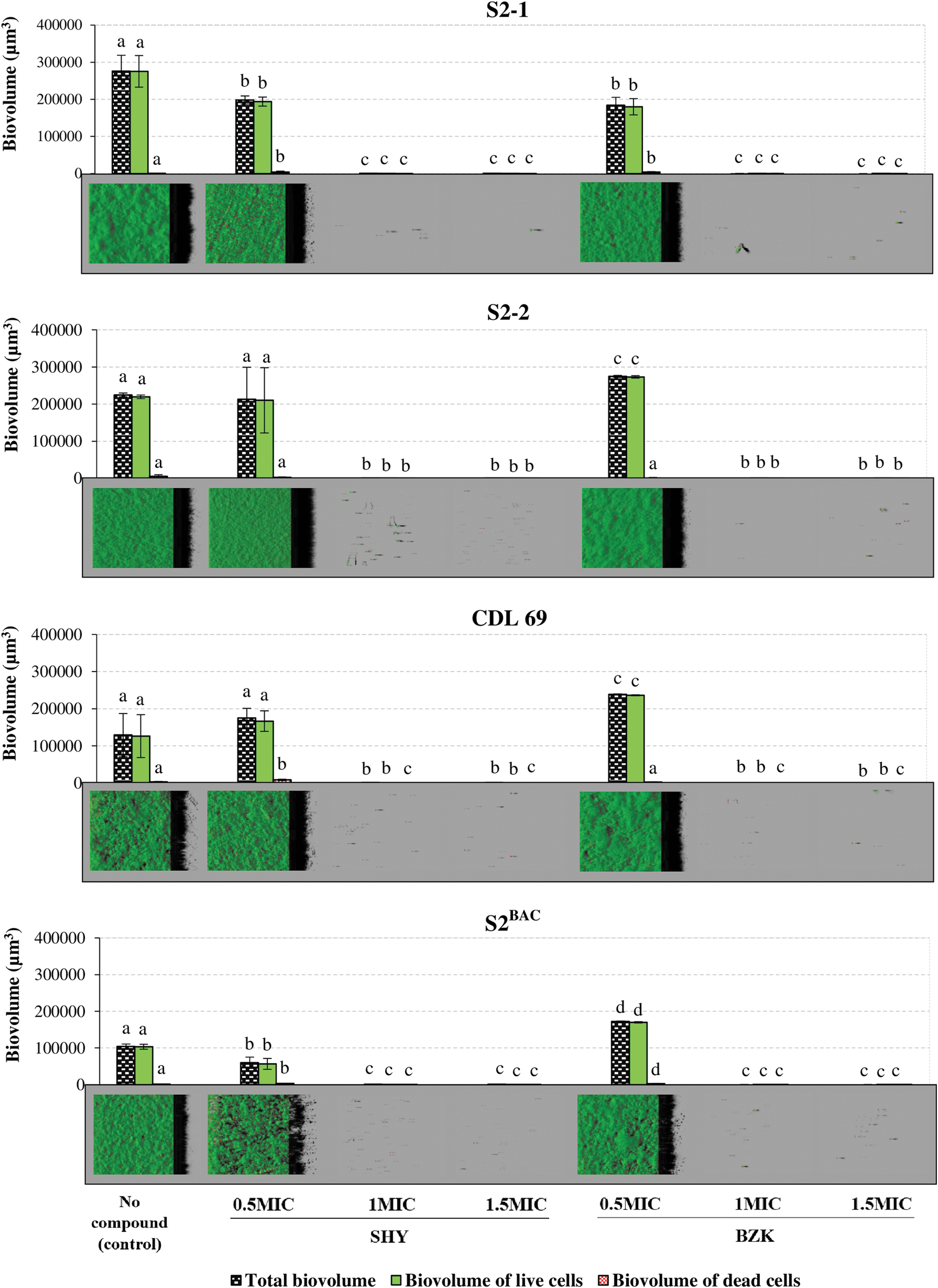
Biovolume in the observation field (14,161 μm2) of 24-h-old biofilms formed by four Listeria monocytogenes strains of molecular serotype 1/2a (S2-1, S2-2, CDL 69, and S2BAC) in the absence of disinfectants (control) or in the presence of SHY or BZK at 0.5, 1, or 1.5 MIC. Bars in the same graphic for the same physiological state (total cells, live cells, or dead cells) without any letter in common are significantly different (p < 0.05). The biofilms were labeled in green with SYTO 9 and in red with Propidium Iodide. The images correspond to three-dimensional reconstructions obtained from confocal stack images by the IMARIS 9.1 software, virtual projections being included on the right. SHY, sodium hypochlorite; BZK, benzalkonium chloride; MIC, minimum inhibitory concentration.
