
Abstract
Eggs are a healthy and nutritious food source, but may be contaminated by bacteria. Previous studies have reported the presence of staphylococci in eggs of farmed chickens, but no study has evaluated the staphylococcal population of eggs from household chickens. In this study, staphylococci from eggs (n = 275) of household chickens collected from November 2016 to March 2017 from different villages of Khyber Pakhtunkhwa province, Pakistan, were characterized. Seven species of staphylococci were identified from 65 eggs, including the predominant species, Staphylococcus xylosus (49/275; 17.8%). S. xylosus isolates (n = 73) were tested for antimicrobial susceptibility, presence of resistance genes, genetic relatedness, and inhibitory activity against other bacteria. The majority of isolates were resistant to oxacillin (83.6%) and tetracycline (24.7%), but also exhibited resistance to daptomycin and linezolid (5.5% each). Of the 10 resistance genes tested, isolates were only positive for mecA (35.6%; 26/73), mecC/C1 (2.7%; 2/73), and tet(K) (14/73; 19%). Using pulsed-field gel electrophoresis (PFGE), nine clusters had identical PFGE patterns. Isolates produced inhibitory activity against a broad spectrum of bacteria; 20.5%, 19.2%, 17.8%, and 16.4% of S. xylosus were able to inhibit growth of Salmonella enterica serotype Typhi, methicillin-susceptible Staphylococcus aureus, Escherichia coli, and methicillin-resistant Staphylococcus aureus, respectively. This study demonstrated the presence of genetically related antimicrobial-resistant S. xylosus from eggs from household chickens. Like table eggs, eggs of household chickens also contain staphylococci that may be resistant to antimicrobials used to treat human infections. These data will allow comparison between staphylococci from eggs from different sources and may indicate the relative safety of eggs from household chickens. Further study of these egg types and their microbial composition is warranted.
Introduction
Chicken eggs are an important nutritionally rich food source for humans globally. Approximately 34 billion kilograms of in-shell chicken hen's eggs were produced collectively by the three largest egg-producing countries, China, the United States, and India, in 2016 (FAO, 2016). With the large production of eggs, it is important to monitor egg safety for the consumer. Eggs may be contaminated with pathogenic microorganisms such as Escherichia coli, Salmonella, Arcobacter, and Listeria monocytogenes (Lee et al., 2016) due to infected ovaries, and contact with contaminated trays, nests, cages, soil, or water (Svobodová and Tůmová, 2015). Although eggs possess several antibacterial proteins, the level of egg contamination is influenced by several factors such as microbial number in the environment and storage conditions (Scott and Silversides, 2000). The potential of biofilm formation on the egg shell by bacteria in the environment as well as fecal contamination of the egg shell during laying can cause the bacteria to become attached to the shell; contamination of the egg content may then occur when the shell is broken (Alvarez-Fernandez et al., 2012; Neira et al., 2017). Egg contamination with bacteria is a significant cause of poultry production pathology and disease spread among humans (Svobodová and Tůmová, 2015). The association of foodborne illnesses with consumption of contaminated eggs has already been established (Gormley et al., 2011; Osimani et al., 2016; Chousalkar et al., 2018).
Staphylococci are widespread in the environment and their presence in food samples, including eggs, is not unlikely as this genus of bacteria is prevalent in poultry and egg production settings (Papadopoulou et al., 1997; Brooks et al., 2010; Vela et al., 2012; Suwannarach et al., 2017). The most significant species of this group is Staphylococcus aureus, the causative agent of a range of illnesses among humans, including foodborne diseases, by production of a number of enterotoxins responsible for staphylococcal food poisoning (Le Loir et al., 2003; Hennekinne et al., 2012). Like other pathogenic bacterial species, S. aureus has also been detected from table eggs. The presence of S. aureus in the eggs not only poses a risk of diarrheal diseases among humans, but can also be transmitted to other humans through contaminated food (Pyzik and Marek, 2012). Other staphylococcal species such as S. cohnii, S. epidermidis, S. hominis, S. hyicus, S. lentus, S. sciuri, S. simulans, and S. xylosus have also been associated with eggs (Stepien-Pysniak et al., 2009; Pyzik and Marek, 2012; Suwannarach et al., 2017). The presence of these strains in eggs is of public health concern due to their abundance in the environment and potential medical impact on both human and animal health (Becker et al., 2014). These species may be of increased importance as, like S. aureus, they may also harbor acquired resistance to antimicrobials.
