
Abstract
Vibrio parahaemolyticus is a halophilic Gram-negative foodborne pathogen that is widely distributed in marine environments. It can cause acute gastroenteritis and other diseases. This study aimed to investigate the antivirulence activity of thymoquinone (TQ) on V. parahaemolyticus. TQ was shown to effectively inhibit V. parahaemolyticus. Subminimum inhibitory concentrations of TQ inhibited swimming and swarming motility, quorum sensing, biofilm formation, the ability of V. parahaemolyticus to adhere and invade the host cells, and the expression of virulence-associated genes of V. parahaemolyticus. These findings suggest that TQ can effectively inhibit the growth of V. parahaemolyticus and significantly reduce its pathogenicity. Considering its safety and various biological activities, TQ has the potential to be developed as a natural antibacterial substance to reduce the diseases associated with V. parahaemolyticus.
Introduction
V ibrio parahaemolyticus is a Gram-negative, halophilic bacterium which is widely distributed in estuarine and marine environments (Zhang et al., 2017). It can be isolated from a variety of seafoods, including shrimp, oyster, crab, clam, scallop, and marine fish (Su and Liu, 2007). Consumption of seafood contaminated with pathogenic V. parahaemolyticus may cause abdominal cramps, diarrhea, vomiting, fever, and headache and lead the gastroenteritis patients to death (Drake et al., 2007; Vengadesh et al., 2014). In Korea, 9–16% of food-poisoning cases were associated with V. parahaemolyticus (Han et al., 2016), and in the United States and China, V. parahaemolyticus is recognized as the leading cause of bacteria-derived food poisoning (Iwamoto et al., 2010; Wu et al., 2014).
Biofilm formation can protect bacteria from various environmental stresses and has been linked to bacterial survival and virulence (Packiavathy et al., 2013). V. parahaemolyticus can form biofilms on biotic and abiotic surfaces, including oysters, crab, shrimp, and stainless steel, and the ability of biofilm formation is thought to be vital for the physiological function of V. parahaemolyticus (Han et al., 2016; Mizan et al., 2016). Chao et al. (2010) examined 71 foodborne and 116 clinical V. parahaemolyticus isolates and the results showed that most of the strains possessed the genes encoding biofilm formation factors. The formation of biofilm by V. parahaemolyticus was found to be regulated by the quorum-sensing (QS) system (Packiavathy et al., 2013), and autoinducer-2 (AI-2)-based interspecies QS controls virulence in several Vibrio species (Brackman et al., 2009).
Antibiotic treatment is a common method to prevent and control the infections caused by pathogens (Silva et al., 2016). Bacteria presenting resistance to conventional antibiotics become more common today, and some antibiotics are no longer effective against various pathogens (Levy and Marshall, 2004). Plant materials have received a great deal of interest regarding as alternative methods to control pathogenic microorganisms. Several researchers have presented the promising effects of natural antibacterial substances on inhibiting infection (Silva et al., 2016).
Thymoquinone (TQ, C10H12O2) is the major bioactive constituent of the volatile oil from black seed (Nigella sativa) (Galimuhtasib et al., 2004), which has been commonly used for medical purposes in the Middle East (Chaieb et al., 2011), and it has been proved that most of the biological activities derive from TQ. Previous studies have demonstrated that TQ has a variety of beneficial properties, such as antioxidative, anti-inflammatory, and antitumor activities (Worthen et al., 1998; Ragheb et al., 2009). In addition, the antibacterial and antifungal activities of TQ have also been reported, the minimum inhibitory concentrations (MICs) of TQ against Staphylococcus aureus, Streptococcus mutans, Listeria monocytogenes, and Bacillus cereus were 8–64 mg/L, and the MICs of TQ against Dermatophyte were in range of 125–250 mg/L (Kouidhi et al., 2011; Forouzanfar et al., 2015).
Although the antimicrobial effects of TQ on microorganisms have been reported extensively (Forouzanfar et al., 2015), its antivirulence effects on V. parahaemolyticus in vitro have rarely been investigated. The aim of this study was to determine the effects of TQ against the virulence factors of V. parahaemolyticus.
Materials and Methods
Reagents
TQ (CAS: 490-91-5, 98% purity) was purchased from Tokyo Chemical Industry Co. Ltd. (Tokyo, Japan). The TQ solutions were prepared in 0.1% dimethyl sulfoxide (DMSO) before use, as described in a previous study (Shi et al., 2015).
