
Abstract
Coenzyme Q0 (CoQ0), one of benzoquinone compounds, has been demonstrated to possess antineoplastic, anti-inflammatory and antioxidant activities. However, its antimicrobial effect has not been extensively reported. In this study, antimicrobial activity of CoQ0 against Staphylococcus aureus was evaluated by measurement of inhibition zone, minimum inhibitory concentration (MIC), and growth curves. Time-kill assay was performed to assess the bactericidal activity of CoQ0 against S. aureus in tryptone soya broth and pasteurized milk. The possible mechanism of action was explored through measuring changes in intracellular ATP concentrations, membrane potential, and cell morphology. Furthermore, propidium iodide (PI) staining assay was performed to evaluate the effect of CoQ0 on cell membrane integrity. The MIC of CoQ0 against tested strains ranged from 7.8 to 62.5 μg/mL. CoQ0 at 2 × MIC showed bactericidal effect on S. aureus in tryptic soy broth (TSB) and pasteurized milk. Decrease in intracellular ATP concentration and membrane potential were detected when cells were treated with CoQ0. PI staining demonstrated destruction of bacterial cell membrane. CoQ0 also induced abnormal cell morphological changes, as confirmed by field emission scanning electron microscopy. These findings suggested that CoQ0 exhibited antimicrobial effect on S. aureus, which was partly because of its ability to damage cell membrane.
Introduction
Staphylococcus aureus, a Gram-positive microorganism, is adaptive to various environments and could survive in dry conditions, such as human skin, clothing, and contact surfaces (Chaibenjawong and Foster, 2011). S. aureus could contaminate many types of food, including raw meat (Wang et al., 2013), meat products (Chokesajjawatee et al., 2009), raw milk and milk products (Gündoğan et al., 2006; Jakobsen et al., 2011), and prepared and frozen foods (Baek et al., 2009; Wang et al., 2017). Milk, especially raw milk, was frequently contaminated by S. aureus and has become an important source of S. aureus food poisoning. Pasteurized milk, sold in supermarket, retail store, farmer market, and milk bar, could also be contaminated by S. aureus because of damaged package during transportation, storage, and marketing or exposure to air, operator, container, and other contact surfaces (Li et al., 2017). S. aureus has imposed huge public health hazard in milk and milk products. Besides, S. aureus is one of the common human pathogens and could cause skin disorders, septicemia, and toxic shock syndrome (Qiu et al., 2010). Therefore, it is extremely vital to seek effective methods to control S. aureus. Traditionally, chemical compounds have been widely applied as antimicrobial agents or preservatives to prevent contamination caused by pathogenic bacteria or retard their growth. However, the potential adverse effects of these synthetic chemicals on human health have made it increasingly unfavorable for consumers. Recently, natural compounds derived from plants, animals, or microorganisms have attracted increasing attention. Clove essential oils (Kovacs et al., 2016), chitosan (Goy et al., 2016), and bacteriocin (Arief et al., 2012) were reported to be effective in inhibiting growth of certain pathogenic microorganisms.
Coenzyme Q0 (2,3-dimethoxy-5-methyl-1,4-benzoquinone [CoQ0]), a kind of benzoquinone compounds, could be extracted from fungus Antrodia cinnamomea (Chung et al., 2014). Although it has been proved that CoQ0 has antineoplastic, anti-inflammatory, and antiangiogenic properties (Yang et al., 2015, 2016; You-Cheng et al., 2016), its antimicrobial and bactericidal activity against pathogenic microorganisms has not been extensively explored.
The objective of this study was to investigate the effect of CoQ0 on S. aureus growth and its possible mechanism. Agar dilution method was used for minimum inhibitory concentration (MIC) determination, and time-kill assay was conducted to evaluate bactericidal activity of CoQ0 against S. aureus. Changes in membrane potential, intracellular ATP concentrations, membrane integrity, and cell morphology of S. aureus were studied to elucidate possible mode of action of CoQ0 against S. aureus.
Materials and Methods
Chemical reagents
CoQ0 (High Performance Liquid Chromatography ≥99%, CAS 605-94-7) was obtained from J&K Scientific Co., Ltd. (Beijing, China). CoQ0 was first dissolved with dimethyl sulfoxide (DMSO) and then diluted with sterile phosphate-buffered saline (PBS) or medium. The final concentration of DMSO in all samples including the control group was 1% (v/v). Tryptic soy agar (TSA) and tryptic soy broth (TSB) were purchased from Land Bridge Technology Co., Ltd. (Beijing, China). The pasteurized milk was obtained from local supermarket.
