
Abstract
Objectives:
The occurrence of Listeria monocytogenes was studied by using cultural and serological methods in cattle housed in a particular gaushala (cattle shelter) and organized dairy farm.
Materials and Methods:
A total of 1201 samples from cattle comprising blood (n = 207), milk (n = 203), vaginal swabs (n = 210), and serum (n = 207) from an organized farm (n = 210) and blood (n = 100), milk (n = 74), vaginal swabs (n = 100), and serum (n = 100) from a gaushala (n = 100) were collected and analyzed for L. monocytogenes. All samples excluding serum were analyzed for isolation and identification of L. monocytogenes, while the serum samples were screened for seropositivity. The isolates were further subjected to assess their virulence potential (in vitro and in vivo), biofilm formation ability, and antibiotic susceptibility patterns.
Results:
Four L. monocytogenes strains were isolated from the cattle; three (0.48%) from the organized farm and one (0.36%) from the gaushala. On serological screening of cattle from the organized dairy farm, 16.42% were found to be positive for antibodies against listeriolysin O, while cattle from the gaushala revealed 36% seropositivity. Furthermore, on characterization of the isolates for their pathogenic potential and biofilm-forming ability, all were found to be pathogenic by both in vitro and in vivo assays and were weak to moderate biofilm formers. The minimum inhibition concentration (MIC) of recovered isolates revealed resistance for ampicillin by two L. monocytogenes isolates (MIC >256 μg/mL), whereas three L. monocytogenes isolates were intermediately resistant (MIC >4 μg/mL) and one resistant against amoxicillin (MIC >8 μg/mL). However, all four isolates were susceptible to gentamicin, cotrimoxazole, and erythromycin.
Conclusions:
Isolation of virulent and antibiotic-resistant strains of L. monocytogenes warrants the need for epidemiological surveillance, antimicrobial susceptibility, and implementation of control measures to combat the occurrence of L. monocytogenes infection in animals as well as humans.
Introduction
Listeriosis caused by pathogenic strains of Listeria monocytogenes and Listeria ivanovii, is an important foodborne zoonotic disease of public health significance (Beye et al., 2016; Camargo et al., 2016). Globally, listeriosis occurs in the sporadic or epidemic form (Barbuddhe et al., 2012) and infections in animals are predominantly of subclinical nature; however, severe forms have also been reported (OIE, 2014). Ruminants, particularly cattle, contribute to the amplification and dispersal of L. monocytogenes into the farm environment, which subsequently serves as a source of infection for humans (OIE, 2014; Castro and Lindstrom, 2016). Cattle can contaminate the environment through their fecal shedding and vaginal secretions, wherein the organisms can survive for prolonged periods under harsh conditions, such as low temperatures, high moisture, and high salt concentrations (Kumar et al., 2007; Linke et al., 2014), and thereby can increase the risk of within-herd or intraherd transmission (Hurtado et al., 2017).
In India, cattle are normally reared for milk production and are housed either at the individual farmer level or in organized farms. However, cattle that are not productive, old, or those exhibiting physical, reproductive, or mammary disorders and are economically unsustainable at the individual level or in organized farms are often housed in gaushalas, that is, cattle shelters (Yadav, 2007; Dahiya, 2014; Khanna, 2017; Kennedy et al., 2018). In fact, gaushalas have played a significant role in protecting the cattle wealth in India (Dahiya, 2014; Kennedy et al., 2018). However, it has been reported that infectious organisms predominate in the environment of gaushalas due to lack of appropriate hygienic conditions (Kumar, 2008). In addition, majority of animals housed in gaushalas have immunocompromised health status and studies highlighting the occurrence of listeriosis in cattle housed in gaushalas are lacking. Similarly, cattle housed in organized dairy farms are often high milk yielders and such animals are under production stress. The objective of the present study was to decipher the occurrence of L. monocytogenes in a cattle population housed in a particular gaushala and an organized dairy farm using cultural and serological methods and their further characterization using in vitro and in vivo pathogenicity assays, biofilm-forming capability, and antimicrobial susceptibility patterns.