Bacterial strains and culture conditions
V. parahaemolyticus ATCC 17802 and ATCC 33847 and Vibrio harveyi BB170 and BB120 were obtained from the American Type Culture Collection (Manassas, VA). Four other V. parahaemolyticus strains were isolated from seafood by the Center for Food Safety and Technology Research of Hong Kong Polytechnic University (Shenzhen, China). All of the V. parahaemolyticus isolates were used in MIC assay, and only ATCC 17802 was used for further experiments, because ATCC 17802 is commonly used in V. parahaemolyticus virulence studies as the type strain, and it contains the phenotypic and genotypic characteristics (including motility, biofilm formation ability, AI-2 production, and adhesive and invasive abilities) tested in the previous experiments (Tarah et al., 2005; Packiavathy et al., 2013).
All V. parahaemolyticus strains were stored in tryptone soya broth (TSB) with 3% NaCl (w/v) and 30% glycerol (v/v) at −80°C. Before each experiment, V. parahaemolyticus was activated on tryptone soya agar (TSA) with 3% NaCl at 37°C for 18 h. Then a loopful of each strain was inoculated into 30 mL of TSB (3% NaCl) and incubated for 12 h at 37°C (100 rpm). V. harveyi BB170 and BB120 were activated in autoinducer bioassay (AB) agar at 30°C for 24 h. Then, the microorganisms were seeded into AB broth with shaking (180 rpm) at 30°C for 16 h before use.
Determination of the MICs and sub-MICs
The MICs of TQ against V. parahaemolyticus were determined by the agar dilution method as Shi et al. (2016b). In brief, TQ was dissolved in DMSO and incorporated into TSA (3% NaCl) to obtain final concentrations of 100, 75, 50, 37.5, 25, 18.75, 12.5, 9.375, and 0 mg/L, respectively. TSA (3% NaCl) with 0.1 mg/mL kanamycin was used as a positive control. The agar media was seeded with 2 μL (∼104 colony-forming unit [CFU]) of the test bacterium and incubated at 37°C for 24 h. The lowest concentration of TQ that results no visible growth of test organisms was determined as MIC.
The sub-MICs of TQ for V. parahaemolyticus ATCC 17802 were determined using the growth curve method as Shi et al. (2017a). The final concentrations of TQ were 50, 25, 12.5, 6.3, 3.2, 1.6, 0.8, 0.4, 0.2, and 0 mg/L. The bacteria were further cultured at 37°C for 24 h, and bacteria growth was determined by measuring the OD600nm (optical density) at 1-h intervals. The three highest concentrations of TQ that did not inhibit bacterial growth were selected as sub-MICs for further tests.
In this study, sub-MICs (the concentrations which did not inhibit bacterial growth) were used to ensure the quantities of V. parahaemolyticus treated with or without the sub-MICs of TQ were almost the same. It means that the results in this study were not affected by the effect of TQ on bacterial quantity.
Motility assay
Swimming and swarming motility assays were performed as described by Li et al. (2014). For the swimming assay, 5 μL of V. parahaemolyticus (∼104 CFU) was seeded onto the surface of semisolid medium (20 mL of LB broth, 0.3% agar, and 3% NaCl) in the absence (control) or presence of TQ (0.8, 1.6, and 3.2 mg/L). The swimming plates were incubated upright at 37°C for 7 h. For the swarming assay, 5 μL of V. parahaemolyticus was seeded onto the surface of the semisolid medium (20 mL of LB broth, 0.5% agar, 0.5% glucose, and 3% NaCl) with or without TQ. The swarming plates were incubated upright at 37°C for 18 h. After incubation, the diameters of the swimming and swarming areas were recorded and analyzed using the ImageJ software (National Institutes of Health).
AI-2 quantification
V. harveyi BB170 (which produces light in response to AI-2) and BB120 (which produces AI-1 and AI-2) were used in this test. The effect of TQ on the growth of V. harveyi BB170 was determined by the growth curve method as described by Shi et al. (2017a). The AI-2 determination assay was performed according to Han et al. (2016) with minor modifications. In brief, V. harveyi BB170 was grown overnight at 30°C in AB broth and then diluted 1:1000 with AB broth. V. parahaemolyticus ATCC 17802 was incubated with or without sub-MICs of TQ for 6 h with shaking (180 rpm) at 30°C. Then, the bacterial suspensions were centrifuged at 8000 × g for 5 min, and the supernatants were then passed through 0.22 μm Tuffryn syringe filters. Next, 500 μL of the cell-free supernatants from each sample was added into 4.5 mL of the suspension of V. harveyi BB170, and then cultured at 30°C with shaking (180 rpm) for 16 h. Subsequently, 200 μL samples were transferred into a white microtiter plate and the bioluminescence intensity was measured by a microplate reader (Infinite™ M200 PRO; Tecan, Männedorf, Switzerland). The cell-free supernatants of V. harveyi BB120 were used as positive control (Han et al., 2016).