At present, most studies conducted on the microbial detection from eggs and risk assessment involved table eggs of farmed chickens (Stepien-Pysniak et al., 2009; Alvarez-Fernandez et al., 2012; Jones et al., 2012; Neira et al., 2017). In 2016, Pakistan produced ∼0.8 tons of table eggs (FAO, 2016); however, eggs of household chickens are widely consumed in rural areas of developing countries such as Pakistan and are considered more nutritious and healthier than eggs of farmed chickens. Such chickens are reared in villager's houses or backyards and their eggs are widely consumed by those living in rural areas. In contrast to farmed chickens, household chickens have a longer average life and seem to be free-living, spending most of their time on soil. Furthermore, these chickens experience more variation in environmental conditions and are fed on a diverse type of feed (Al-Nasser et al., 2007; Hannah et al., 2011). The ability of chicken eggs to resist bacterial entry is also hypothesized to be breed specific (Rathgeber et al., 2013).
As there is a paucity of information on the microbiology of eggs from household chickens, this study focused on staphylococci from eggs of these birds collected from different villages of Khyber Pakhtunkhwa province. Staphylococci were isolated, identified to species, and characterized for antimicrobial susceptibility, presence of resistance genes, genetic relatedness, and for inhibitory activity against other bacteria. To the best of our knowledge, this is the first study on bacterial detection from eggs of household chickens reared in personal homes.
Materials and Methods
Sample collection and processing
Household chicken eggs (n = 275) were collected from different villages of Khyber Pakhtunkhwa province of Pakistan from November 2016 to March 2017. The chickens were free range of different breeds reared by villagers in their backyards and all fed on organic feed. A maximum of two eggs were collected from each house to randomize the samples. Eggs were sealed in plastic bags and transported to the microbiology laboratory at the University of Haripur. The surface of each egg was disinfected by dipping in 70% ethanol for 5–10 s followed by air drying in a safety cabinet. Eggs were then broken open with a sterile rod and the contents were poured into separate sterile Petri plates. The inner membrane, egg white, and egg yolk were each tested for the presence of staphylococci. One milliliter of each egg white and egg yolk was 10 times diluted with phosphate-buffered saline (PBS, 1 × ; Oxoid, Basingstoke, UK) and homogenized up to 2 min. One hundred microliters of the homogenate was inoculated onto a mannitol salt agar plate (MSA; Oxoid) and incubated at 37°C for 24 h. The inner part of the egg shell was washed with 1 mL PBS and 100 μL of the mixture was inoculated onto MSA and also incubated at 37°C for 24 h.
Bacterial identification
Initial identification of the bacterial isolates was made using standard microbiological and biochemical tests, including growth on MSA, mannitol fermentation, microscopy, and catalase and coagulase tests. Identification to the genus level was also carried out by PCR using previously reported genus-specific primers for amplification of the 16S rRNA gene (Al-Talib et al., 2009). Further verification and identification of all strains to the species level was performed using the Vitek 2 system (bioMérieux, Durham, NC) and the Vitek 2 Gram-positive identification cards according to manufacturer's directions.
Antimicrobial susceptibility testing
Minimum inhibitory concentrations (MIC, μg/mL) for S. xylosus were determined by broth microdilution with the Sensititre™ semiautomated antimicrobial susceptibility system (Trek Diagnostic Systems, Inc., Cleveland, OH) and the Sensititre™ Gram-Positive Plate GPN3F according to the manufacturer's directions (Supplementary data). MIC values were manually recorded by Sensitouch® and Clinical and Laboratory Standards Institute (CLSI) standards were used to determine resistance (CLSI, 2018). As resistance breakpoints for daptomycin (range 0.25–8 μg/mL) and streptomycin (1000 μg/mL) have not been established by CLSI, breakpoints for those two antimicrobials were those defined by the National Antimicrobial Resistance Monitoring System-Enteric Bacteria (FDA, 2018). Staphylococcus aureus ATCC 29213 was used as a quality control strain.
Molecular analysis
Presence of resistance genes to macrolides [erm(A), erm(C)], aminoglycosides (aacA-aphD), oxacillin [mecA, mecC/C1], tetracycline [(tet(K), tet(M)], and streptogramins [vat(A), vat(B), vat(C)] was tested using multiplex PCR as previously described (Strommenger et al., 2003; Harrison et al., 2014). Pulsed-field gel electrophoresis (PFGE) was used to generate macrorestriction patterns using 30 U of SmaI (Roche, Indianapolis, IN) (McDougal et al., 2003). Cluster analysis was performed with BioNumerics software v6 (Applied Maths, Belgium) using Dice coefficient and the unweighted pair group method. Optimization settings for dendrograms were 2% with a band tolerance of 2%.
