
Abstract
A total of 189 samples of fresh products (leafy salads, ready-to-eat mixed salads, and fresh herbs) bought in retail in Southwest Germany were investigated for their microbiological quality and the presence of pathogenic bacteria, including Salmonella spp., Listeria monocytogenes, and presumptive Bacillus cereus. Total aerobic mesophilic plate counts (TAC) ranged from 5.5 to 9.6 log colony-forming units (CFUs) per gram. Enterobacteria and pseudomonads were the predominant microorganisms and were detected in all samples with counts between 5.0 and 9.2 log CFU/g. Strains of Escherichia coli were detected in 9 salad (7.9%) and 25 herb samples (33.3%). Significant differences in bacterial counts were found between conventionally and organically-grown products: in herbs the counts of moulds were significantly higher in organically-grown products, while E. coli was only detected in conventionally-grown products. In conventionally-grown salad samples, yeast counts were significantly higher. Salmonella Enteritidis was only detected in two conventionally- and in one organically-produced salad samples (2.6%). No coagulase-positive staphylococci were detected in fresh salads as well as in herbs. High levels of B. cereus sensu lato (≥3 log CFU/g) were detected in 19 vegetable salads (16.7%) and even in 55 samples of fresh herbs (73.3%). Listeria monocytogenes could not be detected in fresh herbs; however, three L. monocytogenes strains were isolated from two conventionally-produced salad samples and belonged to PCR serogroup IIa. Although our results indicate a high microbial load in fresh salads and herbs in Southwest Germany in 2015, the incidences of human pathogenic bacteria, that is, L. monocytogenes, Salmonella spp., and coagulase-positive staphylococci strains, were low.
Introduction
M
Up to now, the implication of plant-based foods in food-borne infections seems to play a minor part in statistics. However, the increasing popularity of raw vegetables and germinated seeds (sprouts) as “fresh” convenience foods and their distribution at retail constitute a special health risk and a potential for food-borne infections. These products are typically associated with high microbial populations. Typical representatives are mainly non-pathogenic bacteria that can be commonly found in the environment (Ragaert et al., 2007; Cardamone et al., 2015).
Nevertheless, the presence of pathogens and opportunistic pathogens cannot be excluded (Al-Kharousi et al., 2016). Fresh vegetable products can be contaminated with microorganisms at different stages of the food chain. Human pathogenic bacteria such as Salmonella spp., Listeria monocytogenes, toxin-producing Escherichia coli, and Bacillus cereus constitute a health risk for consumers and have been isolated from this product group repeatedly (Berger et al., 2010; Jung et al., 2014; Fiedler et al., 2017).
Until today, at least two outbreaks of L. monocytogenes and Salmonella Agona associated with vegetables are still ongoing within the EU (EFSA, 2018a, b). Vegetables were identified as one of the most common food vehicles associated with B. cereus-induced food-borne outbreaks in France (Glasset et al., 2016). In 2014, 16-weak-evidence outbreaks of staphylococcal toxins associated with vegetables and juices have been reported within the EU (EFSA and ECDC, 2015). The aim of our study was to investigate the microbiological quality of different fresh commercial produces obtained at retail in Southwest Germany and to record the presence of the above-mentioned human pathogenic bacteria. In addition, differences in the microbiological contamination of conventionally- and organically-produced products were compared.
Materials and Methods
Sample material
Sample material was bought at retail in Southwest Germany (Baden-Württemberg) from November 2014 to July 2015, mainly in the region of Karlsruhe. A total of 189 fresh products, including 114 leafy salads and 75 fresh herbs, were examined. Samples were tested immediately after purchase.
The investigated 114 leafy salad products could be subdivided into products that needed prior preparation steps such as washing and chopping before consumption (n = 40), while the remaining were labeled as ready for consumption (ready-to-eat, RTE; n = 74). Some RTE salads also included other vegetable ingredients (carrots, beetroots, white cabbage, chard, or spinach) and contained separately packaged products (e.g., croutons, ham, cooked chicken, or cheese). Only the vegetable parts of the RTE salads were investigated. Moreover, 79 of 114 leafy salad products were conventionally-produced, while the remaining 35 products were organically-produced.