Biofilm formation inhibition assay
Specific biofilm formation inhibition assay
Biofilm formation determination was based on the microtiter plate test developed by Naves et al. (2008) with some modifications. In brief, 250 μL of the test organism suspension (OD600nm = 1) was cultured in 96-well microtiter plates with different concentrations of TQ (0, 0.8, 1.6, and 3.2 mg/L). The TSB (3% NaCl) without bacteria and TQ was used as the background. The samples were cultured at 30°C for 24, 48, and 72 h. After incubation, the OD630nm of each sample was measured. The content of each well was removed and the wells were washed with 350 μL of distilled water. The plates were air-dried for 30 min and stained with 250 μL of 0.1% (w/v) crystal violet for 20 min at room temperature. The unbound colorant was discarded, and the wells were washed three times with 350 μL of distilled water. Finally, the crystal violet was solubilized by 250 μL of 33% (v/v) glacial acetic acid and incubated for 20 min at 25°C. The absorbance was read at 570 nm using a microtiter spectrophotometer (Model 680; Bio-Rad Laboratories, Hercules, CA), and the specific biofilm formation (SBF) was calculated by correcting the OD570nm with the OD630nm. The value of OD570nm expresses the amount of biofilm formed, and the number of cells grown in suspended culture was represented by the value of OD630nm (Niu and Gilbert, 2004).
Field emission scanning electron microscopy
The field emission scanning electron microscopy (FESEM) assay was carried out according to Shi et al. (2016a) with minor modifications. The microorganisms treated with or without TQ were seeded into 24-well plate with glass slides incubated at 30°C for 72 h. After incubation, the glass slides were removed from the wells and washed with sterile water. The glass slides were then fixed with phosphate buffer saline (PBS, pH = 7.2) containing 2.5% glutaraldehyde, and then kept at 4°C for 12 h. Subsequently, the slides were washed with PBS gently and then dehydrated gradually in a graded water-ethanol series (30%, 50%, 70%, 80%, 90%, and 100%) for 10 min each. Finally, the samples were dried and gold sputtered, and then examined under a field-emission scanning electron microscope (S-4800; Hitachi, Tokyo, Japan).
Adhesion and invasion assays
Human enterocyte-like Caco-2 cells were obtained from the American Type Culture Collection, the cells were maintained as Shi et al. (2017a). For adhesion assay, Caco-2 cells were seeded at 105 cells/mL into 24-well tissue culture plates containing Dulbecco's modified Eagle's medium (DMEM) with 10% fetal bovine serum (FBS) and cultured at 37°C with 5% CO2 for 18 h. V. parahaemolyticus strains were treated with or without the sub-MICs of TQ for 6 h at 37°C in a shaker (100 rpm). The bacteria were then washed and resuspended by fresh DMEM. Before bacterial infection, the Caco-2 cells were washed with PBS twice, and then the bacteria were applied to Caco-2 cells at an Multiplicity of infection = 10 for 2 h at 37°C. The Caco-2 cells were washed with PBS three times after incubation. Subsequently, the monolayers were lysed with 0.1% Triton X-100 for 20 min at 4°C. Then, all of the liquid was collected, and serial dilutions of the bacterial suspension were plated onto TSA (3% NaCl) and incubated at 37°C overnight.
For invasion assay, after being treated with V. parahaemolyticus, Caco-2 cells were washed with PBS. DMEM containing 100 μg/mL kanamycin was added and incubated for 45 min at 37°C in 5% CO2 to kill the extracellular bacteria. The subsequent steps were the same as the adhesion assay. Viable bacteria were counted and the results were presented as a percentage of the V. parahaemolyticus of control group (V. parahaemolyticus without the treatment of TQ).