Inhibitory activity
All S. xylosus isolates were screened for inhibitory activity against four test bacteria, Salmonella enterica serotype Typhi, E. coli, methicillin-susceptible Staphylococcus aureus (MSSA, this study), and methicillin-resistant Staphylococcus aureus (MRSA) using the cross-streaking method as previously described (De Vliegher et al., 2004) (Supplementary data). The MRSA test strain was isolated from skin and soft tissue infections, whereas Escherichia coli and Salmonella enterica serotype Typhi were from urinary tract infection and typhoid cases, respectively.
Results
Bacterial identification
From the study, 23.6% (65/275) of eggs were positive for staphylococci. The majority of eggs were positive for S. xylosus (49/275; 17.8%), which was also the most prevalent species identified among the staphylococcal isolates (73/93; 78.5%) (Table 1). Other species identified were S. aureus (2/275; 0.73%), S. capitis (1/275; 0.36%), S. gallinarum (3/275; 1%), S. haemolyticus (2/275; 0.73%), S. lentus (3/275; 1%), and S. sciuri (2/275; 0.73%). Unidentified staphylococcal species accounted for 1% (3/275) of the isolates.
Prevalence of Staphylococci Among Household Chicken Eggs
Antimicrobial susceptibility and resistance genes of S. xylosus
Percent resistance of S. xylosus isolates to the tested antimicrobials is shown in Figure 1. The highest level of resistance (83.6%; 61/73 isolates) was to oxacillin. Isolates exhibited much lower levels of resistance to other β-lactam antibiotics, including ampicillin (13.7%; 10/73) and penicillin (12.3%; 9/73). Tetracycline resistance (24.7%; 18/73) was higher than ampicillin and penicillin, but much less so than oxacillin. Approximately 12.3% of isolates exhibited resistance to gatifloxacin, but <10% of isolates were resistant to the other fluoroquinolone antibiotics tested [ciprofloxacin (9.6%) and levofloxacin (6.8%)]. Some of the lowest resistance was observed to the macrolide-lincosamide-streptogramins (erythromycin, clindamycin, and Quinupristin/Dalfopristin) where only 4.1% of isolates were resistant for each antibiotic (Fig. 1). S. xylosus isolates exhibited low-level resistance (6.8%; 5/73) to the only aminoglycoside tested, gentamicin, as well as the newer antibiotics, daptomycin and linezolid (5.5% each). None of the S. xylosus strains was resistant to ceftriaxone, rifampicin, trimethoprim/sulfamethoxazole, or vancomycin.