Bacterial strains and culture conditions
S. aureus ATCC 25923 and ATCC 29213 were purchased from American Type Culture Collection (ATCC, Manassas, VA). Ten other S. aureus isolates were obtained from our laboratory strain collection and originally isolated from food samples in China. The MIC assay was conducted for all 12 S. aureus strains. Other assays were performed for ATCC 25923 only. Tested strains were inoculated into TSB and incubated at 37°C for 12 h. After centrifugation (4°C, 8000 × g, 5 min), bacterial cells were washed three times with PBS and resuspended in TSB to obtain S. aureus working suspension with an optical density at 600 nm (OD600) ≈ 0.5.
Measurement of inhibition zone
To explore the antimicrobial activity of CoQ0 preliminarily, inhibition zone measurement for S. aureus ATCC 25923 was performed by agar diffusion method (Yi et al., 2010). S. aureus suspension (OD600 ≈ 0.5) was diluted 100-fold with PBS, and 100 μL of the suspension was spread onto TSA. After sterile Oxford cups were put on TSA plate, CoQ0 solutions of different concentrations (0, 1.25, 2.5, and 5 mg/mL) were, respectively, added into Oxford cups. Ampicillin (0.1 mg/mL) was used as positive control. The plates were incubated at 37°C for 24 h and the diameter of inhibition zone was measured.
Minimum inhibitory concentration
In this assay, agar dilution method (Griffin et al., 2000) was followed with minor modifications. First, sterile molten TSA (50–55°C) was added into 24-well plate, followed by addition of CoQ0. The final concentration of CoQ0 in each well was 0 (control), 3.9, 7.8, 15.6, 31.2, 62.5, 125, 250, and 500 μg/mL, respectively. After dilution, S. aureus suspension (2 μL) was dropped on the center of each well. Then the plates were placed for a while to dry before being incubated at 37°C for 24 h. The lowest concentration of CoQ0 at which there was no growth of S. aureus strains was defined as MIC.
Growth curve determination
Growth curve determination was performed to investigate effect of CoQ0 on S. aureus ATCC 25923 growth. S. aureus suspension (OD600 ≈ 0.2) was added into 96-well plate, followed by addition of CoQ0 to obtain its final concentration of 0 (control), 1/16 × MIC, 1/8 × MIC, 1/4 × MIC, 1/2 × MIC, and 1 × MIC. Then the plate was incubated at 37°C for 24 h, and the OD600 was determined at 1-h intervals by a multimode plate reader (Infinite™ M200 PRO; Tecan, Mannedorf, Switzerland).
Time-kill assay
The time-kill assay was performed to reflect bactericidal activity of CoQ0 against S. aureus in TSB and pasteurized milk as previously described (Shi et al., 2016). S. aureus ATCC 25923 was inoculated separately into sterile TSB and pasteurized milk to achieve bacterial concentration of 105 colony-forming unit (CFU)/mL approximately. CoQ0 was added into TSB and pasteurized milk, and its final concentrations were 0 (control), 1/2 × MIC, 1 × MIC, and 2 × MIC, respectively. Then samples were incubated at 37°C for 24 h. During this period, samples (100 μL) were taken at 0, 3, 6, 9, 12, and 24 h for viable plate counting.
Intracellular ATP concentrations
The intracellular ATP concentrations were measured as previously described (Turgis et al., 2009). After S. aureus suspension was added into Eppendorf tubes, CoQ0 was added to obtain final concentrations of 0 (control), 1 × MIC, 2 × MIC, and 4 × MIC, respectively. The tubes were incubated at 37°C for 30 min, followed by centrifugation (8000 × g, 4°C, 5 min) and removal of supernatant. Bacterial cells were treated with lysis buffer for breaking and a vortex was performed to speed the lysis of cells. Then bacterial suspension was centrifuged and the supernatant was preserved on ice for subsequent use. ATP assay kit (Beyotime, Shanghai, China) was used to measure ATP concentrations in the supernatant. ATP working solution (100 μL) was mixed with 100 μL supernatant in the black, opaque, and flat-bottomed 96-well plates (Corning, Kennebunk, ME), followed by measurement of luminescence intensity by using microplate reader (Infinite M200 PRO; Tecan). Results were expressed in relative luminescence unit, which represents the luminescence intensity that has been background deducted.
Membrane potential
Bis-(1,3-dibutylbarbituric acid) trimethine oxonol (DiBAC4(3); Sigma), a fluorescent dye sensitive to membrane potential, was used in this assay (Sanchez et al., 2010). S. aureus ATCC 25923 working suspension (OD600 ≈ 0.5) was added into black, opaque, flat-bottomed 96-well plate, followed by addition of DiBAC4(3) and incubation at 37°C in the dark for 30 min. Next, CoQ0 was added into wells and its final concentration was 0 (control), 1 × MIC, 2 × MIC, and 4 × MIC, respectively. Then the plate was incubated at 37°C in the dark for 30 min, and a microplate reader (Infinite M200 PRO; Tecan) was used to detect fluorescence intensity at excitation and emission wavelengths of 492 and 515 nm, respectively. Results were expressed in relative fluorescence unit, which represents the fluorescence intensity that has been background subtracted.