RNA isolation and quantitative real-time polymerase chain reaction
V. parahaemolyticus ATCC 17802 was treated with or without sub-MICs of TQ at 37°C for 6 h with shaking (100 rpm). After incubation, the total RNA was isolated from V. parahaemolyticus using the RNAprep Pure Bacteria Kit (Tiangen, Beijing, China) according to the manufacturer's instructions. The integrity and concentrations of the RNA were measured using a nucleic acid and protein spectrophotometer (Nano-200; Aosheng Instrument Co., Ltd., Hangzhou, China). The RNA was then immediately reverse transcribed to complementary DNA (cDNA) using the Takara PrimeScript RT Reagent Kit (Takara, Kyoto, Japan). Real-time polymerase chain reaction (PCR) was done with the IQ5 system (Bio-Rad Laboratories) by using SYBR® Premix Ex Taq™ II kit (Takara). PCR reactions were performed in a 25 μL system containing 12.5 μL of 2 × SYBR Premix Ex Taq II (Tli RNaseH Plus), 1 μL of 10 μmol/L PCR forward primer, 1 μL of 10 μmol/L PCR reverse primer, 2 μL of cDNA, and 8.5 μL of distilled water. Thermal cycling conditions for the quantitative PCR (qPCR) were as follows: 1 cycle of 95°C for 30 s, 40 cycles of 95°C for 5 s, and 55°C for 30 s. All samples were analyzed in triplicate. The puvA gene was used to normalize the gene expression levels among samples. The sequences of the primers used in the qPCR analysis are listed in Table 1. The expression of target genes versus puvA gene were determined by the 2−ΔΔCT method described previously (Li et al., 2014).
Primers and Polymerase Chain Reaction Programs for the Detection of Target Genes
p ≤ 0.01, b p ≤ 0.05 compared with the control.
F, forward; R, reverse.
Statistical analysis
All data are presented as the mean values ± standard deviation, and differences between means were tested by Student's t-test. Statistics were analyzed using IBM SPSS v.19.0 software (version 19.0; SPSS, Inc., IBM Co., Armonk, NY). p ≤ 0.05 was considered significant. All experiments were performed at least in triplicate.
