
Abstract
Psoralea corylifolia seeds contain many bioactive compounds commonly used in traditional Chinese medicine. In this study, the antibacterial activity and possible mechanism of P. corylifolia seed ethanol extract (PCEE) against foodborne pathogens were investigated. Both methicillin-resistant Staphylococcus aureus (MRSA) and Listeria monocytogenes had similar minimum inhibitory concentrations and minimum bactericidal concentrations of PCEE at 50 and 100 μg/mL, respectively. Furthermore, elevated OD260, protein concentration, and electric conductivity indicated irreversible damage to the cytoplasmic membranes of PCEE-treated cells. Indeed, the treated cells displayed disrupted membranes, incomplete and deformed shapes, and rupture as visualized by scanning electron microscopy. Multidrug-resistance efflux pump gene expression was also analyzed by quantitative reverse transcription PCR. Although the mdrL, mdrT, and lde genes of L. monocytogenes and the mepA gene of MRSA were upregulated, there was no significant difference that indicated an attempt by the efflux pumps to discharge PCEE. MRSA norA expression and abcA expression were significantly downregulated (p < 0.05). A possible mechanism for PCEE may be to cause an energy depletion, either by inhibiting adenosine triphosphate binding or by disturbing the proton gradient, resulting in membrane damage.
Introduction
Microorganisms have continuously invaded humans, resulting in a high incidence of disease and high mortality. With the development of antibacterial agents in the mid-20th century, infections have been effectively controlled. In vitro, however, abuse of these antibacterial agents causes bacteria to develop resistances to many antibacterial agents, such as penicillin-resistant Staphylococcus aureus, tetracycline-resistant Listeria monocytogenes, and multidrug-resistant Escherichia coli and Salmonella (Levy, 2001; Lungu et al., 2011). When drug resistance is acquired, bacteria are less sensitive to antibiotics, resulting in failure of treatment, and becoming detrimental to medical care (McGowan, 2001).
L. monocytogenes is a foodborne pathogen that causes listeriosis in pregnant women, newborns, and immunocompromised individuals. Common symptoms include miscarriage, neonatal death, sepsis, and meningitis, with a mortality rate of ∼15.9% (Scallan et al., 2011). S. aureus is the causative agent of a variety of diseases, skin and soft tissue infections, life-threatening sepsis, endocarditis, and toxic shock syndrome (Diep et al., 2006). Methicillin-resistant S. aureus (MRSA) is the most common bacterium in nosocomial infections, which is resistant to various classes of antibiotics, resulting in many therapeutic limitations (Enright et al., 2002).
Psoralea corylifolia is a popular medicinal plant found in tropical and subtropical regions, particularly India, China, and South Africa (Krishnamurthi et al., 1969). The plant and its seeds are widely used in Ayurveda and traditional Chinese medicine to treat white spot, leprosy, psoriasis baldness, and dermatitis (Gidwani et al., 2010), as well as for their therapeutic effect on kidney and spleen meridians (Lui, 2000). P. corylifolia seeds contain a higher concentration of bioactive compounds than the other parts of the plant and are used as a cardiac stimulant, a vasodilator, and an antitumor and antibacterial drug (Chopra et al., 2013).
In recent years, studies regarding the antibacterial activity of P. corylifolia have been limited to the bacteriostasis of psoralen extracts rather than the antibacterial mechanism of P. corylifolia and its possible applications. Therefore, in this study, we analyzed the antibacterial activity and antibacterial mechanism of P. corylifolia seed ethanol extract (PCEE) against foodborne bacteria. The applications of PCEE regarding field strains collected from our previous studies were also analyzed.
Materials and Methods
Bacterial strains
MRSA ATCC 33591, L. monocytogenes ATCC 19115, E. coli ATCC 2592, S. aureus ATCC 6538P, and Salmonella typhimurium ATCC 23566 were purchased from the Bioresource Collection and Research Center. Strains were stored in tryptic soy broth at −20°C and reactivated twice before further studies. Twenty L. monocytogenes strains isolated from 2 tilapia sashimi production plants (Chen et al., 2015) and 10 S. aureus strains isolated from pig nasal and environmental waste water samples (kindly provided by Dr. Chin-Cheng Chou of the School of Veterinary Medicine, NTU) were also tested for minimum inhibitory concentration (MIC) of PCEE.
P. corylifolia seed ethanol extract
P. corylifolia seeds (5.4 kg) were pulverized and extracted with ethanol. In the extraction process, the pulverized seeds were soaked in 11 L ethanol at room temperature for 1 day for four times, and soaked in 11 L ethanol at 70°C for 4 h for five times. After filtering the extract, the filtrates were combined and concentrated to gain the ethanolic extract (1.4 kg). PCEE was further dissolved in 5% dimethyl sulfoxide (DMSO) before use.