Materials and Methods
Bacterial strains
The standard strains of L. monocytogenes Microbial Type Culture Collection (MTCC) 1143 (NCTC 11994) and Staphylococcus aureus MTCC 737 (ATCC 29213) were obtained from MTCC and Gene Bank, Institute of Microbial Technology, Chandigarh, India.
Synthetic peptide of listeriolysin O
The synthetic peptide of listeriolysin O (LLO) found specifically in the earlier study (Shoukat et al., 2013), designated as LLO-2 peptide (WDEINYDPEGNEIVQHKNWSENNKSKLA), was synthesized commercially (IgY Immunologix Pvt. Ltd., Hyderabad, India).
Farm presentation and collection of samples
The gaushala under investigation housed immunocompromised, weak, and aged animals having reproductive disorders. The gaushala had six animal sheds with one calving box and a large open paddock. Most of the animals were donated by farmers. Animals were fed daily twice with roughage, green fodder or concentrates, or sometimes with donated foods such as jaggery, vegetables, and wheat flour. The animals were always let loose in the open paddock. Animals had access to water ad lib. Disposal of dung and other solid wastes was within the farm premises. Natural service was preferred for breeding, and milking was not practiced regularly. Animals in advanced pregnancy stage were housed in separate sheds or in the calving box. Disinfection of the shed and utensils was rarely practiced. However, treatment of sick animals was performed under veterinary supervision.
In the organized farm, intensive rearing practice was followed. The animals housed at organized farms were stall fed with clear monitoring of feeding, breeding, and management practices. Artificial insemination and treatment of ailments were strictly practiced and recorded under veterinary supervision. A calving box was provided for the animals and quarantine and biosecurity measures were also adopted in the farm.
A total of 1201 samples comprising blood (n = 207), milk (n = 203), vaginal swabs (n = 210), and serum (n = 207) from cattle of the organized farm (n = 210) and blood (n = 100), milk (n = 74), vaginal swabs (n = 100), and serum (n = 100) from cattle of the gaushala (n = 100) were collected. Clinical status of the cattle screened and details of the samples collected from the organized farm and gaushala are presented in Table 1. From each animal, a vaginal swab, and wherever possible, blood, milk, and serum samples were collected aseptically in sterile vials and transported to the laboratory under chilled conditions and processed immediately.
Details of Samples Collected from the Gaushala (Cattle Shelter) and Organized Farm
Isolation of Listeria
Isolation of Listeria species from vaginal swabs, blood, and milk was attempted as per the U.S. Department of Agriculture (USDA) method described by McClain and Lee (1988) with suitable modifications. Briefly, samples were enriched by a two-step enrichment procedure using University of Vermont medium–I (UVM-I, Himedia, India), followed by Modified University of Vermont medium–II (MUVM-II). Each of the collected samples (swab or 1 mL of blood/5 mL milk) was aseptically transferred into UVM-1 in a 1:10 proportion and incubated at 30°C for 24 h. The enriched inoculum (0.1 mL) from UVM-1 was then transferred to MUVM-2 medium (ceftazidime pentahydrate, 37.6 mg/L) and incubated overnight at 30°C. A loopful of inoculum from enriched MUVM-2 was streaked on PALCAM agar plates (Himedia) and incubated at 37°C for 48 h. The greenish yellow, glistening, iridescent, and pointed colonies of about 0.5 mm diameter surrounded by a diffuse black zone of esculin hydrolysis were suspected to be Listeria.
Confirmation of isolates
The presumed colonies of Listeria (at least 5/plate) were morphologically and biochemically tested by Gram staining, catalase reaction, tumbling motility at 20°C–25°C, methyl red–Voges-Proskauer reactions, nitrate reduction, and fermentation of sugars (rhamnose, xylose, and α-methyl-D-mannopyranoside). Isolates found to be positive for L. monocytogenes were further confirmed by Matrix-assisted laser desorption and ionization – Time-of-flight – Mass Spectrometry (bioMérieux, Inc., France), as described earlier (Barbuddhe et al., 2008).