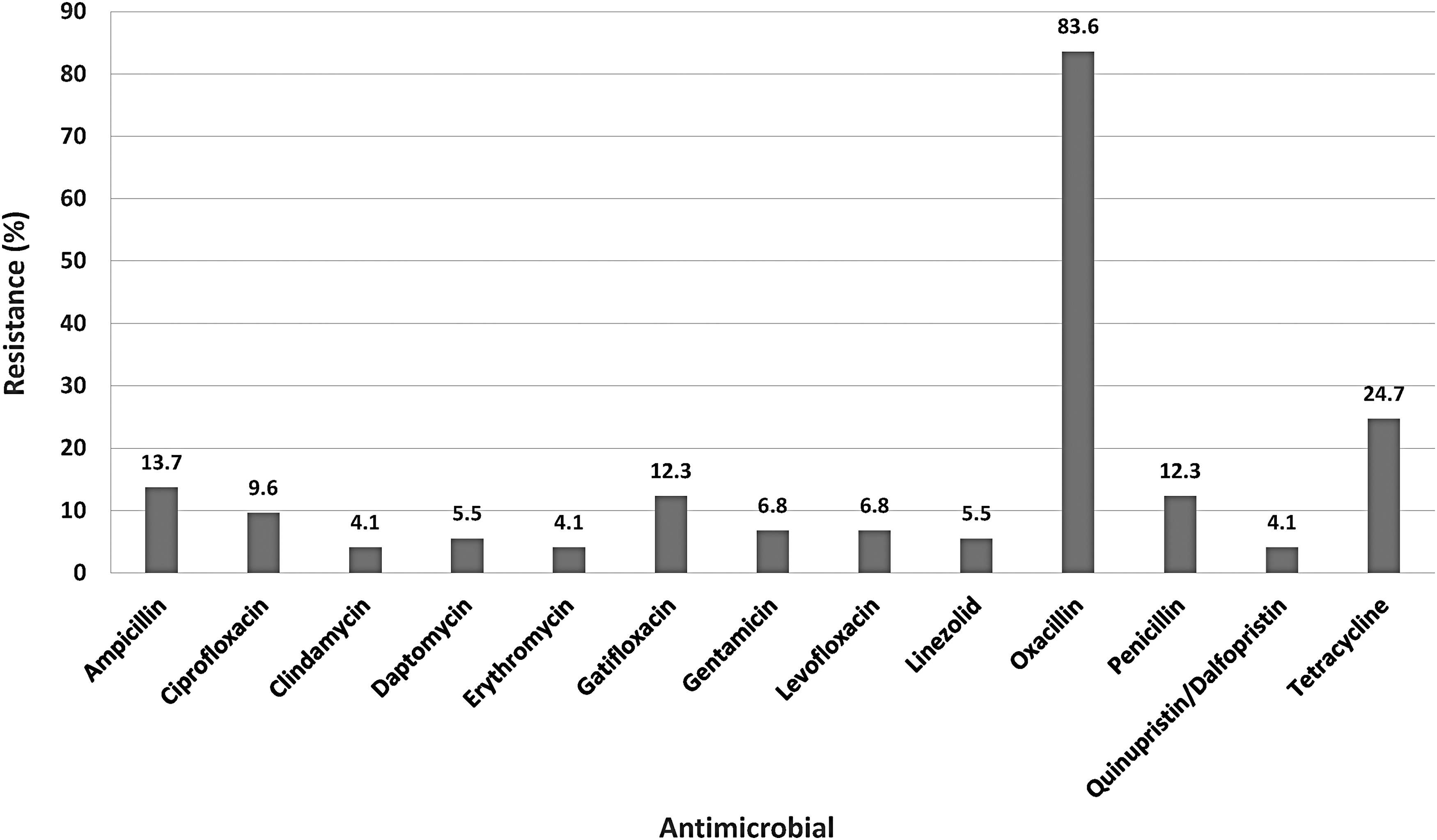
Percent resistance of Staphylococcus xylosus (n = 73) to tested antimicrobials using broth microdilution. No resistance to ceftriaxone, rifampicin, trimethoprim/sulfamethoxazole, or vancomycin was detected.
The majority of the resistance detected in the isolates was single resistance with only four isolates (36c, 128b, 205b, and 206c) exhibiting multidrug resistance (MDR; resistance to three or more antimicrobial classes) (Fig. 2). Tetracycline resistance was the only common resistance among those four isolates and all four were positive for the presence of tet(K), which was found in ∼19% (14/73) of isolates. The other resistance genes detected were mecA (35.6%; 26/73) and mecC/C1 (2.7%; 2/73) (Fig. 2, data not shown). Contrary to tetracycline resistance and the presence of tet(K), which were found together in over 92% of the tetracycline-resistant isolates, the presence of mecA or mecC/C1 did not always correspond with the oxacillin phenotype. Approximately 39% (24/61) of oxacillin-resistant isolates were negative for mecA/C/C1, while 50% (13/26) mecA/C-/C1-positive isolates were susceptible for oxacillin (Fig. 2, data not shown). Other resistance genes tested [erm(A), erm(C), (aacA-aphD), tet(M), vat(A), vat(B), and vat(C)] were not detected in any of the isolates.

Pulsed-field gel electrophoresis (PFGE) analysis, antimicrobial resistance pattern, and genetic profiles of Staphylococcus xylosus (n = 70) from household chicken eggs. DNA for PFGE was digested with SmaI. Levels of similarity were determined using Dice coefficient and the unweighted pair group method. Major clusters were based on ≥ 70% similarity and are labeled A and B. Black boxes represent positive results; gray boxes represent negative results. Antimicrobials are AMP, ampicillin; CIP, ciprofloxacin; CLI, clindamycin; DAP, daptomycin; ERY, erythromycin; GAT, gatifloxacin; GEN, gentamicin; LEV, levofloxacin; LIN, linezolid; OXA, oxacillin; PEN, penicillin; SYN, Quinupristin/Dalfopristin; TET, tetracycline.
