
Abstract
This study is aimed at detecting and characterizing methicillin-resistant Staphylococcus aureus (MRSA) from bulk tank milk samples of cows, sheep, and goats collected from dairy farms in the Czech Republic. All MRSA isolates were identified using PCR detection of the Staphylococcus aureus-specific fragment SA442 and mecA gene. The staphylococcal chromosomal cassettes mec (SCCmec), spa, and multilocus sequence types (MLST) were determined. The presence of genes encoding enterotoxins (ses), Panton–Valentine leukocidin (pvl), exfoliative toxins A, B (eta, etb), and toxic shock syndrome toxin (tst) were assessed. To differentiate human and animal origin, the presence of staphylokinase (sak) gene, ϕSa3 prophage, and susceptibility to tetracycline was tested. Out of 49 bulk tank milk samples examined, 14 (28.6%) were MRSA-positive. Eleven positive samples came from cow's milk (38%) and the remaining three from goat's milk (33%). All samples of ewe's milk were negative. In MRSA isolates three sequence types containing seven spa types were identified. Twelve isolates (85.7%) belonged to ST398 spa types t011/SCCmec IVa, t011/SCCmec V, t034/SCCmec V, t1456/SCCmec IVa, t1255/SCCmec V, and t2346/SCCmec V. Another two isolates belonged to ST5/t3598/SCCmec IVa and ST8/t064/SCCmec IVNT. In six isolates, one or more ses genes (seb, sed, seg, sei, and sej) were confirmed. One isolate from cow's milk harbored the tst gene. Another two isolates (ST398/t1456/SCCmec IVa and ST5/t3598/SCCmec IVa) harbored the sak gene and ϕSa3 prophage, and the latter was the only tetracycline-susceptible isolate in this study. However, none of the isolates was positive for pvl or eta, etb. These results suggest that there is the wide geographical spread of ST398 across different regions of the Czech Republic with no host preference among dairy cattle and goats. Therefore, when evaluating the occupational and foodborne risks, MRSA carriage and infection should be taken into account.
Introduction
Mastitis is an infectious disease of the mammary gland that is responsible for significant financial losses to dairy farmers worldwide, mainly due to reduced milk production and discarded milk (Aires-de-Sousa et al., 2007). Although several bacterial pathogens can cause mastitis, Staphylococcus aureus is one of the most important etiological agents for both clinical and subclinical mastitis, which is the primary reason for therapeutic use of antibiotics in dairy animals (Vanderhaeghen et al., 2010).
The emergence of methicillin-resistant Staphylococcus aureus (MRSA) on dairy farms can be responsible for the colonization or infection of farm personnel. It is also a source of contamination for consumers via the food chain (Papadopoulos et al., 2018). Livestock-associated MRSA (LA-MRSA) has created public health concerns because it is a source of antimicrobial-resistant bacteria, which can be transmitted to humans (Asiimwe et al., 2017). It is known that people working in close contact with animals, particularly on MRSA-positive farms, are at a higher risk of being colonized with LA-MRSA (Schmidt et al., 2017).
In addition to the established risk of direct contact with animals, the presence of LA-MRSA in the human food chain is an additional potential route for the spread of the strain to the human population (Smith and Wardyn, 2015).
LA-MRSA has unique molecular characteristics that distinguish them from human MRSA [hospital-associated (HA) or community-associated (CA)]. Studies have defined LA-MRSA based on the clonal complexes (CCs) and antimicrobial resistance patterns. CC398 has been referred to as the most pandemic LA-MRSA in Europe (Köck et al., 2013). It represents a distinct subgroup of MRSA mostly characterized by sequence type (ST) 398. The staphylococcal chromosomal cassettes mec (SCCmec) found in LA-MRSA are limited mainly to SCCmec IVa and SCCmec V (Butaye et al., 2016). Analyses of the virulence properties have shown a lack of the major virulence factors such as Panton–Valentine leukocidin (pvl), toxic shock syndrome toxin (tst), and exfoliative toxins. However, a small number of ST398 isolates carry the enterotoxin genes (Cuny et al., 2013). The human-specific immune evasion cluster (IEC), such as staphylokinase (sak) and ϕSa3 prophage, is currently considered to be the marker for human host adaptation of MRSA (Rinsky et al., 2013).