Phosphatidylinositol-specific phospholipase C activity on Agar Listeria Ottavani and Agosti medium
The biochemically characterized Listeria isolates were tested for their phosphatidylinositol-specific phospholipase C (PI-PLC) activity using Agar Listeria Ottavani and Agosti (ALOA) medium (Ottaviani et al., 1997). The pathogenic Listeria spp. exhibit halo formation, indicating production of PI-PLC.
Hemolytic titer assay
The hemolytic titer assay for test isolates was performed as described earlier (Young et al., 1986) and hemolytic units were expressed as the reciprocal of the highest dilution of toxin, which is required to lyse 50% of the erythrocytes.
Chick embryo inoculation test
The virulence potential of L. monocytogenes isolates was assessed by the chick embryo inoculation test (Notermans et al., 1991). In brief, the blood vessel-free surface of precandled, 10-day-old, embryonated chicken eggs was inoculated with 0.1 mL of brain heart infusion broth containing test culture (107 CFU/mL) through the chorioallantoic membrane route. For each isolate, two eggs were used. The inoculated eggs along with the controls (positive and negative) sealed with molten paraffin were incubated at 37°C for 7 days. Test isolates exhibiting mortality of embryo after 24 h up to 7 days postinfection were considered to be virulent.
Galleria mellonella larval infection
Infection of Galleria mellonella larvae was performed as described earlier (Mukherjee et al., 2010). The log-phase-grown test isolates of L. monocytogenes (106 CFU) were injected dorsolaterally into the hemocoel of the last instar of each larva using a Hamilton syringe (capacity 50 μL; 24 G; point style 2 with beveled tip). For each test isolate, three larvae were used. L. monocytogenes (MTCC 1143) was used as the reference control. Mortality in infected larvae was monitored at 12-h intervals for up to 7 days postinfection.
Biofilm-forming assay
The biofilm-forming ability of L. monocytogenes isolates was estimated (Borucki et al., 2003; Wang et al., 2017) with slight modifications. In brief, a single isolated colony of test isolate was inoculated into 5 mL of tryptone soya yeast broth and incubated at 37°C in a shaker incubator at 180 rpm for 24 h. An aliquot (200 μL) of the overnight grown culture was dispensed into 7 wells of a column of sterile polyvinylchloride 96-well microtiter plate (GenAxy). The eighth well of the column was kept as medium control. The inoculated culture plate was incubated at 37°C for 72 h for biofilm formation. Then, the plates were washed three times with sterile water (250 μL) and air-dried for 30 min, after which an aqueous 0.1% (w/v) crystal violet (CV) solution (200 μL) was added to each well and incubated for 45 min at room temperature. Unbound dye was removed by rinsing three times in 250 μL of sterile water. The CV-stained biofilm-forming bacteria were then solubilized by adding 210 μL of 95% ethanol for 30 min at 4°C. The contents of each well (200 μL) were then dispensed to another sterile polystyrene microtiter plate and the optical density (OD) at 595 nm of each well was measured using a microplate reader (Thermo Scientific Multiskan GO). The biofilm-forming ability was calculated by subtracting the OD of control wells from the average OD of the test isolate.
Antibiotic susceptibility test
The minimum inhibition concentration (MIC) of isolates was determined using HiComb™ MIC strips (Himedia, India). Staphylococcus aureus MTCC 737 was used as a quality control strain. In brief, the log-phase-grown test cultures (1.5 × 108 CFU/mL) were swabbed on Mueller-Hinton agar (Himedia) plates supplemented with 5% sheep blood. The plates were then placed with the respective HiComb antibiotic MIC strip and incubated at 37°C for 24 h. MIC values obtained for each test isolate against antibiotics (ampicillin, cotrimoxazole) were used to grade the test isolate as per CLSI (2017) guidelines, whereas for other antibiotics, CLSI criteria (M45-CLSI 2016) for Streptococcus were applied to grade the test isolates.