Membrane integrity
With minor modifications, propidium iodide (PI) staining method (Yan et al., 2016) was applied to evaluate the integrity of S. aureus cell membrane. ATCC 25923 working suspension (OD600 ≈ 0.5) was diluted 100-fold with PBS before the suspension was pipetted into 24-well plate. CoQ0 was added into wells and its final concentrations was 0 (control), 1 × MIC, 2 × MIC, and 4 × MIC, respectively, followed by incubation at 37°C for 3 h. Then PI solution was added into each well and the plate was kept at room temperature for 20 min. Samples were observed and photographed by using an inverted fluorescence microscope (E50I; Nikon).
Scanning electron microscopy observation
First, CoQ0 was added into S. aureus ATCC 25923 working suspension (OD600 ≈ 0.5) to obtain the final concentration of 0 (control), 1 × MIC, 2 × MIC, and 4 × MIC, respectively. Then the suspension was incubated at 37°C for 3 h, centrifuged at 5000 × g for 10 min, washed twice with PBS, and fixed with glutaraldehyde (2.5%) at 4°C for 10 h. After centrifugation at 3000 × g for 10 min, bacterial cells were washed three times and postfixed with 1% of osmic acid. Then cells were dehydrated by water–ethanol solutions of 30%, 50%, 70%, 80%, 90%, and 100% for 10 min, respectively. The bacterial suspension (5 μL) dehydrated by absolute ethanol was dropped on sterile glass slide to prepare samples for observation, and scanning electron microscope (S-4800; Hitachi, Tokyo, Japan) was used to observe morphology of samples.
Statistical analysis
All experiments were performed independently for three times. All data are expressed as mean ± standard deviation. One-way analysis of variance and t-test by SPSS software were used to conduct statistical analysis. Difference was considered significant at p ≤ 0.05.
Results
Diameter of inhibition zone
CoQ0 at the concentration of 1.25, 2.5, and 5 mg/mL resulted in inhibition zones of 19.8 ± 0.2, 24.2 ± 0.4, and 30.8 ± 0.2 mm, respectively. Ampicillin (0.1 mg/mL) exerted an inhibition zone of 32.3 ± 0.6 mm (data not given). It was suggested that CoQ0 possesses antimicrobial activity against S. aureus ATCC 25923.
Minimum inhibitory concentration
The MICs of CoQ0 against 12 S. aureus strains are given in Table 1. It was evidenced that CoQ0 exhibited good antibacterial effect on tested S. aureus with MICs ranging from 7.8 to 62.5 μg/mL.
Minimum Inhibitory Concentrations of Coenzyme Q0 Against Staphylococcus aureus Strains
S. aureus suspension was inoculated on TSA in which the final concentration of CoQ0 was 0 (control), 3.9, 7.8, 15.6, 31.2, 62.5, 125, 250, and 500 μg/mL, respectively. Samples were incubated at 37°C for 24 h. The lowest concentration of CoQ0 at which there was no growth of S. aureus strains was defined as MIC.
ATCC, American Type Culture Collection; CoQ0, coenzyme Q0; MICs, minimum inhibitory concentrations; TSA, tryptic soy agar.
Growth curve
Growth curves of S. aureus ATCC 25923 in the presence of CoQ0 are given in Figure 1. CoQ0 at 1 × MIC showed a strong inhibitory effect on the growth of S. aureus, with no significant increase of optical density values over 24 h (p > 0.05). CoQ0 at 1/2 × MIC reduced the growth rate of S. aureus. In addition, the growth curve of S. aureus treated with CoQ0 at low concentrations (1/4 × MIC to 1/16 × MIC) was similar to that of the control, suggesting that CoQ0 at 1/4 × MIC to 1/16 × MIC exerted no significant effect on S. aureus growth (p > 0.05).