Antimicrobial activity
Antimicrobial activity was determined using an agar-well diffusion test as described by Olajuyigbe and Afolayan (2012) with some modifications. Bacteria were diluted with 0.1% peptone water to 106 colony-forming unit (CFU)/mL and spread onto Mueller-Hinton agar (MHA). Eight-millimeter diameter wells were then made with a sterile plastic borer. PCEE (50 μL of 128 mg/mL) was added to each well, and ciprofloxacin and 5% DMSO were used as a positive and negative control, respectively.
MIC and minimum bactericidal concentration
MIC and minimum bactericidal concentration (MBC) were determined as described by Silva et al. (2011) with some modifications. PCEE was diluted with two-fold serial dilutions to a final concentration of 6.25–12,800 μg/mL. Bacteria were collected, washed, and diluted with phosphate-buffered saline (PBS) to ∼106 CFU/mL. MIC analysis was performed in microtitration plates with 10 μL PCEE, 20 μL bacterial suspension, and 180 μL Mueller-Hinton broth followed by incubation at 37°C for 24 h. MIC was determined by naked eyes, and MBC analysis was performed by subculturing 100 μL of the “no visible growth” MIC samples onto MHA and incubating at 37°C for 24 h.
Time-kill analysis
The time-kill analysis of PCEE was performed as described by Bajpai et al. (2013) with minor modifications. PCEE at the MBC (500 μL) was mixed with 8.5 mL Mueller-Hinton Broth (MHB) in a test tube. The bacterial suspension (1 mL of ∼106 CFU/mL) was then added to the PCEE/MHB solution and incubated at 37°C with shaking at 150 rpm for 24 h. The viable count of each assay was performed every 2 h. MHB alone with the bacterial sample was included as a control.
Cell membrane integrity
Membrane integrity was measured as previously described (Lv et al., 2011; Zhao et al., 2015) with some modifications. Bacteria were collected, washed, and diluted with PBS to ∼106 CFU/mL. The bacteria suspensions (5 mL) were then treated with three different concentrations of PCEE (control, MIC, and MBC) and incubated at 37°C for 4 h (L. monocytogenes) or 14 h (MRSA) according to the results of the time-kill analysis. The suspensions were then centrifuged at 6000 g for 5 min and the supernatant collected. The released constituents were measured by measuring the absorption at 260 nm and using Bradford protein quantification method (Bradford, 1976). Ciprofloxacin was used as a negative control (Devi et al., 2010).
Cell membrane permeability
Membrane permeability was measured using a conductivity meter (CON500; CLEANL'EAU, Taiwan) according to Zhao et al. (2015) with some modifications. Bacteria were collected by centrifugation at 6000 g for 5 min, washed three times with 0.1 M PBS (pH 7.4), and resuspended in PBS. Bacterial suspensions (5 mL) were treated with 0 and 1.6 mg/mL PCEE and incubated at 37°C. The conductivity was measured every 2 h for 24 h.
Scanning electron microscopy
Bacterial morphology was observed by scanning electron microscopy (SEM) as described previously by Wang et al. (2013). Bacteria were diluted with PBS to ∼106 CFU/mL. Bacterial suspensions (5 mL) were treated with 0 and 1.6 mg/mL PCEE and incubated at 37°C for 4 h (L. monocytogenes) or 14 h (MRSA) according to the results of the time-kill curve. The sample was fixed and processed for preparing specimens and then visualized using a scanning electron microscope (HitachiS4700, Japan).
Quantitative reverse transcription PCR
Quantitative reverse transcription PCR (RT-qPCR) was performed as previously described by Bae et al. (2014) and Romanova et al. (2006) with some modifications. Total RNA extractions were performed using Macherey NucleoSpin® RNA (Macherey-Nagel, Düren, Germany) and then Nanodrop-1000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA) was used to confirm the quality of RNA. RT-qPCR experiments were performed using the LightCycler® EvoScript RNA SYBR® Green I Master (Roche, Basel, Switzerland) with the LightCycler® 480 Real-Time PCR System (Roche). L. monocytogenes target genes were mdrL, mdrT, and lde, with GAPDH used as a reference (Bae et al., 2014). MRSA target genes were norA, mepA, and abcA, with 23S rRNA used as a reference (Romanova et al., 2006; Cervinkova et al., 2013; and Villet et al., 2014; Dabul et al., 2018). For each reaction, the concentration of RNA was adjusted to 100 μg/mL and combined with 0.5 μM of each primer in the SYBR Green reaction mixture. Results were expressed as the ratio of target gene to reference gene expression using the ΔΔCT method (Bae et al., 2011).