In the Czech Republic, the prevalence of LA-MRSA from food-producing animals was low, and MRSA strains predominantly belonged to the ST398 (Bardoň et al., 2012). In recent years, the diversity and detection rate of MRSA in farm animals is on the rise (Tegegne et al., 2017). Although LA-MRSA is a growing problem in the country, its prevalence on dairy farms has not been assessed satisfactorily. Therefore, this study was designed to assess the occurrence and molecular characteristics of MRSA from bulk tank milk samples of cows, sheep, and goats on Czech dairy farms.
Materials and Methods
Isolate collection
Forty-nine bulk tank milk samples of cows, sheep, and goats were collected from dairy farms between 2005 and 2017. The farms were located in 27 districts of 10 regions in the Czech Republic. In total, 49 dairy farms were included: 29 having cows, 9 having sheep, and 11 having goats. From each farm, a single bulk tank milk sample was collected. For the detection of MRSA, 25 mL of bulk milk was added into 225 mL of Mueller-Hinton broth (Oxoid, UK) containing 6.5% NaCl (Penta, CZ) and incubated at 37°C overnight. Subsequently, 1 mL of enriched broth was transferred into 10 mL of tryptone soya broth containing cefoxitin (3.5 mg/L) and aztreonam (10 mg/L) (Labmediaservis, CZ). After incubation (37°C/16–20 h) a loopfull of the enriched selective broth was streaked onto Brilliance™ MRSA 2 (Oxoid). Presumptive MRSA colonies were confirmed by PCR for the S. aureus-specific fragment SA442 (Martineau et al., 1998) and the mecA gene encoding resistance to methicillin (Oliveira and de Lencastre, 2002). Simultaneously, the resistance to cefoxitin in S. aureus-positive strains was tested by disk diffusion method (cefoxitin 30 μg; Oxoid) to avoid exclusion of non-mecA MRSA isolates (CLSI, 2017). MRSA-confirmed isolates were stored at −80°C for further analysis.
SCCmec, spa, and multilocus sequence typing
SCCmec typing was carried out by differentiating the mec from ccr gene complex as described previously (Milheiriço et al., 2007a). Subtyping of the SCCmec type IV was performed by applying the method described by Milheiriço et al. (2007b). Typing of the S. aureus protein A locus (spa typing) was performed according to the methodology by Friedrich et al. (2008). Primers were used according to Stegger et al. (2012). Specific spa sequences were analyzed through website (
Multilocus sequence typing (MLST) was performed based on seven housekeeping genes according to Enright et al. (2000). The STs were assigned according to the MLST website (
Minimum spanning tree (MST) of MRSA isolates was calculated using a modified version of Kruskal's algorithm incorporated in Ridom SeqSphere+ version 5.0.0 software (Ridom).
Detection of virulence factor encoding genes
Detection of clinically important staphylococcal enterotoxins (sea, seb, sec, sed, see, seg, seh, sei, and sej) and virulence factor encoding genes [pvl, exfoliative toxins A, B (eta, etb), tst, and sak gene] was performed using PCR according to Monday and Bohach (1999), Hososaka et al. (2007), and Sung et al. (2008), respectively. All MRSA isolates were analyzed for the acquisition of the ϕSa3 prophage—marker of human host adaptation (identified by Sa3int integrase gene)—according to Goerke et al. (2009).
Resistance to tetracycline
Resistance to tetracycline (30 μg) as a marker of LA-MRSA was conducted by disk diffusion test following the guidelines of the Clinical and Laboratory Standards Institute (CLSI, 2017).