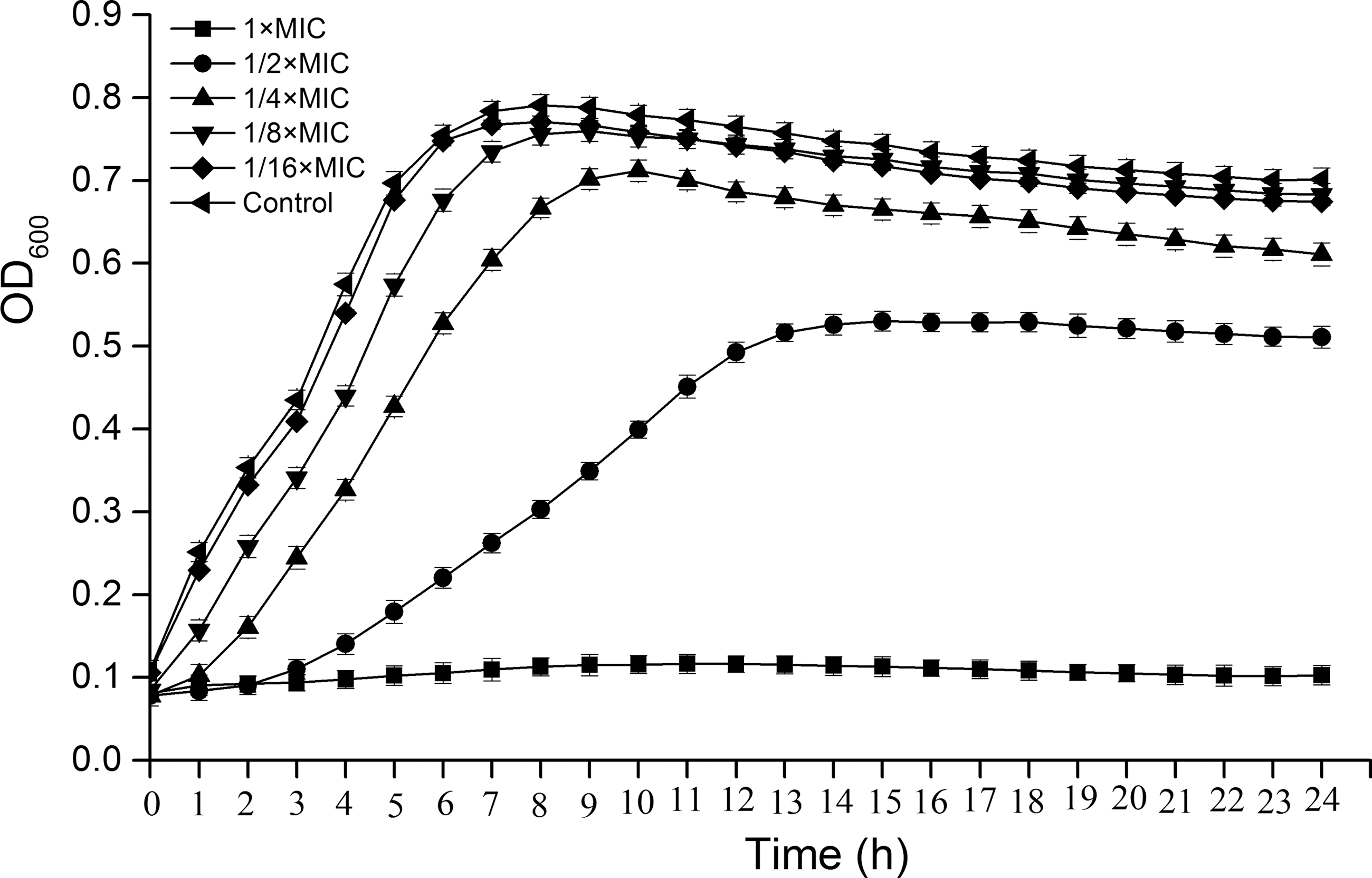
Growth curves of Staphylococcus aureus ATCC 25923 in the presence of CoQ0 (1 × MIC to 1/16 × MIC). S. aureus suspension was treated with CoQ0 at the concentration of 0 (control), 1/16 × MIC, 1/8 × MIC, 1/4 × MIC, 1/2 × MIC, and 1 × MIC for 24 h. Error bars represent standard deviation of three replicates. ATCC, American Type Culture Collection; CoQ0, coenzyme Q0; MIC, minimum inhibitory concentration; OD600, optical density at 600 nm.
Time-kill curves
As given in Figure 2, CoQ0 exhibited better bactericidal activity in pasteurized milk than in TSB. At each time point, CoQ0 at 2 × MIC resulted in a greater reduction of viable counts in pasteurized milk than in TSB. Viable cells in TSB and pasteurized milk was reduced below the detection limit after treatment with CoQ0 (2 × MIC) for 24 h. The initial number of viable bacterial cells in TSB and pasteurized milk was 5.3 log CFU/mL. After treatment with CoQ0 at 1 × MIC, viable cell counts in TSB were reduced to 3.6 log CFU/mL at 12 h, and increased in the following 24 h. In contrast, viable cells counts in pasteurized milk were reduced to 2.8 and 1.0 log CFU/mL at 12 and 24 h, respectively. In addition, in TSB and pasteurized milk, CoQ0 at 1/2 × MIC did not result in the decrease of viable cell counts. These data indicated that CoQ0 at 2 × MIC exerted good bactericidal effect on S. aureus ATCC 25923 in both TSB and pasteurized milk, whereas CoQ0 at 1/2 × MIC showed no inactivation effect on S. aureus ATCC 25923 cells both in TSB and pasteurized milk, but merely slowed down its growth rate.