Statistical analysis
A completely randomized design with three replications was performed and data expressed as the mean ± standard deviation. One-way analysis of variance using Fisher's protected Least Significant Difference to determine the significant differences between the means (p < 0.05).
Results
Antibacterial activity analysis
PCEE had antibacterial effects on Gram-positive bacteria, with inhibition zone diameters of 14, 16, and 17 mm for S. aureus, MRSA, and L. monocytogenes, respectively. No inhibition zone was observed for selected Gram-negative bacteria and 5% DMSO. Therefore, S. aureus, MRSA, and L. monocytogenes were selected for subsequent MIC and MBC tests. An MIC of 50 μg/mL for MRSA and L. monocytogenes, and an MIC of 100 μg/mL for S. aureus were determined. The MBC of PCEE for L. monocytogenes and MRSA was 100 μg/mL, and for S. aureus was 200 μg/mL. In addition, the 5% DMSO treatment showed no bactericidal effect (Table 1).
Minimum Inhibitory Concentration and Minimum Bactericidal Concentration of Psoralea corylifolia Seed Ethanol Extract Against Listeria monocytogenes ATCC 19115, Staphylococcus aureus ATCC 6538P, and Multidrug-Resistant Staphylococcus aureus ATCC 33591
TNTC, too numerous to count; MIC, minimum inhibitory concentration; MBC, minimum bactericidal concentration; DMSO, dimethyl sulfoxide; MRSA, methicillin-resistant Staphylococcus aureus.
The 20 L. monocytogenes isolates had MICs of 6.25 (1 strain), 12.5 (5 strains), 25 (12 strains), and 50 (2 strains) μg/mL, and the 10 S. aureus isolates had MICs of 50 (1 strain) and 100 (9 strains) μg/mL. All field isolates of both bacteria had equal or less resistance to PCEE, indicating a potential application for PCEE.
Time-kill analysis
Based upon the MIC and MBC analyses, PCEE had more effective antibacterial activity on L. monocytogenes and MRSA, which were selected for follow-up experiments. An MBC of 100 μg/mL was used for the time-kill analysis of both L. monocytogenes and MRSA. The viability of L. monocytogenes was severely reduced after 2 h of exposure, and complete inhibition was achieved after 4 h of exposure to PCEE. The viability of MRSA was gradually reduced, achieving complete inhibition after 14 h of exposure (Fig. 1).
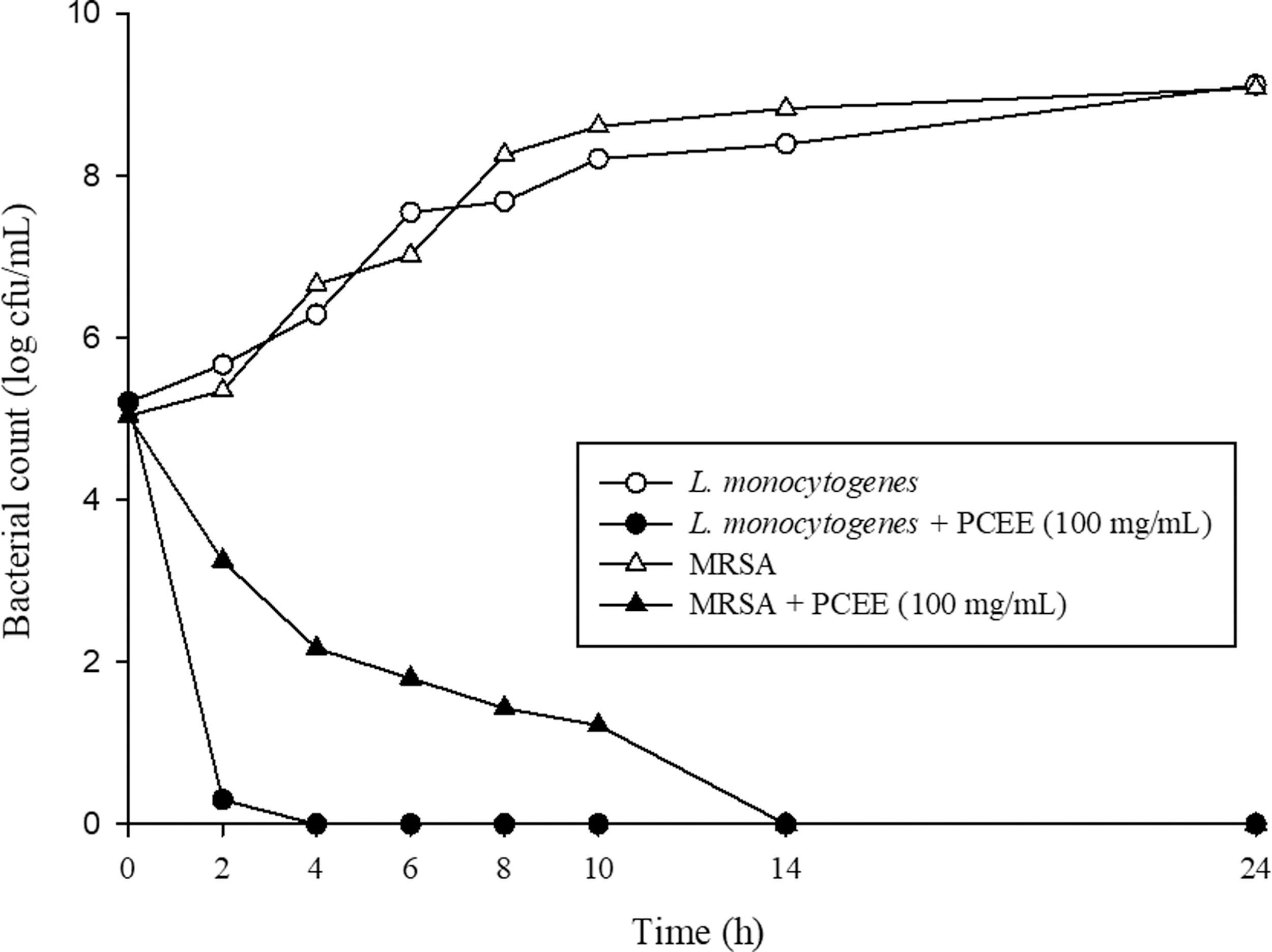
The effect of PCEE on the viability of Listeria monocytogenes ATCC 19115 and MRSA ATCC 33591. PCEE, Psoralea corylifolia seed ethanol extract; MRSA, methicillin-resistant Staphylococcus aureus.