Results
The MICs and sub-MICs of TQ against V. parahaemolyticus
TQ showed a good antimicrobial effect on the tested V. parahaemolyticus strains. The MICs of TQ against six V. parahaemolyticus were 12.5–25 mg/L (Table 2). V. parahaemolyticus ATCC 17802 was chosen for further studies.
Minimum Inhibitory Concentrations of Thymoquinone Against Different Vibrio parahaemolyticus Strains
MIC, minimum inhibitory concentration.
The effect of TQ on the growth of V. parahaemolyticus ATCC 17802 is shown in Figure 1. TQ at 0.2–3.2 mg/L had no significant effect on the growth of V. parahaemolyticus, and the three highest concentrations of TQ (0.8, 1.6 and 3.2 mg/L) that did not inhibit V. parahaemolyticus were selected as sub-MICs for the following experiments.
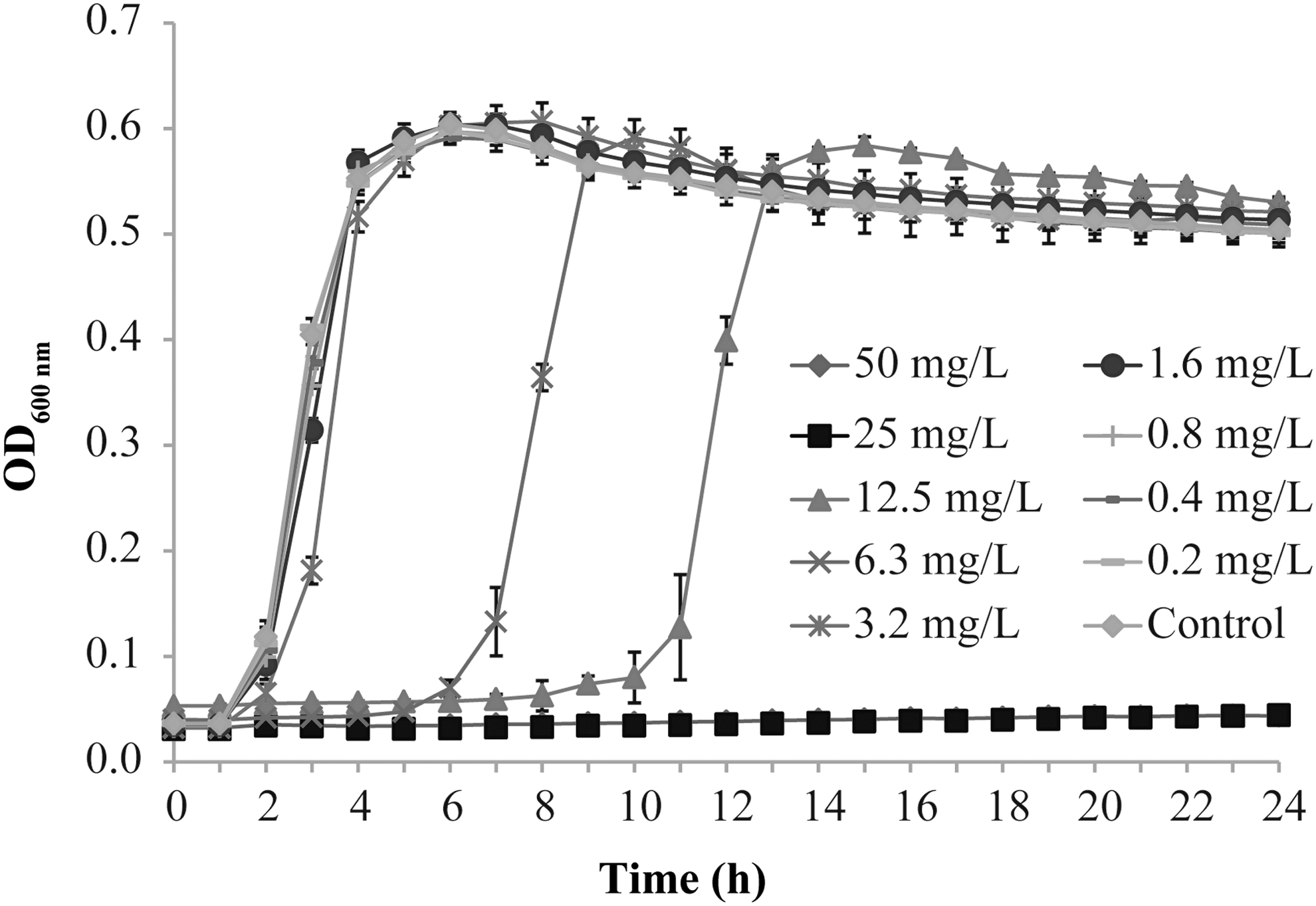
Effect of TQ on the growth curve of Vibrio parahaemolyticus ATCC 17802. Bars represent the SD. Each value represents the average of six independent measurements. SD, standard deviation; TQ, thymoquinone.
Swimming and swarming inhibition assay
As shown in Figure 2, TQ reduced the swimming and swarming motility of V. parahaemolyticus ATCC 17802. In the swimming assay, the swimming areas of bacteria treated with 1.6 and 3.2 mg/L of TQ were 39.13% and 25.06% that of the control, respectively (Fig. 2A). The swarming area of the control was 1.93 ± 0.20 cm2. The swarming areas were reduced to 0.70 ± 0.13 cm2 and 0.70 ± 0.07 cm2 by TQ at 1.6 and 3.2 mg/L, respectively (Fig. 2B).

Effect of TQ on the motility of V. parahaemolyticus ATCC 17802.
AI-2 quantification