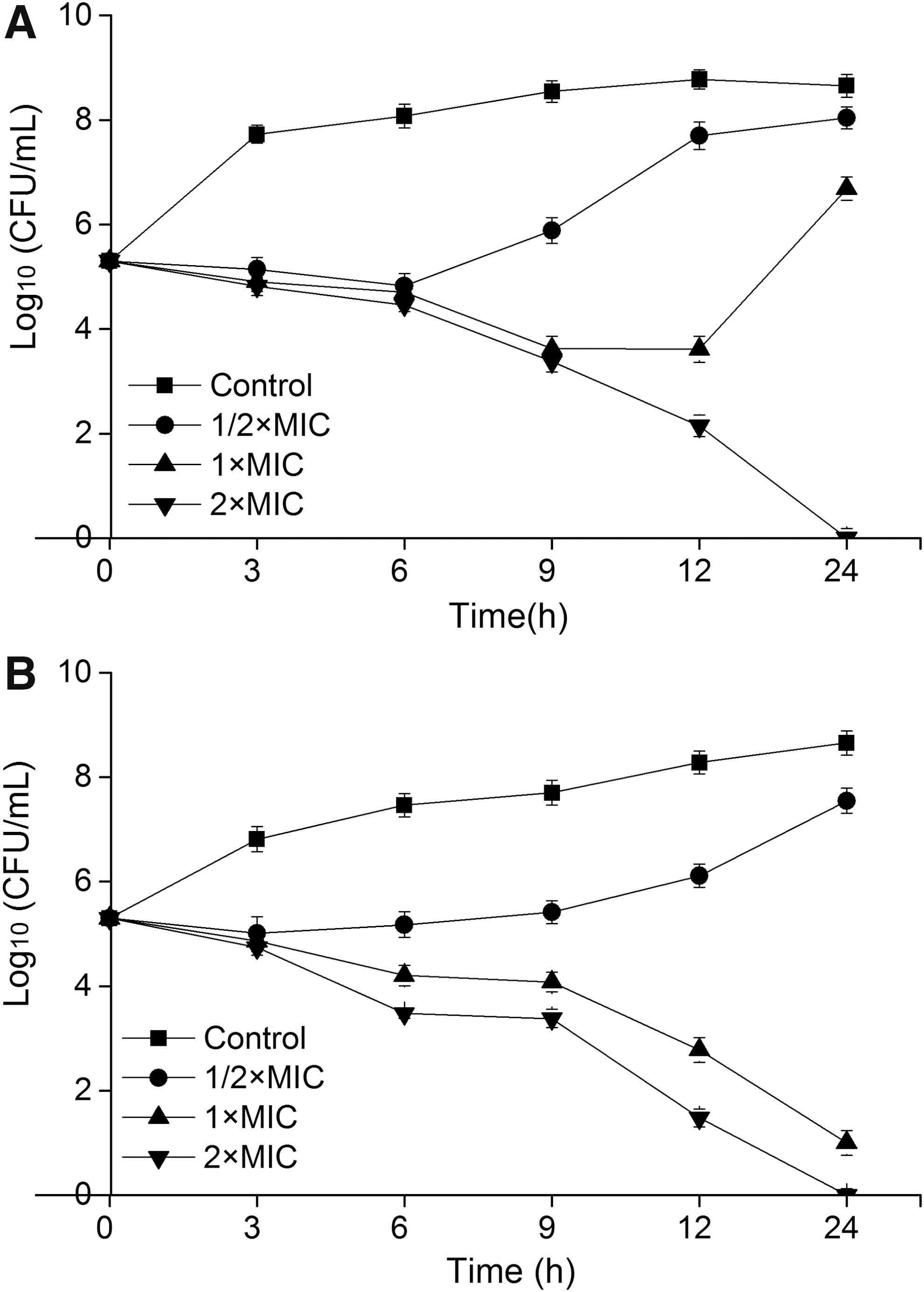
Time-kill curves of CoQ0 against Staphylococcus aureus ATCC 25923 in TSB
Intracellular ATP concentrations
Changes in intracellular ATP concentrations of S. aureus ATCC 25923 after treatment with CoQ0 are given in Figure 3. Compared with the control, a significant decrease of ATP concentration inside S. aureus cells occurred after exposure to CoQ0 (p ≤ 0.01), and there was no significant difference in ATP concentration change as the concentration of CoQ0 increased from 1 × MIC to 4 × MIC (p > 0.05). The ATP concentration of control group was 0.990 μmol/L, and the values decreased to 0.087, 0.058, and 0.050 μmol/L in the presence of CoQ0 at 1 × MIC, 2 × MIC, and 4 × MIC, respectively.