The 75 herb samples comprised potted herbs, bulk goods, as well as foil-packed herbs. Comparable to the salads, herbs that were conventionally- (n = 50) as well as organically-farmed (n = 25) were investigated. All components of the packed herbs, including leaves and stalks, were used for microbial examination. In case of potted herbs, care was taken that neither soil nor roots were included in the study.
Determination of microbiological quality
Total aerobic mesophilic plate count (TAC), combined counts of enterobacteria and pseudomonads, and counts of yeast and moulds were analyzed. Twenty-five grams of each sample were mixed with 225 mL buffered peptone water (Biokar Diagnostics, Pantin, France) and were homogenized in a stomacher (AES Chemunex GmbH, Germany) for 2 min (10−1 dilution), followed by serial dilutions until 10−7 in one-fourth strength Ringer's solution (Merck KGaA, Germany). A volume of 100 μL of each dilution was spread in duplicate on selected media (Supplementary Table S1) using a Drigalski spatula.
Detection and characterization of human pathogenic bacteria
All produces were investigated for the presence of Listeria spp., Salmonella spp., presumptive B. cereus, and Staphylococcus aureus according to DIN ISO 11290-1 and -2, DIN ISO 6579, DIN ISO 7932, and DIN ISO 6888-1, respectively, using selective media for plate count (Supplementary Table S1) and enrichment of Listeria spp. and Salmonella spp.
Suspicious isolates were picked from selective medium, transferred to standard I broth (ST-I), and incubated at 37°C. Cultures were streaked out onto different selective media (Supplementary Table S1) to obtain pure cultures, which were identified by the application of biochemical or molecular methods.
For the identification of E. coli and suspicious Salmonella isolates, API ID 32 E test strips from bioMérieux (Nürtingen, Germany) were used according to the manufacturer's instructions. All suspicious Salmonella isolates were screened using different selective media (Supplementary Table S1). Xylose lysine desoxycholate and brilliant-green phenol-red lactose sucrose media were selected in accordance with section 64 of the German Food and Feed Act (LFGB, 2013), together with RAPID'Salmonella agar (Bio-Rad Laboratories GmbH, Munich, Germany), IBISA® (bioMérieux), and Rambach agar (Merck KGaA). All isolates were preserved at −80°C by the addition of 15% glycerol.
After positive confirmation of Salmonella strains by API ID 32 E, molecular confirmation was performed by DNA isolation using the Blood and Tissue Kit (Qiagen, Hilden, Germany) and by PCR with primers specific for the invA gene according to Kobayashi et al. (2009). Isolates that were identified as belonging to the genus Salmonella were sent to the National Reference Laboratory of Salmonella at the German Federal Institute for Risk Assessment (BfR) for subsequent serological differentiation.
All coagulase-positive staphylococci and L. monocytogenes were confirmed using API Staph and API Listeria (bioMérieux), respectively. For molecular typing, DNA was isolated according to Pitcher et al. (1989). Molecular serotyping of L. monocytogenes was done using multiplex PCR, targeting the fragments lmo1118, lmo0737, orf2110, orf2819, and prs (Doumith et al., 2004). Random amplifcation of polymorphic DNA (RAPD) analyses with two different primers HLWL 85 (Wernars et al., 1996) and UBC 155 (Farber and Addison, 1994) were used for typing. Clustering was performed with the Pearson correlation coefficient and unweighted pair group method with arithmetic mean using BioNumerics (bioMérieux) (Sneath and Sokal, 1973).
Data analysis of bacterial counts
The number of colonies is expressed in log10 colony-forming units (CFU) throughout the article. Statistical analyses were carried out using JMP® (version 13.2.1; SAS Institute GmbH, Heidelberg, Germany). Presumptive B. cereus values were calculated based on cell counts on MYP agar except for 5 of 75 herb samples for which counts on Bacara agar were used. Missing values were replaced by group means. For descriptive analysis, mean CFU values of each microorganism and product group were calculated from the raw data and log-transformed. Standard deviation was calculated from the log-transformed single values. Minimum and maximum values were identified.