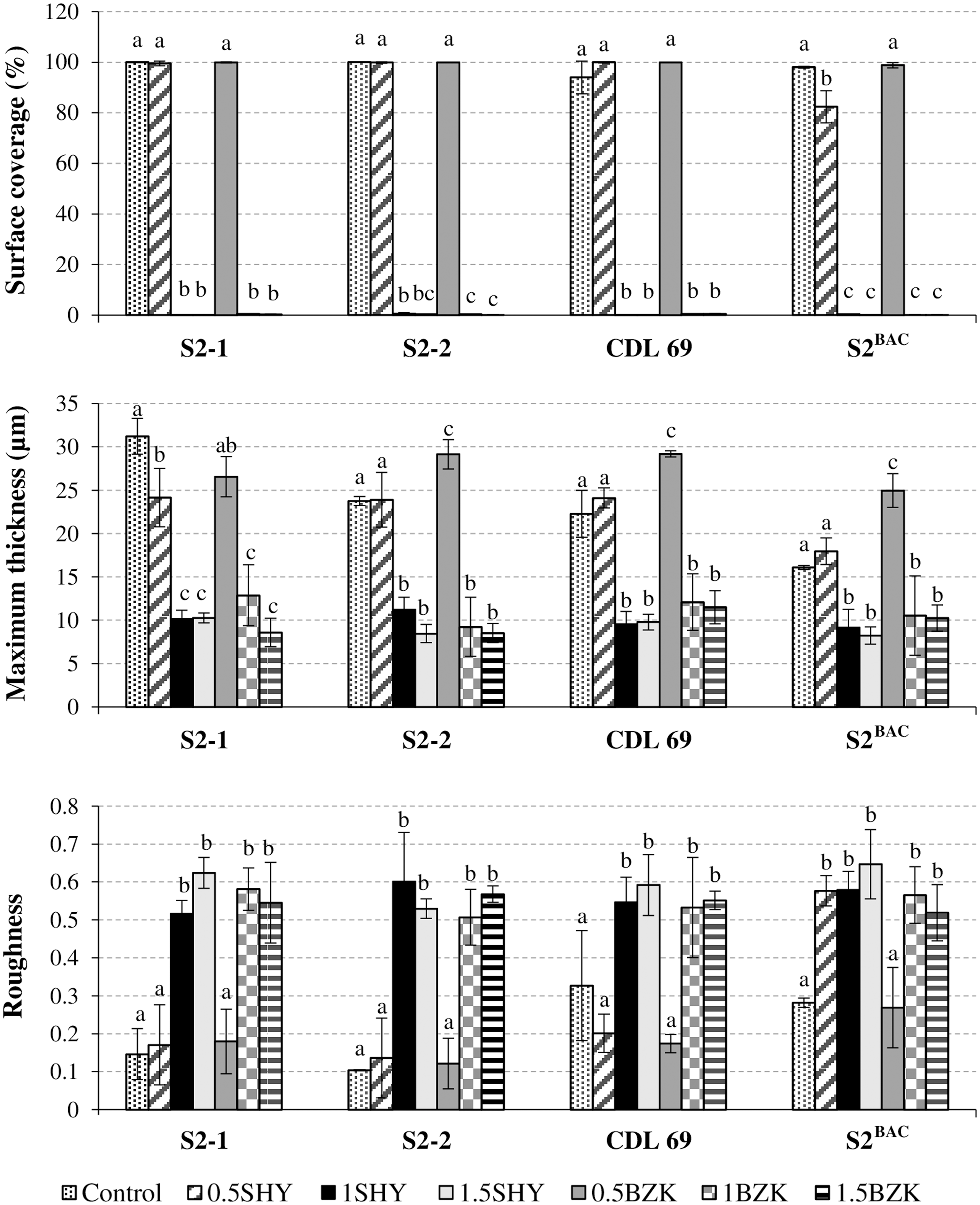
Percentage of surface coverage, maximum thickness, and roughness of 24-h-old biofilms formed by four L. monocytogenes strains of molecular serotype 1/2a (S2-1, S2-2, CDL 69, and S2BAC) in the absence of disinfectants (control) or in the presence of SHY or BZK at 0.5, 1, or 1.5 MIC. Bars in the same graphic for the same strain (S2-1, S2-2, CDL 69, or S2BAC) without any letter in common are significantly different (p < 0.05).