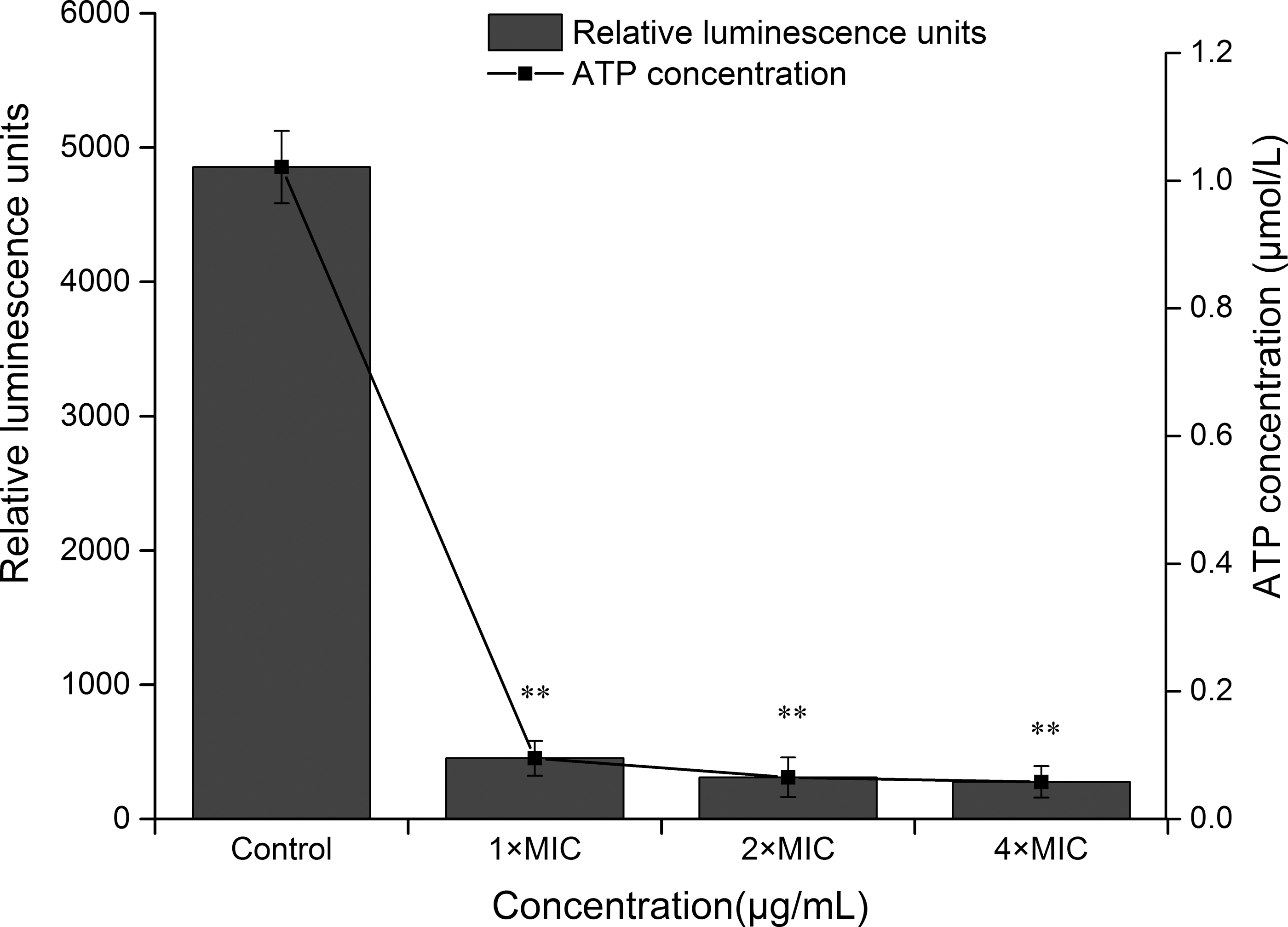
Effect of CoQ0 on intracellular ATP concentration of Staphylococcus aureus ATCC 25923. Samples were treated with CoQ0 at the concentration of 0 (control), 1 × MIC, 2 × MIC, and 4 × MIC. The relative luminescence intensity of control was 4855, and the corresponding intracellular ATP concentration was 0.99 μmol/L. Error bars indicate the SD. Values represent means of three independent experiments ± SD. **p < 0.01 versus the control group. ATCC, American Type Culture Collection; CoQ0, coenzyme Q0; MIC, minimum inhibitory concentration; SD, standard deviation.
Membrane potential
The membrane potential was examined by measuring the fluorescence intensity of samples with or without CoQ0 treatment. The effect of CoQ0 on membrane potential of S. aureus ATCC 25923 are given in Figure 4. Compared with the control, the fluorescence intensity of samples experienced a significant reduction as the concentration of CoQ0 increased (p ≤ 0.05), which indicated that S. aureus membrane potential was reduced by CoQ0, and hyperpolarization in S. aureus cells occurred.