Logarithmized data were prepared for further statistical analysis as follows: 0 value and values below the detection limit were set to 0.01, while missing values were replaced by group means. Significant differences in cell counts were identified using a one-way analysis of variance (ANOVA). Normal distribution of residuals was checked for all pairs with p-values <0.05 in one-way ANOVA. Pairs showing non-normal distributed residuals or heterogeneity of variance were analyzed for significant differences using Wilcoxon/Kruskal–Wallis and Steel–Dwass tests. In contrast, pairs showing normal distribution of residuals and homogeneity of variance were analyzed by Tukey–HSD test. Logarithmized and processed data were used for descriptive purposes and visualization by boxplots.
Results
Microbiological quality of fresh herbs
The TAC of 75 herb samples ranged from 5.5 to 9.6 log CFU/g (Fig. 1; Supplementary Tables S2 and S3). The counts of enterobacteria and pseudomonads ranged between 5.0 and 9.2 log CFU/g. Yeast and mould counts were detected from below detection limit to 6.3 and 5.7 log CFU/g, respectively. Counts of E. coli ranged between below detection limit and 4.2 log CFU/g. All staphylococci were confirmed as coagulase-negative and could be detected in all samples, with the exception of rosemary, with bacterial counts up to 5.2 log CFU/g. Salmonella spp., S. aureus, or L. monocytogenes were not detected. Presumptive B. cereus could be detected in 71 samples (94.7%) in counts up to 7.1 log CFU/g.
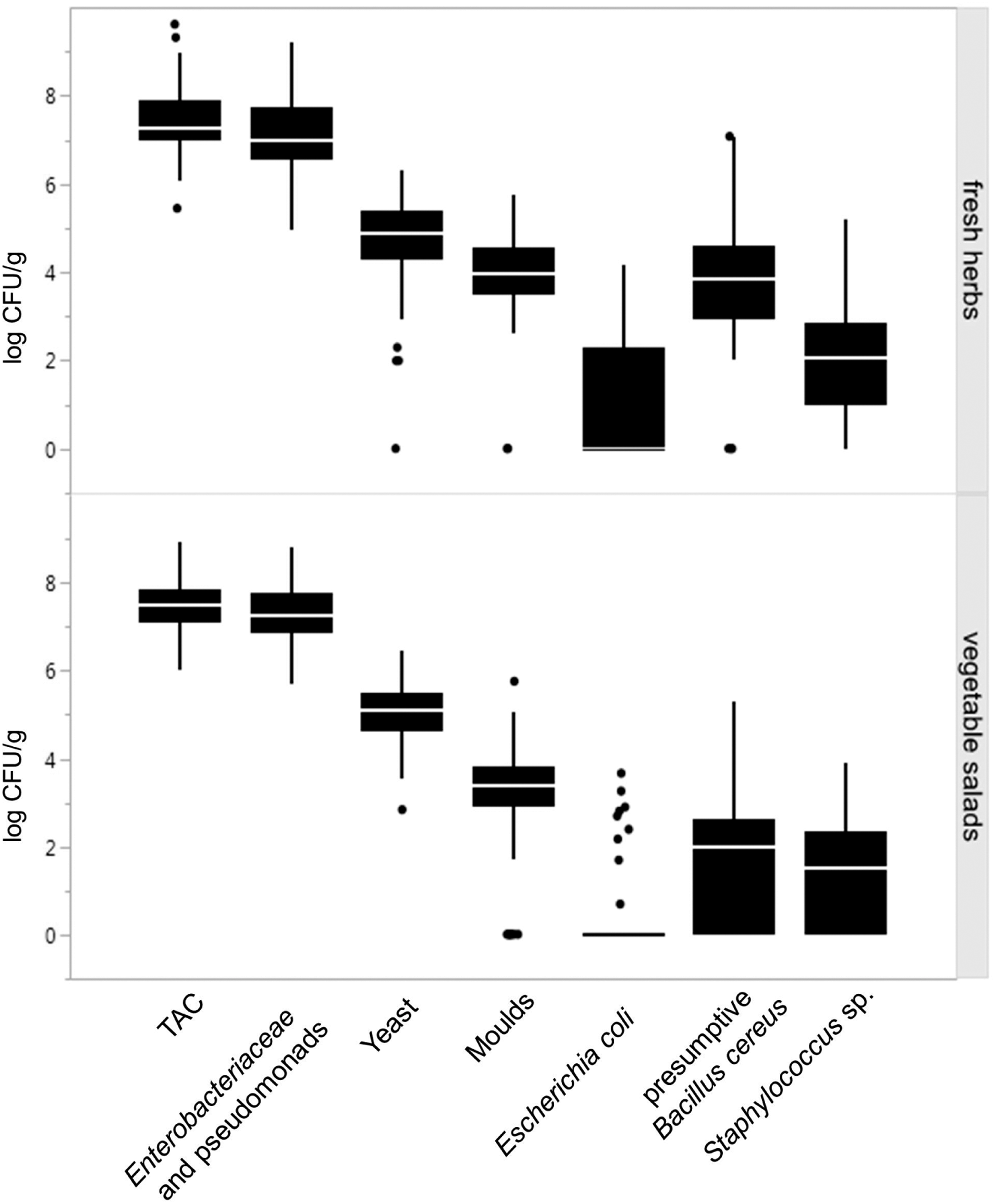
Box plots indicating the distribution of log CFU/g counts of TAC, Enterobacteriaceae and pseudomonads, yeast, moulds, Escherichia coli, presumptive Bacillus cereus, and Staphylococcus spp. determined in fresh herbs (upper part) and vegetable salads (lower part). CFU, colony-forming unit; TAC, total aerobic mesophilic plate count.