Cultures exposed to SHY at 0.5 MIC produced biofilms with similar (S2-2 and CLD 69) or lower (p < 0.05; S2-1 and S2BAC) biovolume than those from unexposed cultures (control) (Fig. 2); data for biovolume were 198,044.7 ±11,072.2 μm3 (S2-1), 212,619.9 ± 86.675,0 μm3 (S2-2), 174,989.4 ± 26,055.4 μm3 (CDL 69), and 59,636.1 ±15,259.0 μm3 (S2BAC). Exposure to SHY at 0.5 MIC reduced maximum thickness of biofilms formed by S2-1 (p < 0.01; 24.15 ± 3.36 μm vs. 31.23 ± 2.07 μm), and reduced surface coverage (p < 0.05; 82.33 ± 6.38% vs. 98.00 ± 0.37%) and increased roughness (p < 0.001; 0.5768 ± 0.0400 vs. 0.2818 ±0.0125) of biofilms by S2BAC.
On the other hand, BZK at 0.5 MIC reduced (p < 0.05) the biovolume of biofilms formed by S2-1 (BZK-susceptible strain; 184,035.5 ± 21,115.0 μm3) and increased the biomass of biofilms formed by BZK-resistant strains (S2-2, CLD 69, and S2BAC), which showed a larger (p < 0.05) biovolume (274,521.6 ± 2,901.6 μm3, 238,496.3 ± 1,583.4 μm3, and 172,029.6 ± 1,348.6 μm3, respectively) and a higher (p < 0.05) maximum thickness (29.14 ± 1.70 μm, 29.20 ±0.35 μm, and 24.96 ± 1.95 μm, respectively) than did unexposed strains (23.76 ± 0.52 μm, 22.28 ± 2.71 μm, and 16.10 ± 0.24 μm for strains S2-2, CLD 69, and S2BAC, respectively).
The percentage of dead or damaged bacteria in biofilms (calculated as the biovolume of PI-stained cells relative to total biovolume) increased in the presence of sub-MICs of SHY. In the presence of 0.5 MIC of BZK, percentage of PI-stained cells increased in biofilms formed by S2-1 (BZK-susceptible strain). By contrast, in the case of BZK-resistant strains (S2-2, CLD 69, and S2BAC), percentage of dead or damaged cells did not differ significantly (p > 0.05) between biofilms growing in the absence of biocides and in the presence of sub-MICs of BZK. Percentage of PI-stained cells in control (unexposed) biofilms ranged from 0.21 ± 0.11% (S2-1) to 2.40 ± 0.91 (CLD 69). Values for biofilms growing in the presence of sub-MICs of SHY ranged from 1.48 ± 1.42% (S2-2) to 4.99 ± 0.49% (S2BAC). For biofilms growing in the presence of BZK at 0.5 MIC, values ranged from 0.52 ± 0.37% (S2-2) to 2.27 ± 0.79% (S2BAC).