When the cell-free supernatants of V. parahaemolyticus were added into the suspension of V. harveyi BB170, the concentration of TQ was diluted to 10% of original concentration. As shown in Figure 3A, TQ at 0.16 and 0.32 mg/L has no apparent antimicrobial activity against V. harveyi BB170. According to Figure 3B, the bioluminescence intensity of V. parahaemolyticus was significantly decreased to 74.15% and 67.99% that of the control by TQ at 1.6 and 3.2 mg/L, respectively.
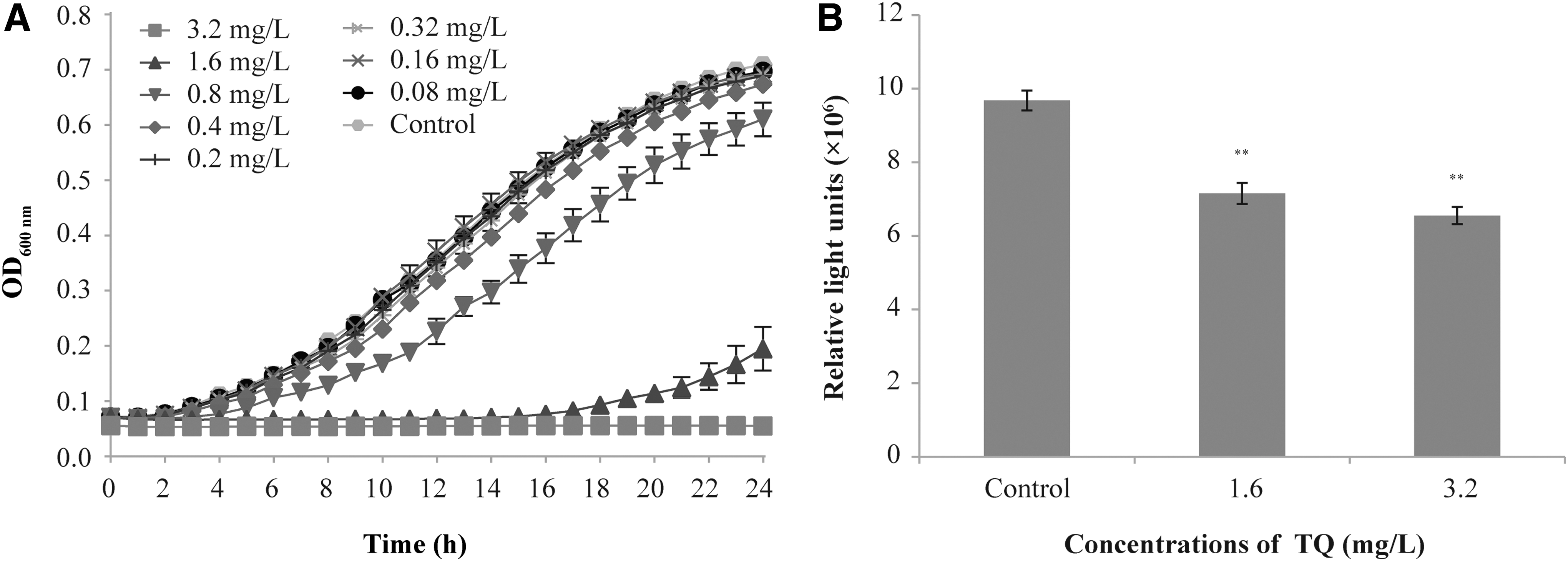
Biofilm formation
TQ showed a dose-dependent inhibition effect on the biofilm formation of V. parahaemolyticus. Compared to the control, SBF was decreased to 64.10%, 58.40%, and 47.44% by TQ at 0.8, 1.6, and 3.2 mg/L after treatment at 30°C for 72 h, respectively (Fig. 4).

Effect of TQ on biofilm formation by V. parahaemolyticus ATCC 17802 at 30°C. Mean values of triplicate independent experiments and SD are shown. *p ≤ 0.05, **p ≤ 0.01 compared with the control.
FESEM was used to assess the antibiofilm potential of TQ against biofilm morphological structures (Fig. 5). The untreated V. parahaemolyticus showed a dense biofilm layer and extracellular matrix on the surface of the glass slides after incubation for 72 h (Fig. 5A). By contrast, V. parahaemolyticus treated with sub-MICs of TQ at 30°C for 72 h showed lower biofilm content in comparison with the control and individual cells were loosely attached to the surface (Fig. 5B–D).
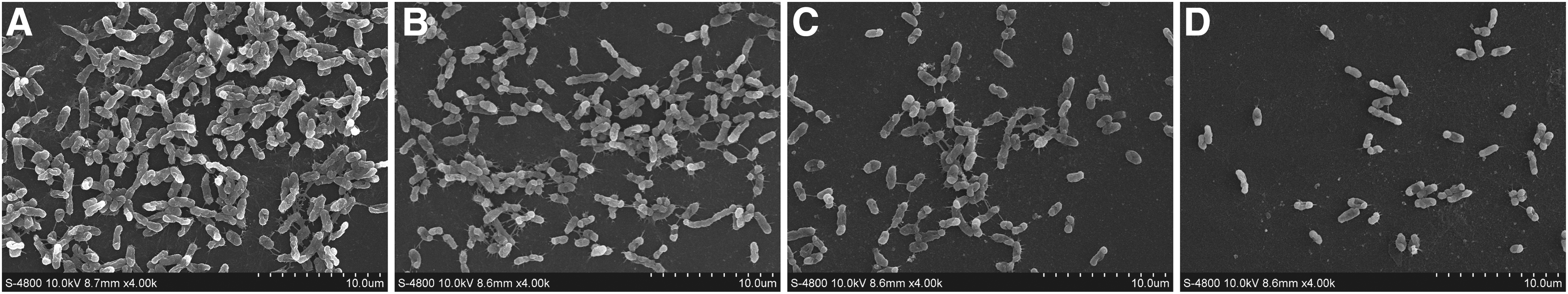
Field emission scanning electron micrographs (4000 × magnification) of biofilm formed by V. parahaemolyticus ATCC 17802 either untreated
Adhesion and invasion
The effect of TQ on the ability of V. parahaemolyticus to adhere and invade Caco-2 cells is shown in Figure 6. The numbers of adhesive bacteria were decreased to 65%, 54%, and 19% of the control by TQ at 0.8, 1.6, and 3.2 mg/L, respectively (Fig. 6A). TQ at sub-MICs also showed the inhibitory effect on the invasive ability of V. parahaemolyticus (Fig. 6B). TQ at 0.8, 1.6, and 3.2 mg/L led to a 14%, 35%, and 41% reduction of the invasive V. parahaemolyticus, respectively.