Comparing the results of conventionally- and organically-cultivated herbs, no significant differences with regard to TAC and counts of enterobacteria and pseudomonads, yeast, presumptive B. cereus, and staphylococci were observed (Fig. 2A). In the organically-cultivated herbs, the mean count of moulds was 0.7 log CFU/g higher. In contrast to the conventionally-cultivated herbs with 1.4 log CFU/g, no E. coli could be detected in the organically-produced herbs.

Bar plots indicating the mean values and standard errors of log CFU/g counts of TAC, Enterobacteriaceae and pseudomonads, yeast, moulds, Escherichia coli, presumptive Bacillus cereus, and Staphylococcus spp. Significantly differing pairs are labeled by an asterisk, indicating a p-value <0.05.
Microbiological quality of fresh vegetable salads
The levels of TAC in leafy salad samples ranged from 6.0 to 8.9 log CFU/g (Fig. 1; Supplementary Tables S2 and S4). The counts of enterobacteria and pseudomonads ranged from 5.7 to 8.8 log CFU/g. This microbial group was predominant and could be detected in all salad samples. The yeast and mould counts ranged between 2.9 and 6.5 log CFU/g and from below detection limit to 5.8 log CFU/g, respectively. Strains of E. coli could be detected above 3.0 log CFU/g in two samples. Staphylococci reached counts up to 3.9 log CFU/g; however, no coagulase-positive S. aureus was detected. Presumptive B. cereus strains were present in 71 samples (62.3%) with counts up to 5.3 log CFU/g.
Two conventionally-produced samples were positive for L. monocytogenes; the three herein obtained isolates were confirmed by API Listeria and characterized by multiplex PCR to serogroup IIa (containing serotypes 1/2a and 3a). Additionally, five isolates of the nonpathogenic species L. innocua were obtained from one organic sample. RAPD analysis for bacterial strain typing showed low genetic diversity among L. innocua isolates (Supplementary Fig. S1). Six suspicious Salmonella spp. isolates obtained from one organic and two conventional salad samples were identified as Salmonella enterica subsp. enterica serovar Enteritidis (serotype (1),9,12:g,m:-). The draft genome sequence of Salmonella isolate MS 501 was previously published (Stoll et al., 2018).