Genetic relatedness using PFGE
PFGE analysis was performed on 70 S. xylosus; three isolates did not yield discernible patterns. Using PFGE, two major groups of isolates with at least 70% similarity were observed (Fig. 2, Supplementary Fig. S1). Group A was the smaller of the two groups and included 23 isolates that were more diverse (70% similar) than group B, which included 47 isolates with a minimum of 80% similarity. Within those groups, nine clusters contained isolates that were 100% identical (A1–A4 and B1–B5). Those isolates with identical PFGE patterns did not appear to cluster by gene or antimicrobial profile; however, group A contained all but one isolate harboring tet(K). Eleven of the 23 isolates (47.8%) in that cluster were also positive for mecA/C/C1 compared to 29.7% (14/47) of the isolates in group B. The majority of clusters were composed of S. xylosus from chicken eggs from two or more towns. For example, cluster B2 contained the largest number of identical S. xylosus (n = 20), mostly from the town of Mansehra, but also contained one isolate, 175a, from Haripur. Furthermore, identical isolates in this cluster originated from different villages within Mansehra and the village of KTS in the town of Haripur located over 60 km from Mansehra. Three clusters (B1, B3, and B4) containing isolates with identical PFGE patterns originated from eggs from three different towns and multiple villages. Each of three of the four MDR isolates (36c, 205b, and 206c) shared an identical PFGE pattern with at least one other isolate without an identical MDR phenotype.
Inhibitory activity
Overall, less than 21% of all tested S. xylosus (n = 73) exhibited phenotypic production of inhibitory activity against the four tested bacteria (Table 2). Approximately 20.5% (15/73) of isolates were able to inhibit growth of the Salmonella Typhi strain. Slightly less isolates, 19.2% (14/73), 17.8% (13/73), and 16.4% (12/73) were able to inhibit growth of the MSSA, E. coli, and MRSA, respectively. No isolate of S. xylosus was able to inhibit growth of all four bacteria, but five isolates, 131b, 160c, 162a, 189a, and 249b, had inhibitory activity against at least three target bacteria with varying patterns. Isolates 131b and 160c inhibited MSSA, MRSA, and Salmonella Typhi, while 162a inhibited MSSA, MRSA, and E. coli; isolates 189a and 249b were active against MRSA, Salmonella Typhi, and E. coli (Table 2). In addition to isolate 249b, five other S. xylosus, 185a, 211c, 228b, 4c, and 52c, also demonstrated activity against the two Gram-negative bacteria, Salmonella Typhi and E. coli.
Bacteriocin Production in Staphylococcus xylosus from Household Chicken Eggs
Forty isolates did not exhibit any inhibitory activity.
MRSA, methicillin-resistant Staphylococcus aureus; MSSA, methicillin-susceptible Staphylococcus aureus.
Discussion
This study revealed that, just like table eggs, eggs of the household chickens from the home setting also contain staphylococci (Stepien-Pysniak et al., 2009; Svobodová and Tůmová, 2015). The species detected from eggs of the household chicken were quite diverse, which has also been observed in studies on prevalence of staphylococci from various other types of eggs (Stepien-Pysniak et al., 2009; Pyzik and Marek, 2012). Our previous study on table eggs from Pakistan with comparable sample size revealed that there was a very low prevalence of S. aureus in those egg types (Syed et al., 2018). S. xylosus was by far the dominant species detected from household chicken eggs compared to S. aureus, S. hominis, or S. epidermidis from the previous studies, suggesting that the source of the egg may influence species prevalence.
S. xylosus has also been detected from poultry and the poultry environment (Nawaz et al., 1999; Aarestrup et al., 2000) and was the primary species detected in barn bioaerosols from broiler chicken poultry houses (Vela et al., 2012). Although considered a commensal bacterium and commonly found on skin of humans and animals (Nagase et al., 2002), S. xylosus has been implicated as an opportunistic pathogen in human infections, including those of the cornea (Pinna et al., 1999), root canal (Siqueira and Lima, 2002), bloodstream (Tompkins and Steele, 2004), urinary tract (Orrett and Shurland, 1998), and kidneys (Tselenis-Kotsowilis et al., 1982). Despite this association with clinical disease, S. xylosus are used in food as starter cultures for meat fermentation as this species has not been identified as a source of foodborne illness (Hammes and Hertel, 1998; Ordonez et al., 1999; Hennekinne et al., 2012). This versatility of S. xylosus strains was demonstrated using suppressive and subtractive hybridization, which showed that the genome of meat and dairy starter strains clustered separately from those strains that may have potential clinical relevance (Dordet-Frisoni et al., 2007).