Results
All results are summarized in Table 1. A total of 49 bulk tank milk samples were examined, of which 14 were MRSA-positive. Eleven isolates originated from cow's milk (38%) and three from goat's milk (33%). None of the samples from ewe's milk was positive for MRSA. All isolates were phenotypically resistant to cefoxitin and harbored the mecA gene; mecA-negative MRSA were not observed. Among the MRSA isolates typed, three different STs containing seven different spa types were confirmed. Most of the spa types (12/14) were associated with ST398 (t011, t034, t1255, t1456, and t2346). Two of the isolates were non-livestock-associated ST, such as ST8/t064 (goat's milk) and ST5/t3598 (cow's milk). Only three types of staphylococcal chromosomal cassettes were confirmed. In 10 isolates (71.4%) SCCmec type V was detected; the other three isolates harbored SCCmec type IVa (21.4%), and one isolate harbored SCCmec type IVNT (7.1%). All isolates were resistant to tetracycline except one of the non-livestock-associated strains (ST5/t3598) isolated from cow's milk (Table 1).
Characteristics of Methicillin-Resistant Staphylococcus aureu s Isolated from Bulk Tank Milk Samples
CA, community-associated; LA, livestock-associated; MLST, multilocus sequence typing; MRSA, methicillin-resistant Staphylococcus aureus; NT, non-typeable; R, resistant; S, sensitive; sak, staphylokinase gene; SCCmec, staphylococcal chromosomal cassettes mec; ses, genes encoding enterotoxins; TE, tetracycline; tst, toxic shock syndrome toxin gene.