No significant differences in the microbial counts were detected between conventionally- and organically-produced salads except yeast values, which were 0.3 log higher in conventionally-grown salads (Fig. 2B).
The DGHM [Deutsche Gesellschaft für Hygiene und Mikrobiologie (German Society for Hygiene and Microbiology)] publishes guidance and critical values for specific food products. These values can be regarded as recommendations concerning microbial quality and hygiene. The results of all 74 salads labeled as RTE were assessed based on the 2016 DGHM values. The guidance values were reached or exceeded: in 43.2% for TAC and in 70.3% for yeast. The critical values for E. coli, moulds, and presumptive B. cereus were reached or exceeded in 6.8%, 13.5%, and 20.3%, respectively.
A comparison between RTE salads and salads that needed further processing showed significant differences (Fig. 2C). In the RTE salads, the values for TAC, as well as for enterobacteria, pseudomonads, and yeast were 0.3, 0.5, and 0.6 log higher, respectively. However, the mould counts were even 0.7 log higher in the products that needed processing before consumption.
Discussion
In several studies dealing with the microbiological quality of these products as well as in the present study, a high microbial load was described (Abadias et al., 2008; De Oliveira et al., 2011; Jeddi et al., 2014; Fiedler et al., 2017). Such products have been implicated as vehicles for the transfer of microbial foodborne diseases worldwide (Beuchat, 2006; FAO and WHO, 2008).
Many freshly packaged salads are labeled with the information that these could be consumed directly without washing, that is, RTE. In this study, we could detect significantly higher average microbial counts for TAC, enterobacteria, pseudomonads, and yeast in RTE salads in contrast to products that still needed processing before consumption. Processing including, for example, cutting creates a favorable environment for the growth of microorganisms, which is even enhanced during storage (Allende et al., 2004; Saini et al., 2017).
The overall microbial quality of foods is often assessed by determining TAC. However, this is not a reliable indicator for the sanitary and sensorial quality of leafy greens when considered without further analyses (Ceuppens et al., 2015). A major part of the microbiota associated with fresh leafy salads and herbs is a natural epiphytic microbiota, which is mainly nonpathogenic.
The TAC mean values of both types of products were 7.8 and 8.2 log CFU/g. Populations of mesophilic aerobic bacteria up to 8 log CFU/g were commonly reported on fresh produce at various points in the food chain (FAO and WHO, 2008). It is commonly agreed that TAC levels ≤7.0 log CFU/g are acceptable for fresh products. Compared with other studies (Johnston et al., 2005; Abadias et al., 2008; Seo et al., 2010; Najafi et al., 2012), we detected samples with a higher TAC: 70.6% of all herb samples and even 81.5% of all salad samples showed TAC levels ≥7.0 log CFU/g.
We were able to confirm that pseudomonads and enterobacteria constituted the dominant microbiota of all investigated products and could be detected in every sample with counts between 5.0 and 9.2 log CFU/g. Al-Kharousi et al. (2016) described that enterobacteria were present in 91% of vegetable samples. Najafi et al. (2012) reported that 100% of mixed fresh-cut salads and mixed green leafy vegetables were contaminated with Enterobacteriaceae, and the level of contamination varied from 3.0 to 8.3 log CFU/g. Abadias et al. (2008) showed that the incidence of Enterobacteriaceae in whole and fresh-cut vegetables was 78.6%.
As B. cereus sensu lato are known as typical soil inhabitants (Granum et al., 2005; Stenfors Arnesen et al., 2008; Gekenidis et al., 2017), soil depicts a probable contamination source. B. cereus levels >103 or 104 CFU/g are considered potentially hazardous (NSW Food Authority, 2009; DGHM, 2016). In our study, 7.1% salad samples (n = 8) and 49.4% herb samples (n = 37) were contaminated with presumptive B. cereus in counts equal to or higher than 4.0 log CFU/g. According to the guidance and critical values for microbial criteria of the DGHM, 19 salad samples (16.7%) exceeded the critical value of 3.0 log CFU/g; 4 of those were organically-cultivated, while 15 were conventionally-produced.