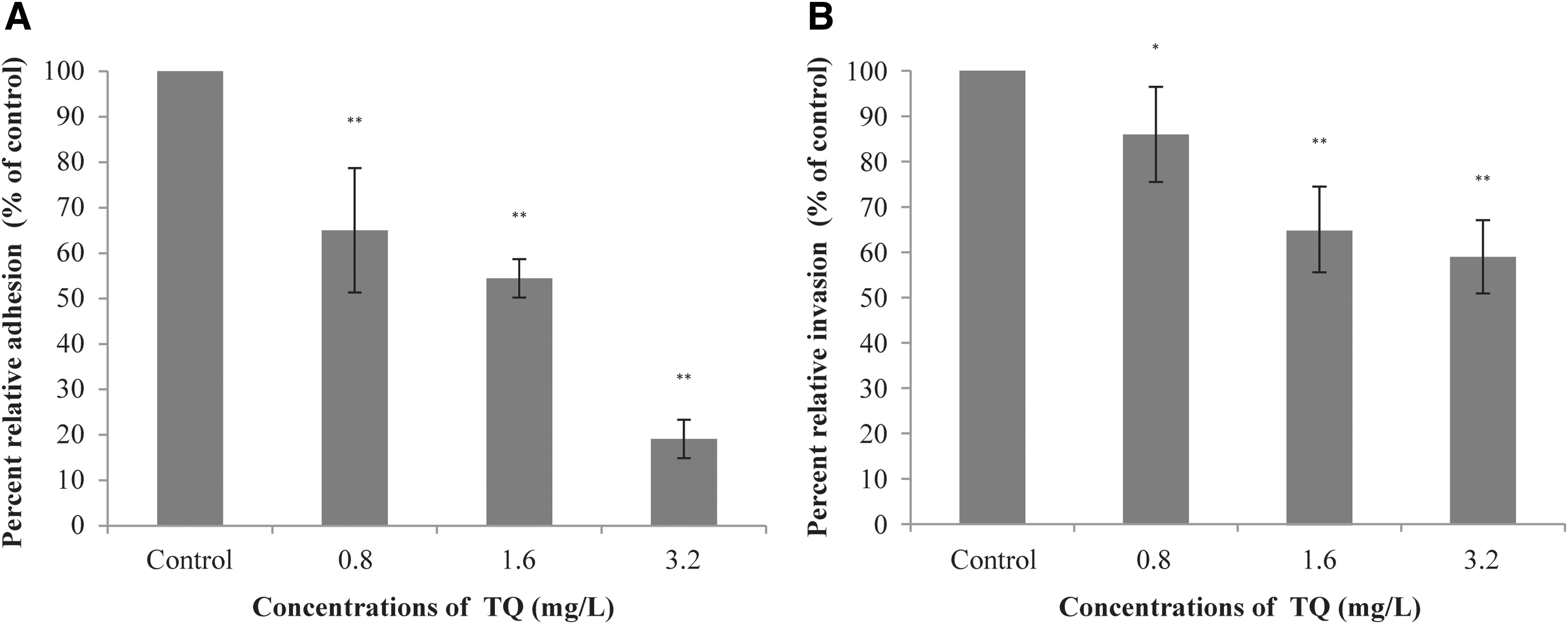
Effect of TQ on the ability of V. parahaemolyticus ATCC 17802 to adhere
Real-time quantitative polymerase chain reaction (RT-qPCR)
The expressions of flaA, flaL, flaM (critical for flagella regulation), tolc, fla, ompW (membrane protein-related gene), aphA, luxS (QS-related gene), and toxR (transmembrane transcriptional activator gene) were inhibited to various degrees by TQ at sub-MICs. TQ also downregulated the expressions of three genes (VP0950, VP0952, and VP0962) associated with biofilm formation.