Six isolates carried one or more genes encoding enterotoxins (seb, sed, seg, sei, and sej), but only two isolates harbored the classical enterotoxin genes (goat's milk—seb; cow's milk—sed).
In contrast, genes encoding virulence factors lukF-PV/lukS-PV, eta, and etb were not detected in any of the isolates. One isolate from cow's milk carried the tst gene, and another two isolates from cow's milk (ST5/t3598/and ST398/t1456) harbored a gene encoding sak. Those isolates that carried the sak gene were also positive for ϕSa3 prophage.
The isolates originating from cow's milk were more diverse in terms of spa types. The isolates from goat's milk were dissimilar (ST398/t011, ST398/t2346, and ST8/t064). The close relatedness of strains is indicated in the MST below (Fig. 1).
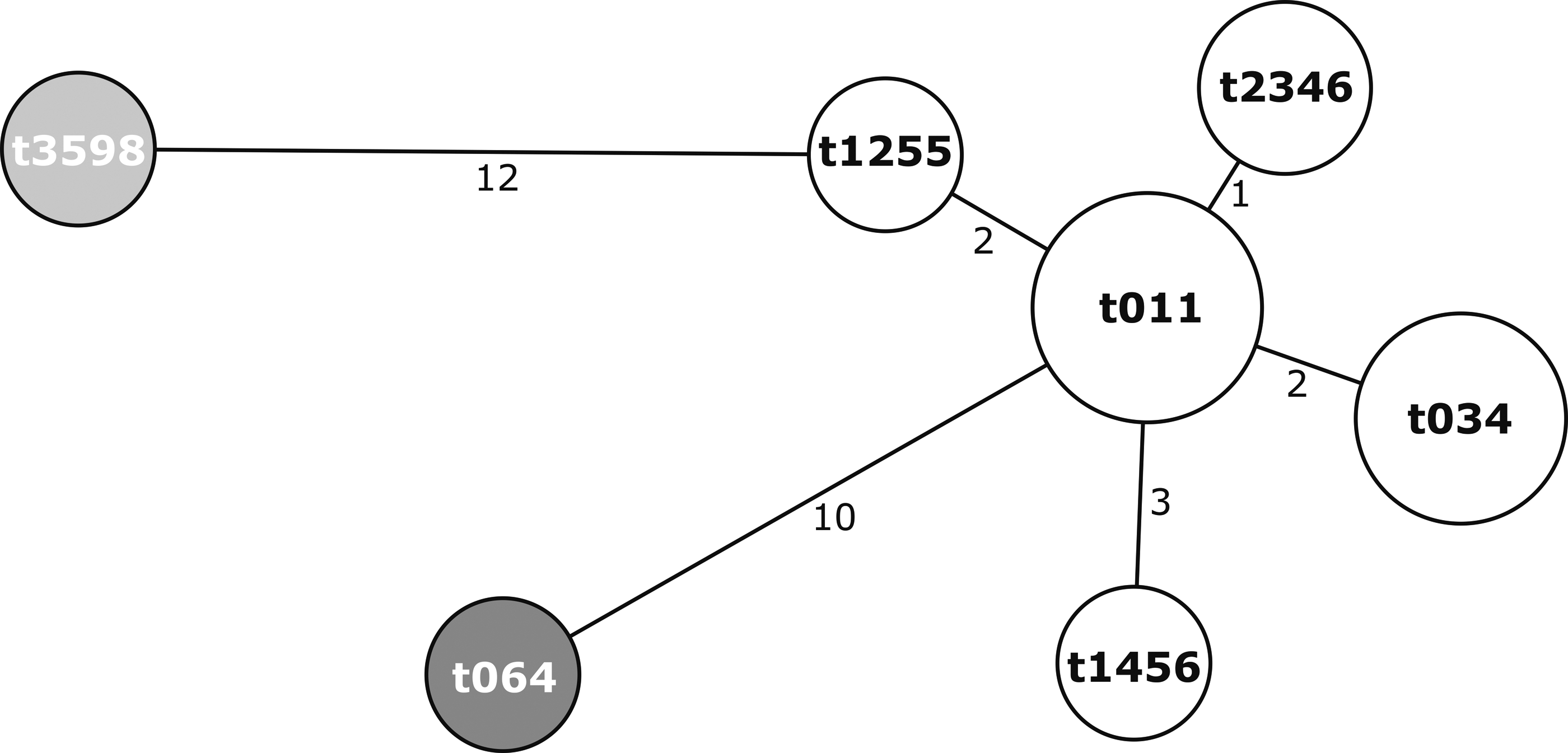
Minimum spanning tree of 14 methicillin-resistant Staphylococcus aureus isolates. Nodes indicate spa types and their size shows the relative number of isolates for each spa type. Every color represents a distinct multilocus sequence type (white—ST398; dark grey—ST8; light grey—ST5).