Listeria monocytogenes was detected only in 2 (1.8%) conventionally-produced salad samples. The critical value of DGHM for L. monocytogenes and the European Union limit of 100 CFU/g was not exceeded in any of the samples. Obtained isolates were confirmed as belonging to PCR serogroup IIa (including serotypes 1/2a and 3a). The results of our study of fresh produce obtained in Southwest Germany indicated that the incidence of L. monocytogenes was low. Similar incidence was obtained within one study in northern Germany (Fiedler et al., 2017). The authors determined an incidence of 1.0% of L. monocytogenes in fresh produce. PCR serogrouping showed that the two isolates from two mixed salad samples belonged to serogroups IIb (containing serovars 1/2b, 3b, and 7) and IVb (containing serotypes 4b, 4ab, 4d, and 4e), respectively.
Meloni et al. (2009) examined 50 prepackaged mixed vegetable salads marketed in Italy for the occurrence of L. monocytogenes and detected only one L. monocytogenes strain belonging to serovar 1/2b. Comparable incidences of L. monocytogenes have already been reported in vegetable salads, for example, 3.2% in RTE vegetables in Brazilian markets (Porto and Eiroa, 2001) and 4.8% in raw vegetable salads in the United Kingdom (Little et al., 2007). In contrast, L. monocytogenes was detected in 22.5% of minimally-processed vegetable samples within a study in Malaysia (Ponniah et al., 2010).
According to German, European, and international regulations (Commission of the European Communities, 2005; NSW Food Authority, 2009; DGHM 2016), the presence of Salmonella in RTE vegetables is unacceptable in 25 g of sample material. Salmonella outbreaks associated with fresh plant produce have been described frequently worldwide (Berger et al., 2010; Food Safety Authority of Ireland, 2015). Especially the outbreaks with Salmonella Typhimurium, Thompson, Anatum, Saintpaul, and Senftenberg were associated with RTE plant produce (Berger et al., 2010).
In our study, Salmonella was detected in 2.6% of all salad samples, and all six isolates belonged to serovar Enteritidis. Within a parallel study in northern Germany (Fiedler et al., 2017), one Salmonella isolate (incidence 0.5%) was identified as Salmonella Szentes. Low prevalence of Salmonella in minimally processed leafy green vegetables has been also reported in other countries (Abadias et al., 2008; Berger et al., 2010; Sant'Ana et al., 2011).
In a recently published study (Szczech et al., 2018), the microbial quality of organic and conventional vegetables in Poland was investigated, and significant differences could be detected. In our study, the microbial loads were similar between organically- and conventionally-cultivated produce with regard to TAC and counts of enterobacteria, pseudomonads, and staphylococci. However, significant differences were observed for the counts of yeast in salads. In herbs, significant differences could be detected for counts of moulds, and E. coli could only be detected in conventionally-grown herbs.
Conclusions
Our results showed that leafy salads and herbs may contain pathogenic bacteria, especially L. monocytogenes and Salmonella; however, the incidence in the produce sold in Southwest Germany in 2015 was very low. A high level of contamination of mesophilic aerobic bacteria, and a high occurrence of presumptive B. cereus was observed. Under nonoptimal storage conditions, the microbiota present in these products may increase and constitute a certain health risk for consumers, especially for people with a weakened immune system, namely children and elders. Therefore, it is important to minimize the microbiota by washing or heating (e.g., herbs) and paying attention to good preparation hygiene. In addition, it should be considered that a low storage temperature is maintained and storage time is reduced to a minimum.
Footnotes
Acknowledgments
The authors acknowledge the technical assistance of Luisa Martinez, Lilia Rudolf, Andrea Schavitz, and Lena Schmid. They also thank the National Reference Laboratory of Salmonella at the German Federal Institute for Risk Assessment (BfR) for serotyping the Salmonella isolates. This study was supported by the German Federal Ministry of Food and Agriculture; the work was conducted within the project “Humanpathogene in der pflanzlichen Erzeugung: Status quo, Dekontamination, Eintragswege und Einfluss der Lagerungsbedingungen.”
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.