Effect of disinfectants used at 1 or 1.5 MIC
When cultures were exposed to biocides at 1 or 1.5 MIC, biofilm formation was inhibited, and only a few small, scattered cell aggregates were detected in the observation field of 14,161 μm2, with biovolumes ranging from 6.4 ± 4.2 μm3 to 266.3 ± 77.2 μm3, and surface coverage ranging from 0.10 ± 0.07% to 0.62 ± 0.34%.
Considering all conditions (exposure to SHY and BZK at 0.5, 1, and 1.5 MIC) for all strains simultaneously, a positive correlation (from 0.799 to 0.918; p < 0.001) was found between biovolume, surface coverage, and maximum thickness of biofilms. All of these parameters were negatively correlated with biofilm roughness (from −0.835 to −0.603; p < 0.01).
The post-hoc analysis showed higher (p < 0.001) percentages of dead bacteria in the presence of 1 or 1.5 MIC of SHY or BZK (the figures ranged from 23.57 ± 4.14% to 93.68 ± 2.85% for SHY, and from 32.60 ± 21.36% to 57.40 ± 7.30% for BZK) than in the absence of biocides or in the presence of such compounds at 0.5 MIC.
Discussion
Effect of disinfectants used at sub-MICs
MIC values of SHY and BZK observed for L. monocytogenes planktonic cultures are similar to data obtained by other authors, which ranged from 512 to 2500 ppm for SHY (Lundén et al., 2003; Gao and Liu, 2014), and from 0.63 to 15 ppm for BZK (Lundén et al., 2003; Soumet et al., 2005). L. monocytogenes strains tested were classified as susceptible (S2-1) or resistant (S2-2, CDL 69, and S2BAC) to BZK (Martínez-Suárez et al., 2016).
All strains tested were able to produce biofilm on polystyrene. However, there was interstrain variability. In the absence of biocides, the BZK-susceptible isolate (S2-1) produced stronger biofilms than the BZK-resistant L. monocytogenes strains (S2-2, CDL 69, and S2BAC). On the other hand, exposure to sub-MICs of BZK increased biofilm development by BZK-resistant strains and reduced biofilm by S2-1. These results agree with previous observations, where biofilm-forming ability of L. monocytogenes was assessed using Crystal Violet assays (Ortiz et al., 2014), and suggest that for BZK-resistant strains, sub-MICs of such disinfectant may represent an advantage, compensating for the weak biofilm formation level that might be associated with resistance.
Our findings are also in agreement with quantitative observations performed in earlier studies using different bacterial species and antimicrobials, where biofilm formation was induced by conditions that are potentially toxic for cells, such as the presence of sub-MICs of disinfectants (Capita et al., 2014, 2017; Buzón-Durán et al., 2017). The enhancement of biofilm formation in the presence of low doses of antimicrobials may represent an adaptive response to subinhibitory stress, and is likely to be related to alterations in the cellular morphology or composition (e.g., chemical and physical cell-surface characteristics), the increased expression of specific genes (e.g., those involved in quorum sensing), or the increase in production of extracellular polymeric substances in the biofilm matrix (Bisbiroulas et al., 2011; Kumar and Ting, 2013; Capita et al., 2014; Buzón-Durán et al., 2017). In the case of S2BAC, with an MDR phenotype, an association between efflux pumps and biofilm production has been previously suggested (Ortiz et al., 2014).