Antimicrobial resistance in S. xylosus is not uncommon as resistant isolates have been isolated from poultry, the poultry environment, and food (Aarestrup et al., 2000; Resch et al., 2008; Vela et al., 2012). With the exception of oxacillin resistance, the lower level of antimicrobial resistance to the other antimicrobials tested in this study may be due to decreased use of antimicrobials in household chickens compared to the massive antibiotic use in poultry farms either as growth promoters or as therapeutic agents. However, other factors are necessary to account for the very high level of resistance to oxacillin and resistance to other drugs used only in human medicine, such as daptomycin and linezolid, which is reported for the first time in this study. As a close physical relationship between humans and the household chickens is present due to rearing practices for the birds, resistance in the household chicken eggs could be due to dissemination of antimicrobial-resistant bacteria or antimicrobial residues from the human host since neither those antimicrobials nor their analogs are used in poultry or egg production. Alternatively, resistance in the birds could also arise from antimicrobial resistance in the environmental habitat of the household chicken.
Resistance to the β-lactam antimicrobials was mediated by mecA or mecC/C1, while tetracycline resistance was conferred by tet(K). Both mecA and tet(K) have been commonly reported in S. xylosus (Aarestrup et al., 2000; Dordet-Frisoni et al., 2007; Resch et al., 2008; Vela et al., 2012); however, this study represents the first identification of mecC/C1 in S. xylosus to our knowledge. While other resistance genes, including blaZ, erm(A), and erm(C), have been found in S. xylosus from poultry (Nawaz et al., 1999; Vela et al., 2012), none of the isolates in this study was positive for those genes. Some discrepancies were observed between antimicrobial phenotype and genotype. For example, although tet(K) is considered ubiquitous in tetracycline-resistant staphylococci (Levy et al., 1989), not all tetracycline-resistant isolates were positive for that gene or other tetracycline resistance genes tested in this study, suggesting that other tetracycline resistance genes may be present. Conversely, some isolates harbored tet(K), but were not phenotypically resistant to tetracycline, possibly due to an inactive or defective gene. These discrepancies between phenotype and genotype were even more pronounced for oxacillin resistance. However, according to CLSI (CLSI, 2018), the breakpoints for oxacillin may falsely predict resistance for some coagulase-negative staphylococci. In the case of some S. epidermidis strains, the strains are phenotypically resistant (oxacillin MICs are 0.5–2 μg/mL), but are negative for mecA.
Bacterial diversity in eggs of household chickens is expected as they live in diverse habitats and environmental conditions; however, high genetic diversity was not observed among the S. xylosus in this study. The ability to roam and mixing of birds and their flora may have instead contributed to the high percentage of similarity and very little diversity in their genetic makeup using PFGE. These data vary greatly from that of Vela et al. (2012), in which only 15% similarity was seen among the two main groups of S. xylosus from poultry bioaerosols, even though those isolates were collected from a discrete group of birds. The analysis used may also have accounted for the differences as PFGE is a standardized method and considered a gold standard for typing staphylococci, while RAPD profiling results can vary from laboratory to laboratory and even within laboratories (Foley et al., 2007).
In addition to their nutritional value, eggs possess several proteins exhibiting antibacterial effects (Eckert et al., 2013). Due to the differences in dominant staphylococcal species between household chicken eggs compared to table eggs, it was possible that S. xylosus in household chicken eggs inhibited the S. aureus. This was confirmed using a cross-streaking method in which S. xylosus inhibited the growth of MSSA, MRSA, Salmonella, and E. coli strains, suggesting the presence of bacteriocins, peptides produced by bacteria that are capable of inhibiting the growth of closely related bacteria (Cotter et al., 2005). Although not all S. xylosus were able to inhibit all bacterial targets tested, the inhibition could be described as broad spectrum as both the closely related Gram-positive bacteria, MSSA and MRSA, were inhibited as well as the Gram-negative bacteria, Salmonella and E. coli. Additional study is needed to confirm the presence of bacteriocins and the antibacterial properties of S. xylosus from the eggs.
Footnotes
Acknowledgments
The authors are grateful to the U.S. Department of Agriculture-Agricultural Research Service and the University of Haripur for financial support for this research.
Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Data
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.