Indirect enzyme linked immunosorbent assay
The indirect enzyme linked immunosorbent assay (ELISA) (iELISA) was performed (Low et al., 1992; Shoukat et al., 2013) with suitable modifications. In brief, the assay was standardized by checkerboard titration using known positive (culturally and serologically positive) and negative (culturally and serologically negative) cattle serum. The serum sample at a dilution of 1:200 with a positive-to-negative ratio greater than or equal to 2.5 (OD492 nm) was considered as positive for listeriosis in a standardized iELISA by employing the synthetic peptide of LLO designated as LLO-2 (40 ng/well) as the antigen and rabbit anti-bovine HRP conjugate (1:2000; Santacruz). This standardized synthetic peptide-based iELISA was employed for screening antibodies against LLO (ALLO) from cattle serum samples collected from the gaushala and organized farm. The serum samples found to be positive initially were further adsorbed with streptolysin O (SLO) (Sigma-Aldrich) and rescreened as described earlier (Berche et al., 1990; Shoukat et al., 2013).
Statistical analysis
Biofilm formation of test isolates was analyzed using the one-way ANOVA test (Bonferroni's multiple comparison test) to observe the difference in biofilm formation within isolates at the 95% confidence interval level.
Results
Isolation of Listeria
Results of isolation of L. monocytogenes from samples collected from the gaushala and organized dairy farm are presented in Table 2. Microbiological screening of 620 samples from the organized farm and 274 samples from the gaushala (excluding serum samples) yielded four L. monocytogenes isolates. Three (0.48%) L. monocytogenes isolates were recovered from the organized farm, while one (0.36%) L. monocytogenes isolate was recovered from the gaushala. Of the three L. monocytogenes isolates from the organized farm, two were recovered from apparently healthy animals (BM242 and BB117), while one was isolated from an animal having a history of miscarriage (BM205). In the gaushala, the animal found to be positive for L. monocytogenes (RB26) had a history of miscarriage (Table 2).
Biochemical, Pathogenicity, and Serological Profiles of Listeria monocytogenes Isolates Recovered from Cattle
G, Gram's staining; C, Catalase test; M, tumbling motility (20–25°C); MR, methyl red test; VP, Voges-Proskauer test; NR, nitrate reduction; R, rhamnose fermentation; X, xylose fermentation; α, α-methyl-D-mannopyranoside fermentation; ALLO, against listeriolysin O; SLO, streptolysin O; ALOA, agar listeria ottavani and agosti; PI-PLC, phosphatidylinositol-specific phospholipase C.
Serological testing
On screening of serum samples from cattle (n = 207) from the organized dairy farm, 38 (18.35%) samples were found to be positive for ALLO. However, after adsorption with SLO, seropositivity was observed in 34 (16.42%) samples. Seropositivity for ALLO antibodies before and after SLO adsorption was observed in 41 (41%) and 36 (36%) cattle housed in the gaushala, respectively.
Of the four animals that were culturally positive, three revealed seropositivity for ALLO antibodies before and after SLO adsorption of serum (Table 2).
Pathogenicity testing
All the four isolates were found to be pathogenic by employing in vitro and in vivo assays. Of the four L. monocytogenes isolates, the isolate recovered (BM205) from the animal with reproductive disorder from the organized farm revealed higher hemolytic titer (256 hemolytic units), prominent PI-PLC activity on ALOA medium within 24 h, and was equally lethal to chick embryo and G. mellonella larvae (mortality within 48–72 h postinfection) compared with the other three L. monocytogenes isolates (Table 2).
Biofilm-forming ability
The biofilm-forming abilities of the test isolates were graded as described earlier (Harvey et al., 2007; Doijad et al., 2015). The isolate (BB117) recovered from the organized farm was found to be a moderate biofilm producer (OD595 = 0.34), while the remaining two (BM205, BM242) isolates recovered from the organized farm and one isolate (RB26) recovered from the gaushala were weak biofilm producers (OD595 < 0.323) (Fig. 1). Statistically significant differences (p > 0.05) were not observed between the isolates for biofilm formation; however, significant differences (p < 0.05) in biofilm formation were noted when compared with medium control.