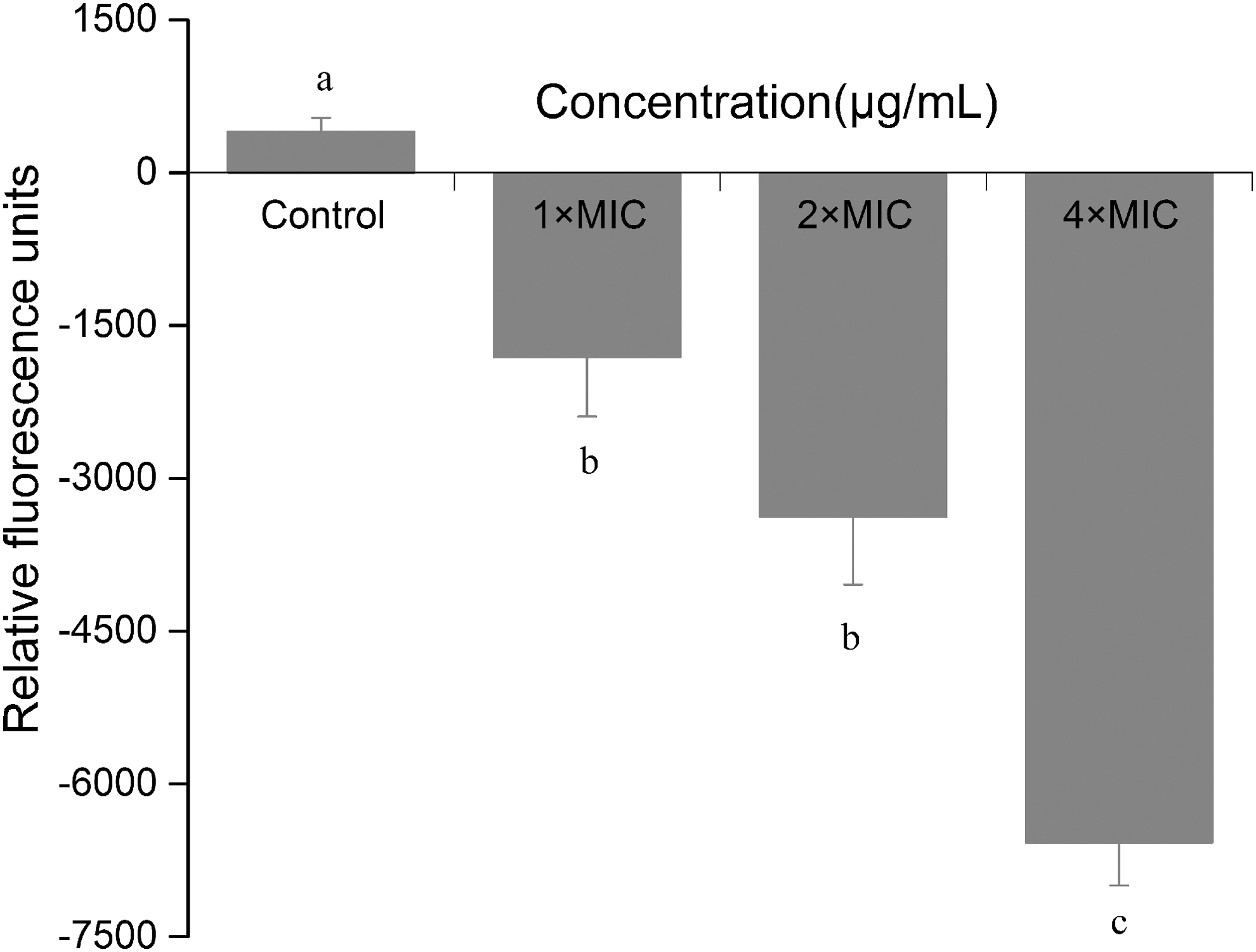
Effect of CoQ0 on membrane potential of Staphylococcus aureus ATCC 25923. Samples were treated with CoQ0 at the concentration of 0 (control), 1 × MIC, 2 × MIC, and 4 × MIC. The relative fluorescence intensity of control was 402. Error bars indicate SD. Values represent means of three independent experiments ± SD. Different letters represent statistically significant difference at p ≤ 0.05. ATCC, American Type Culture Collection; CoQ0, coenzyme Q0; MIC, minimum inhibitory concentration; SD, standard deviation.
Integrity of cell membrane
The red fluorescence intensity was used to evaluate the integrity of S. aureus cell membrane. As given in Figure 5, there was nearly no red fluorescence in the sample treated without CoQ0 (Fig. 5A). However, the intensity of red fluorescence was increasingly enhanced as the concentration of CoQ0 increased from 1 × MIC to 4 × MIC. Samples exposed to CoQ0 at 4 × MIC exhibited the strongest red fluorescence intensity, followed by 2 × MIC and 1 × MIC (Fig. 5B–D). The results suggested that CoQ0 impaired integrity of S. aureus cell membrane in a dose-dependent manner.