Geographically, MRSA is distributed widely across dairy farms throughout the country. Many of the recovered isolates from cow's milk (8/12) were spa types t011 and t034. Out of 77 districts in 14 regions, farms from 27 districts in 10 regions were included in the sampling of this study. Dairy farms from nine districts, each from a different region, were positive for MRSA (Fig. 2).
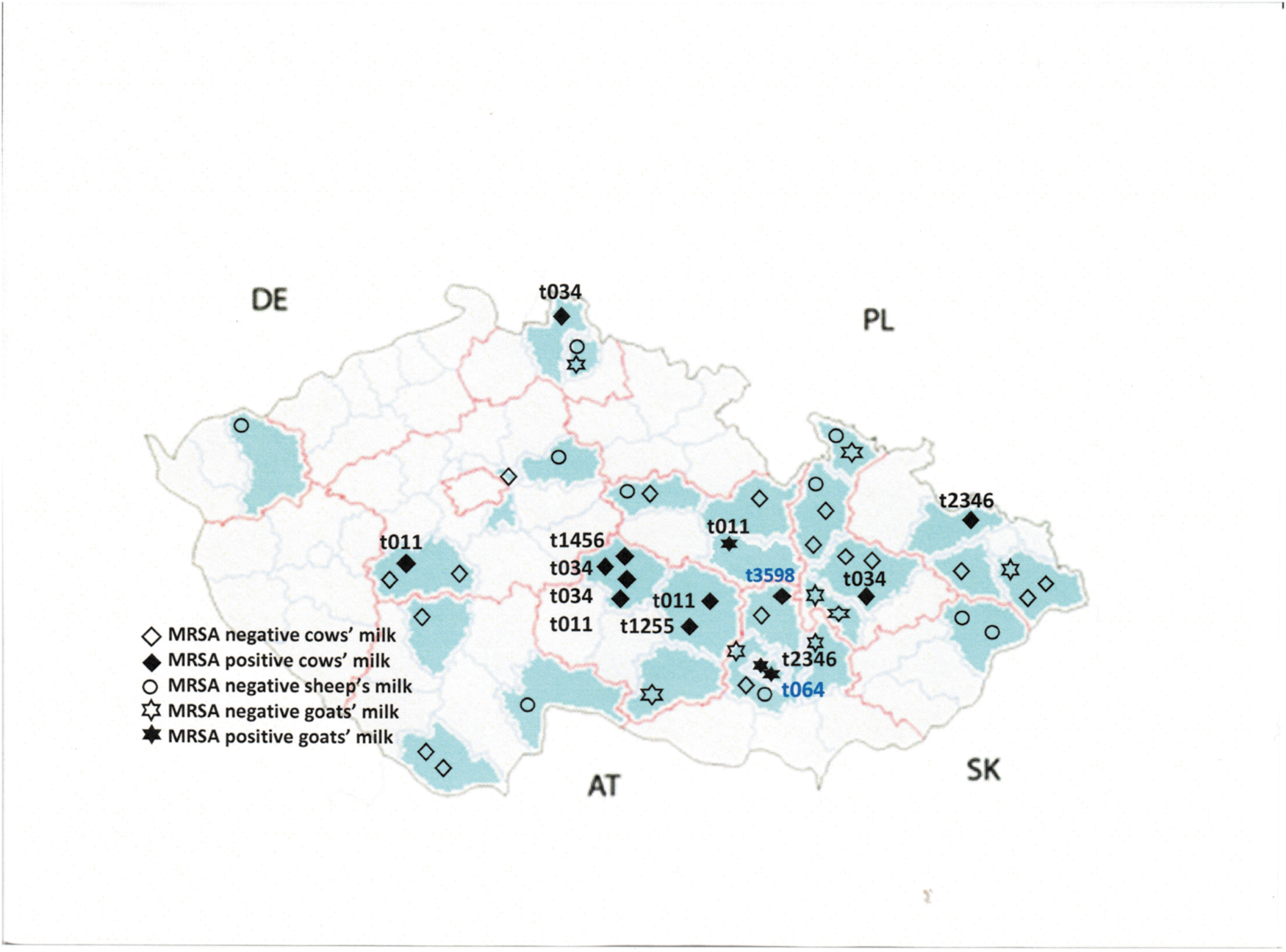
Map of the Czech Republic showing the locations of different regions (red line) and districts (blue line). Locations sampled, positive and negative findings of MRSA, and spa types are indicated. AT, Austria; DE, Germany; MRSA, methicillin-resistant Staphylococcus aureus; PL, Poland; SK, Slovakia.
Discussion
Based on the results obtained, all isolates belonged to sequence type ST398, except two of them: ST8/t064 (goat's milk) and ST5/t3598 (cow's milk). PVL-negative ST398 is considered to be an important livestock-associated lineage, mainly related to pig farming, but also found in cattle, sheep, and goats (Monecke et al., 2011). Because ST398 is able to overcome the species barrier, it has zoonotic significance. Previous studies have shown that nasal colonization or infection by ST398 was found in 23–86% of farmers and veterinarians (Cuny et al., 2015). The high colonization rate of ST398 MRSA in farmers and its emergence as a cause of clinical infections in humans indicate that MRSA ST398 in dairy animals may pose a public health risk (Köck et al., 2013).