Discussion
Bacterial motility is closely associated with virulence and affects the colonization and invasion ability of bacteria toward host cells (Haiko and Westerlundwikström, 2013). In this study, TQ was shown to inhibit the motility of V. parahaemolyticus both in terms of swimming and swarming. Our findings were consistent with those of a previous study (Packiavathy et al., 2013), in which the curcumin exhibited a significant inhibitory effect on the motility of Vibrio spp. Similarly, Shi et al. (2017a) showed that citral inhibited the motility of Cronobacter sakazakii and decreased the expression of genes associated with flagella structure and biosynthesis. In the current study, RT-qPCR results showed that TQ downregulated the expression of genes associated with the synthesis of flagella in V. parahaemolyticus (flaA, flaM, and flaL). Therefore, it is hypothesized that TQ inhibits the motility of V. parahaemolyticus by modulating the synthesis of the flagellum.
Bacterial biofilms can endanger the public health by helping pathogens to resist antibiotic and environmental stress (Silva et al., 2016). Our results showed that sub-MICs of TQ can significantly decrease biofilm formation of V. parahaemolyticus at 30°C and affect the morphological structures of the biofilm. This result was consistent with that of a previous study (Xie et al., 2017), which reported that a sub-MIC of chitosan was effective in preventing biofilm formation. Similarly, Santhakumari et al. (2018) demonstrated the antibiofilm activity of 2,6-di-tert-butyl-4-methylphenol on V. parahaemolyticus isolates, as shown by analyzing the biomass and architecture of strains through a light microscope and scanning electron microscope. Shi et al. (2017b) reported that TQ significantly inhibited the biofilm formation of C. sakazakii by reducing cellulose and flagella production. The genes selected for this study were VP0950, VP0952, and VP0962, which have been found to be associated with biofilm formation of V. parahaemolyticus (Chao et al., 2010), and RT-qPCR results demonstrated that the expressions of these genes were downregulated by TQ at sub-MICs. However, the effect of TQ on biofilm formation at other temperatures, such as seawater (e.g., 26°C) and common storage refrigerated temperatures (4–10°C), was not determined in this study. More research is needed to study the temperature-dependent effect.
The QS system mediates communication among bacteria by secreting extracellular signaling molecules called AIs, and this system can regulate biofilm formation, bacterial motility, and the expression of virulence factors (Mizan et al., 2016). V. harveyi BB170 is a mutant strain that is capable of sensing AI-2 but not AI-1 (Niu et al., 2006). In this study, TQ was shown to significantly inhibit the production of the QS signal molecule AI-2 by measuring the bioluminescence intensity of V. harveyi BB170. Similarly, Niu et al. (2006) reported that cinnamaldehyde had the capacity to inhibit QS as observed by the reduction of the bioluminescence intensities of AI-2 bioreporter strain. In the current study, RT-qPCR demonstrated that luxS, which encodes the key enzyme in AI-2 synthesis, was downregulated by TQ at sub-MICs. AphA is a master QS regulator operating at low cell density of vibrios (Sun et al., 2012). In this study, the cell densities of V. parahaemolyticus treated with or without the sub-MICs of TQ were the same, but the transcription of aphA was significantly downregulated (p ≤ 0.01) by TQ at sub-MICs.
Adherence to host tissues is a prerequisite for a microbe to establish infection since it avoids the organism from being swept away by the natural cleansing mechanisms of the host (Sharon and Ofek, 2000). In this study, we confirmed that TQ significantly suppressed the ability of V. parahaemolyticus to adhere and invade Caco-2 cells. This was consistent with the findings of a previous study (Xu et al., 2015), in which the tannin-rich fraction from pomegranate rind was shown to significantly repressed the adhesion and invasion ability of L. monocytogenes. Similarly, Li et al. (2014) demonstrated that punicalagin reduced the invasive Salmonella Typhimurium in HT29 cells and downregulated the expression of several virulence genes of Salmonella Typhimurium required for pathogen colonization. Furthermore, it has been reported that carvacrol, thymol, and eugenol can significantly reduce the adhesive ability of Salmonella Enteritidis to chicken oviduct epithelial cells (Upadhyaya et al., 2013).
Conclusion
This study demonstrated that TQ at sub-MICs can inhibit swimming and swarming motility, AI-2-based QS system, biofilm formation, adhesive and invasive capacities to Caco-2 cells and the expression of virulence-associated genes of V. parahaemolyticus. These findings suggest that TQ is a promising antivirulence therapeutic agent to prevent and reduce the risk of diseases associated with V. parahaemolyticus. However, before its practical application, the antivirulence effects of TQ in vivo need to be confirmed.
Footnotes
Acknowledgments
This work was supported by the Fundamental Research Funds for the Central Universities (2452017228), National Natural Science Foundation of China (31801659), and General Financial Grant from the China Postdoctoral Science Foundation (No. 2017M623256).
Disclosure Statement
The authors declare that there are no conflicts of interest.