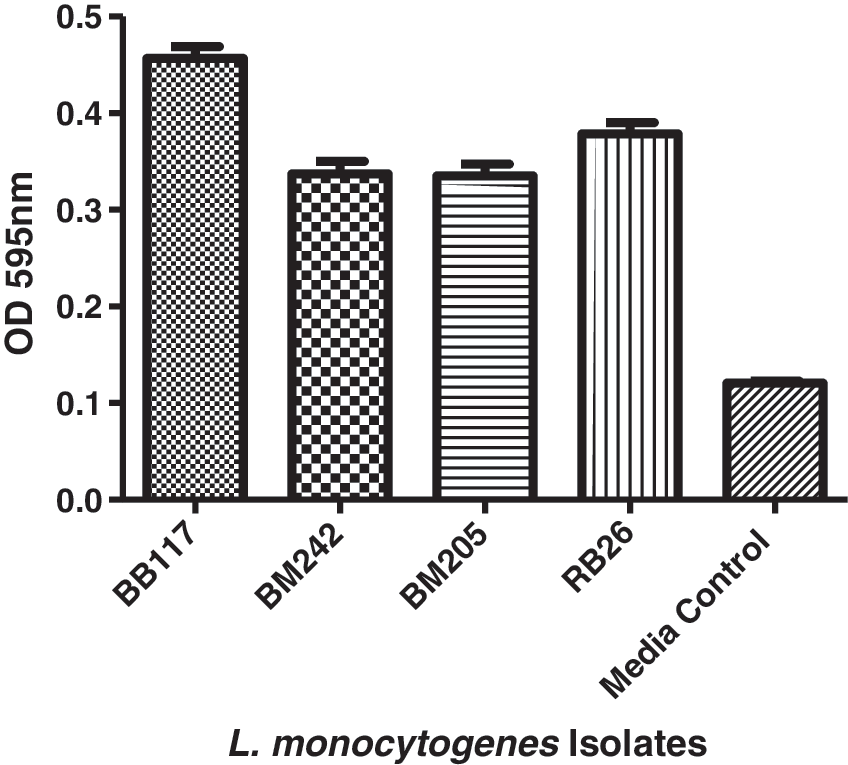
Biofilm formation of Listeria monocytogenes isolates: Mean OD 595 nm values of biofilm formation by L. monocytogenes isolates stained with crystal violet. The isolates were categorized as weak (OD value <0.32), moderate (OD value between 0.32 and 0.64), or strong (OD value >0.64) biofilm formers. OD, optical density.
Antibiotic susceptibility
Of the four L. monocytogenes isolates, two (BM205, BB117) were found to be sensitive to ampicillin, while the remaining two isolates (BM242, RB26) were resistant to ampicillin (MIC range >256 μg/mL). Three isolates were intermediately resistant (MIC >4 μg/mL) and one resistant against amoxicillin (MIC >8 μg/mL) (Table 3). However, all four isolates were susceptible to gentamicin, cotrimoxazole, and erythromycin (Table 3).
Minimum Inhibitory Concentrations of Antibiotics Tested Against Listeria monocytogenes Isolates
MIC, minimum inhibition concentration.
Discussion
In ruminants, several predisposing factors such as lowered immunity, production stress, and management practices, including improper housing, overcrowding, insanitary conditions, poor access to good quality pasture, and drinking water, favor the occurrence of listeriosis (Burgess and Lohmann, 2006; Amene and Firesbhat, 2016). Gaushalas and organized farms are managed differently and are quite contradictory to each other, therefore we studied the occurrence of L. monocytogenes in the gaushala and organized dairy farm.
In this study, owing to history of animal health status and management practices at the gaushala, we were expecting higher cultural and serological positivity for listeriosis; however, only one animal from the gaushala was found to be positive for L. monocytogenes, although we observed higher seropositivity (36%) for ALLO antibodies. This higher seropositivity suggested that the animals housed in the gaushala might have been exposed to L. monocytogenes infection earlier. Previous studies revealed the persistence of antibodies (ALLO) to L. monocytogenes infection for up to 126 days in orally infected calves (Barbuddhe et al., 2000) and these antibodies can even last for more than 2 years (Baetz and Wesley, 1995). Additionally, these antibodies had also shown speedy clearance of L. monocytogenes from the gastrointestinal tract of experimentally infected animals (Miettinen et al., 1990: Borku et al., 2006), which could be a probable reason for the low isolation rate at the gaushala.