Fluorescence microscopy photographs of Staphylococcus aureus ATCC 25923 treated with CoQ0 for 3 h at the concentrations of 0
Scanning electron microscopy observation
The effect of CoQ0 on S. aureus cell morphology was analyzed by scanning electron microscopy (Fig. 6). Bacterial cells without CoQ0 treatment exhibited a regular, smooth, and well-rounded morphology (Fig. 6A), whereas smaller cell size and shrinked, pitted, and irregular cell surface occurred as the concentration of CoQ0 increased (Fig. 6B–D).

Scanning electronic images of Staphylococcus aureus ATCC 25923 after treatment with CoQ0 for 3 h at different concentrations of 0
Discussion
Many natural compounds possessed inhibitory effect on S. aureus growth. Previous studies demonstrated the MIC values of Cinnamon essential oil (Zhang et al., 2016), punicalagin (Xu et al., 2017), and chitosan (Yan et al., 2016) against S. aureus were 1.0, 0.25, and 0.25 mg/mL, respectively. In this study, CoQ0 inhibited growth of S. aureus with MICs ranging from 7.8 to 62.5 μg/mL. These results suggest that CoQ0 exhibits stronger antimicrobial effectiveness against S. aureus than those aforementioned compounds. In addition, CoQ0 exhibited better bactericidal activity in pasteurized milk than in TSB, which may be the result of the better growth of S. aureus in TSB than in milk or the interactions of CoQ0 with milk components. The findings also suggested that CoQ0 has the potential to be applied as natural preservative in food processing.
Cytoplasmic membrane plays a vital role in maintaining normal physiological and metabolic activities and keeping vitality. Typical characteristics, including obvious changes in membrane potential, decrease of intracellular ATP concentration, leakage of intracellular materials, impaired membrane integrity, and abnormal morphological alterations, will occur when bacterial cell membrane is subjected to damage (Stojanovic-Radic et al., 2012; Li et al., 2013). These characteristics could be used to evaluate whether damages occur to cell membrane and have been widely used as indicators to assess effectiveness of antimicrobial agents and to elucidate associated mechanism of action. In this study, membrane potential and intracellular ATP concentration of S. aureus were significantly reduced by CoQ0, which was in agreement with other natural compounds reported previously. S. aureus and Escherichia coli experienced reduction of 69.61% and 98.27%, respectively, in intracellular ATP concentration after treatment with salvia sclarea oil (Cui et al., 2015). According to Li et al. (2013), chlorogenic acid reduced membrane potential of S. aureus and caused hyperpolarization. The disruption of ion and energy balance provoked by increased membrane permeability and loss of ATP may be responsible for this change in membrane potential (Murdock et al., 2010).
It has been reported that many benzoquinone compounds, especially naturally occurring ones, exhibited significant antimicrobial potential against pathogenic bacteria. For example, 1,4-benzoquinone, 2,6-dimethoxy-1,4-benzoquinone, methoxybenzoquinone, hydroquinone, 2,5-dihydroxy-3-ethyl-2,5-cyclohexadiene-1,4-dione, and alkylbenzoquinone derivatives from Ardisia kivuensis showed antibacterial activity against S. aureus with MICs of 8, 8, 16, 64, 64–256, and 6.25–25 μg/mL, respectively (Kim et al., 2010; Paul et al., 2014; Omosa et al., 2016). In contrast, CoQ0 had a different MIC of 7.8–62.5 μg/mL against S. aureus. This may result from different structure among the benzoquinone compounds. According to previous reports, the antibiotic effects of quinones depended on the substituents at the 3 and 6 positions of the 2,5-dihydroxy-1,4-benzoquinone ring and the activity enhanced with increase in side chain length (Omosa et al., 2016). In addition, characteristic groups and their positions are critical for the antibacterial activity. For benzoquinone compounds, the correlations between structure and antimicrobial activity seem to be an interesting topic for further studies.
Conclusions
In this study, CoQ0 exhibited good antimicrobial activity against S. aureus with MICs ranging from 7.8 to 62.5 μg/mL. CoQ0 at 2 × MIC showed bactericidal effect on S. aureus in both TSB and pasteurized milk, especially in pasteurized milk. After CoQ0 treatment, decrease of membrane potential and intracellular ATP concentration, impaired cytoplasmic membrane integrity and adverse morphological changes occurred, which indicated that CoQ0 exerted antimicrobial and bactericidal effect on S. aureus possibly by damaging cell membrane. These findings suggest that CoQ0 has the potential to serve as an effective antimicrobial agent to control S. aureus. In future studies, the effect of CoQ0 on S. aureus enterotoxin production could be assessed.
Footnotes
Acknowledgments
This work was supported partly by the Fundamental Research Funds for the Central Universities (2452017146 and 2452017228), National Natural Science Foundation of China (31772084, 31801659 and 31801658), and State Key Laboratory Breeding Base for Zhejiang Sustainable Pest and Disease Control (2010DS700124-ZM1608).
Disclosure Statement
No competing financial interests exist.