In terms of ST398 diversity, our results differ from the report from the Netherlands where only spa types t011, t108, and t889 were detected out of 38,000 bulk milk samples (Tavakol et al., 2012). In our study, a wider variability in spa types of ST398 was detected (t011, t034, t1255, t1456, and t2346), and non-ST398 ST were also found (Table 1). LA-MRSA began as a single CC (CC 398), most of which was ST398, with few spa types involved (Feßler et al., 2011). This observation implies that the LA-MRSA population is still evolving quite rapidly (Peeters et al., 2015). The variability of spa types in European countries was described in the European Food Safety Authority surveillance report, and the spa types t011, t034, and t1255 are widely distributed in most European countries. However, spa type t2346 is particularly prevalent in Central European countries, such as Germany, Czech Republic, and Austria (EFSA, 2009). In different countries, specific spa types are dominant in most of the isolates, possibly indicating that certain clones only circulate within a given locality (Butaye et al., 2016).
All LA-MRSA ST398 isolated in our study were resistant to tetracycline. Tetracycline is one of the most common antibiotic classes administered to food animals (Price et al., 2012). Tetracycline resistance is common among CC398 LA-MRSA strains and is absent in human-associated strains. The overlap between phenotypic (resistance to tetracycline) and molecular (sak-negative) markers of LA-MRSA was observed among the isolates of ST398. A single livestock-associated isolate showed resistance to tetracycline and was sak-positive. It remains unclear whether the acquisition of specific tetracycline resistance genes (e.g., tetK) plays a role in the co-selection of methicillin resistance, given that virtually all LA-MRSA ST398 isolates are already resistant to tetracycline due to the presence of another tetracycline resistance gene, tet(M) (Rinsky et al., 2013).
In addition to the LA-MRSA CC398/ST398 isolates, CC5/ST5 that corresponded to types commonly found in humans were also detected in this study. CC5/ST5 is a common and widespread CC that comprises both HA- and CA-MRSA (Fitzgerald, 2012). This lineage can also be considered an animal-adapted clone since it was reported in humans as well as in animals (Pantosti, 2012). The studies of MRSA in animals have shown that the majority of isolates belong to CC5/ST5 and to CC/ST398 lineages (Geenen et al., 2013; Krupa et al., 2014). ST5/t3598 represented our CC5/ST5 lineage. This spa type is rare, but it has already been reported in the Czech Republic (Tegegne et al., 2017) and Sweden (methicillin-sensitive S. aureus) (
Another ST detected in this study was ST8/t064 in goat's milk. Pvl-positive ST8 MRSA has been recognized as the leading cause of community- as well as health care-associated infections in North American hospitals (O'Hara et al., 2012). This lineage is increasingly being reported in hospitals outside of North America (El-Mahdy et al., 2014) with reports from Kuwait and many other countries in the Middle East (Boswihi et al., 2016). The loss of human-specific genes (pvl, sak, and ϕSa3 prophage) in our isolate may be a result of adaptation to non-human hosts. The spread from humans to animals appears to have been followed by its acquisition of animal-specific genes and partial loss of human-specific genes (Rinsky et al., 2013). Detection of ST8/t064 MRSA in this study highlights its growing importance as an international epidemic MRSA clone.
Conclusions
These results gave evidence that the diversity of MRSA lineages in dairy production is evolving. A wide geographical spread of MRSA, predominantly ST398, across dairy farms in the Czech Republic was observed. ST398 has no host preference in dairy cattle and goats. The presence of ST398 in milk samples indicates the potential introduction of the LA-MRSA lineage into the food chain. Detection of other ST such as ST5 and ST8 suggests a potential for contamination by personnel during processing. In general, the presence of MRSA in raw milk poses a potential risk of colonization and/or infection of humans, via both contact with dairy animals and raw milk consumption. Therefore, when evaluating occupational and foodborne risks, MRSA carriage and infection have to be taken into account.
Footnotes
Acknowledgments
The study was supported by projects of the Ministry of Agriculture of the Czech Republic (CR), NAZV KUS QJ1210284, QJ1510216, and project LO1218 from MEYS of the CR under the NPU I program.
Disclosure Statement
The authors confirm that this article content has no conflicts of interest.