In the organized dairy farm, of the three animals that were culturally positive, two were serologically positive. The absence of ALLO in the culturally positive animal might be a possible cross-contamination of samples originating from environmental sources (Kampelmacher and van Noorl Jansen, 1972). It has been well documented that L. monocytogenes exists and multiplies as a saprophytic organism in the soil and plants as well as in sewage and river water (Farber and Peterkin, 1991; Wegener et al., 1993). Such finding also underlines the importance of multiple tests for screening of listeriosis in animals. Of the three animals that tested positive, two were recovered from healthy animals and one from an animal having a history of miscarriage. Previous studies reported the presence of L. monocytogenes in healthy animals (in their gastrointestinal tract) and such animals quite often excrete the pathogen in their excretion, including feces, without any apparent clinical signs (Skovgaard and Morgen, 1988; Iida et al., 1991; Elezebeth et al., 2007).
We characterized the recovered isolates for their pathogenic potential because large number of Listeria isolates of various kinds (hemolytic and pathogenic, hemolytic and nonpathogenic, as well as nonhemolytic and nonpathogenic) and types (typical and atypical) exit in clinical cases as well as surrounding environmental sources (Shoukat et al., 2013). In this study, irrespective of their place of isolation, all the L. monocytogenes isolates were found to be variably pathogenic. Similar observations have also been reported earlier (Shakuntala et al., 2006; Negi et al., 2014; Shoukat et al., 2014).
Furthermore, we characterized the recovered isolates for biofilm formation as biofilm-forming bacteria had been reported to be highly resistant to desiccation, ultraviolet light, and antimicrobial agents (sanitizers and disinfectants) and could also persist in the environment for longer periods, leading to cross-contamination of food (Giaouris et al., 2014; Puga et al., 2016). In this study, three of the four L. monocytogenes isolates were weak biofilm formers and one was moderate. Significant differences (p > 0.05) in biofilm formation were not observed between the isolates. Previous studies have also reported that listerial cells are generally weak to moderate biofilm producers (Borucki et al., 2003; Harvey et al., 2007; Doijad et al., 2015).
In an earlier study, several strains of L. monocytogenes were found to be resistant to one or more antibiotics, which are commonly used for the treatment of listeriosis (Yan et al., 2010). In addition, multidrug-resistant L. monocytogenes isolates have been reported from food, animals, and humans (Shen et al., 2013). In this study, irrespective of the place of isolation, two isolates were found to be resistant to ampicillin and three isolates were intermediately resistant to amoxicillin, whereas all the isolates were found to be susceptible to gentamicin, cotrimoxazole, and erythromycin. These findings are of concern for animals as well as public health as ampicillin, either alone or in combination with an aminoglycoside (gentamicin), is the recommended choice of treatment for listeriosis in immunocompromised patients.
Conclusions
The present study revealed low cultural positivity for L. monocytogenes among cattle housed in the gaushala and organized farm; however, high seropositivity was observed among cattle from the gaushala when compared with the organized farm. In the absence of appropriate control measures, the presence of virulent and antibiotic resistance strains of L. monocytogenes in the raw milk of farm animals may pose potential risk to consumers and farm workers. Thus, it warrants further epidemiological investigations of predisposing factors and antimicrobial resistance patterns along with strict regulatory measures to minimize the incidence of listeriosis in animals as well as humans.
Footnotes
Acknowledgments
The authors thank the Director, ICAR-Indian Veterinary Research Institute, Izatnagar, India, for providing necessary facilities for the research. The technical assistance offered by Mr. K.K. Bhat and Dr. Deepa Ujjawal is duly acknowledged.
Funding
This work was supported by grants from the Department of Biotechnology, Government of India, under Grant [BT/01/CEIB/11/VI/13] to S.V.S.M and a Junior Research Fellowship to S.R. by University Grants Commission.
Disclosure Statement
No competing financial interests exist.