Several regulatory elements could be at play with the observations of this study. For example, regulatory factors controlling biofilm formation in L. monocytogenes could be affected by both SHY and BZK at 0.5 MIC. However, the enhancement of biofilm formation was only observed for BZK and BZK-resistant strains. The three BZK-resistant strains in spite of being genetically different, could share a similar resistance mechanism based on efflux pump activity. Both bcrABC resistance cassette (in strain CDL 69) or the qacH gene of the Tn6188 transposon (in strain S2-2) are examples of genetic determinants of transporters belonging to the small multidrug resistance protein family (Müller et al., 2013). For S2BAC, the overexpression of endogenous efflux pumps may be related to its MDR phenotype (Ortiz et al., 2016). Thus, it is suggested that regulatory elements affecting efflux pump activity could affect biofilm formation of different BZK-resistant strains. However, the exact mechanism responsible for the BZK-enhanced biofilm-forming capacity of L. monocytogenes cultures remains to be characterized. Further studies to elucidate the various possible morphological, biochemical, or molecular changes in L. monocytogenes biofilms induced by sub-MICs of this disinfectant are warranted.
The BZK-induced biofilm formation demonstrated in this study, for BZK-resistant L. monocytogenes strains, suggests that the use of this compound at inappropriate doses in the food industry may increase the ability of L. monocytogenes to produce biofilms. This raises food safety concerns, because biofilm structures are thought to play a major role in the persistence of bacteria and the contamination of foodstuffs (Capita et al., 2017; Ripolles-Avila et al., 2018). These findings have practical implications, because the levels of BZK at which L. monocytogenes biofilm formation is enhanced can easily be found in food-handling premises under some circumstances, as previously stated (Capita et al., 2014). Thus, the data from this study suggest the importance of maintaining doses higher than MICs of disinfectants to achieve efficient prevention of biofilm formation by L. monocytogenes.
The low percentage of dead or damaged cells observed in biofilms grown in the absence of biocides or in the presence of 0.5 MIC of SHY or BZK indicates that the membranes of most cells had not been compromised in such circumstances, as previously observed for several bacterial groups, when biocides are used at low doses (Buzón-Durán et al., 2017; Capita et al., 2017).
Effect of disinfectants used at 1 or 1.5 MIC
Only a few cell aggregates were detected in wells exposed for 24 h to disinfectants at 1 or 1.5 MIC. Because a similar number of cells were also observed in the wells tested immediately after adhesion and rinsing (before incubation; data not shown), it is suggested that cell aggregates present in the wells incubated for 24 h in the presence of biocides at 1 MIC (350 ppm of free chlorine or 9–13 ppm of SHY) or 1.5 MIC (525 ppm of free chlorine or 13.5–19.5 ppm of BZK) are likely to have been cells that were inoculated and adhered to the bottom of the well, which remained after rinsing. Thus, the findings in this study show that SHY and BZK may be able to prevent the development of biofilm even when below the concentrations conventionally used. These are 800–2000 ppm of free chlorine for chlorine-based compounds, such as SHY (Norwood and Gilmour, 2000; Henriques and Fraqueza, 2017), and 1000–5000 ppm for quaternary ammonium compounds, such as BZK (Tamburro et al., 2015; Poimenidou et al., 2015; Henriques and Fraqueza, 2017).
Conclusions
This study suggests that, while SHY or BZK may contribute to the prevention of biofilm formation by L. monocytogenes at doses equal or higher than MIC, even when doses are below the concentrations recommended for use, sub-MICs of BZK can enhance biofilm development in BZK-resistant strains, in a different extent according to the genetic marker or resistance mechanism involved. This phenomenon is worthy of note because biofilm formation is thought to play a major role in the persistence of L. monocytogenes. Hence, the findings in this work raise concern over the inappropriate use of disinfectants and suggest the importance of maintaining, in the food industry, concentrations of biocides higher than MICs so as to combat foodborne infections by L. monocytogenes biofilms. It should be noted, however, that results in this research should be interpreted with caution because only four strains of L. monocytogenes of molecular serotype 1/2a were tested. Further studies should be performed to elucidate the mechanisms through which BZK-resistant L. monocytogenes cells can enhance their biofilm-forming ability in the presence of sub-MICs of BZK.
Footnotes
Acknowledgment
The authors wish to thank the Spanish Ministerio de Economía, Industria y Competitividad (INIA, RTA2014-00045-C03).
Disclosure Statement
No competing financial interests exist.