Cell membrane integrity
The OD260 for both bacteria significantly increased (p < 0.05) with increasing concentrations of PCEE. The OD260 values of L. monocytogenes were 0.043 ± 0.006, 0.090 ± 0.005, and 0.113 ± 0.008 for the control, MIC, and MBC, respectively. The OD260 values of MRSA were relatively lower, 0.030 ± 0.001, 0.059 ± 0.008, and 0.081 ± 0.008 for the control, MIC, and MBC, respectively. There was no significant difference in either of the bacteria upon treatment with ciprofloxacin (Fig. 2).
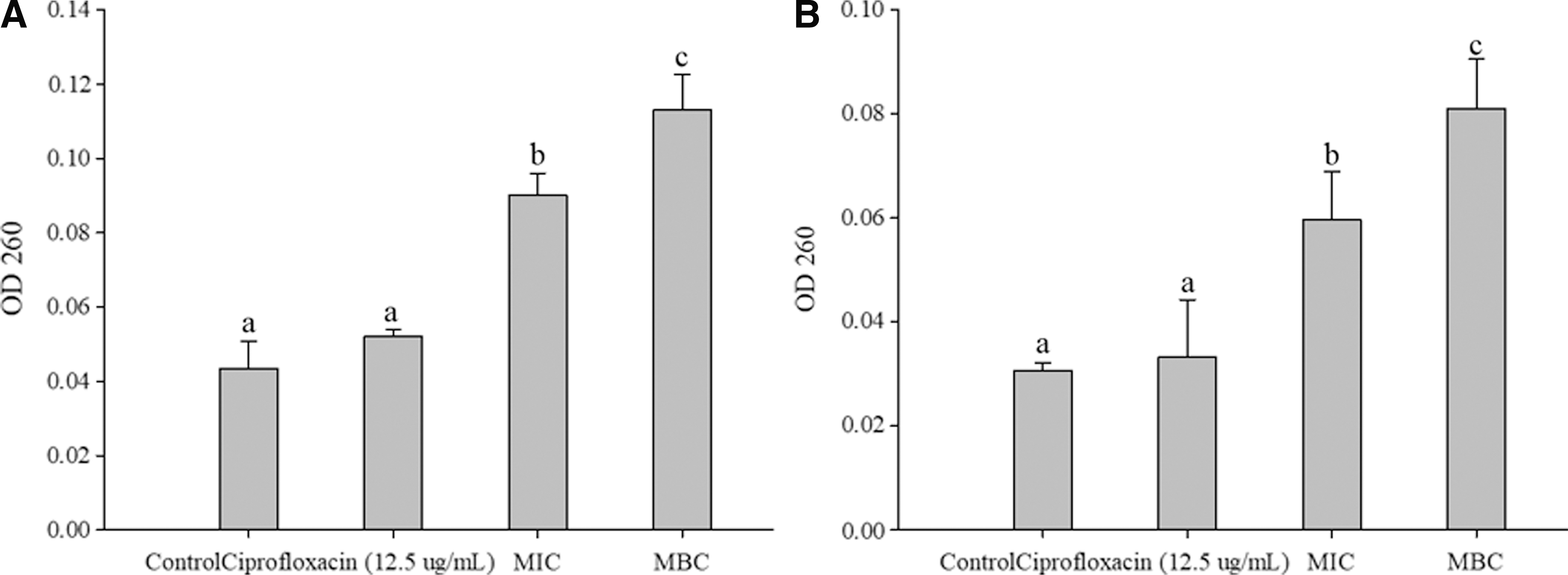
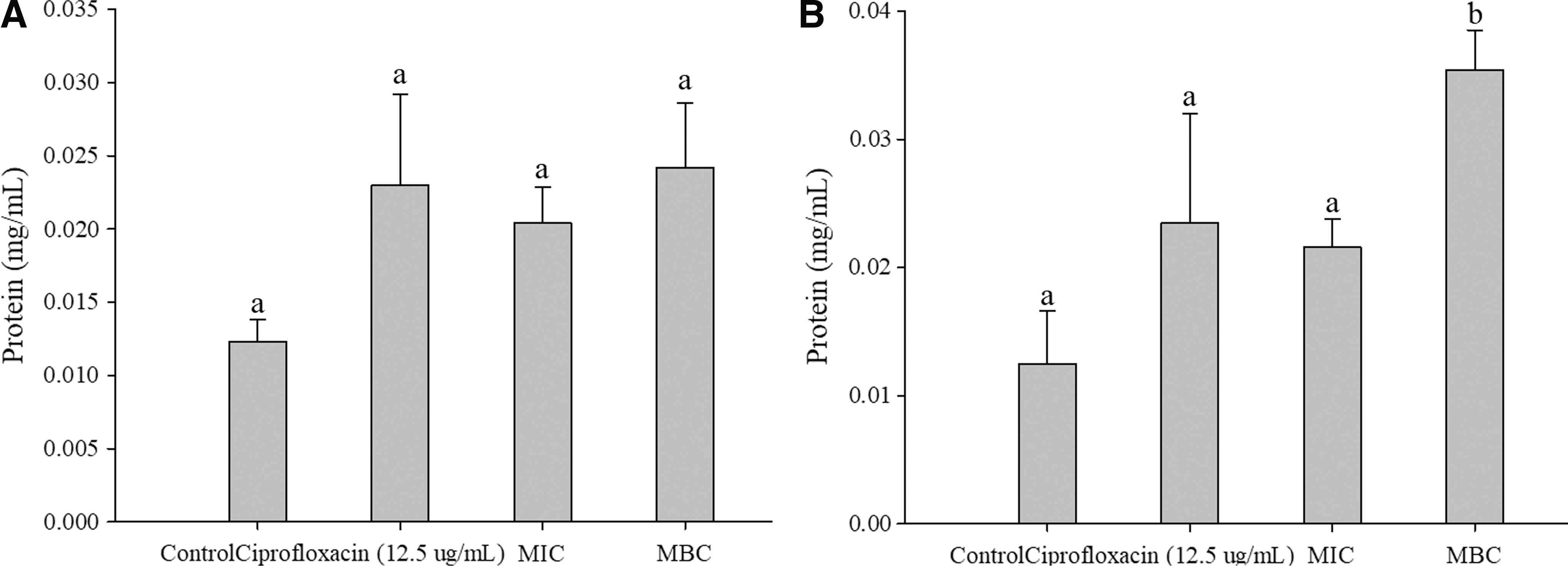
There was a significant difference (p < 0.05) in MRSA protein leakage between MBC (0.035 ± 0.003 mg/mL), MIC (0.022 ± 0.002 mg/mL), and the control (0.013 ± 0.003 mg/mL). However, although L. monocytogenes protein leakage showed a slight increase, there was no significant difference among the three groups: control (0.012 ± 0.001 mg/mL), MIC (0.021± 0.002 mg/mL), and MBC (0.024 ± 0.004 mg/mL). The protein concentrations for both L. monocytogenes and MRSA treated with ciprofloxacin were 0.023 ± 0.015 mg/mL and 0.023 ± 0.009 mg/mL, respectively, which were not significantly different from the control (Fig. 3).
Cell membrane permeability
The conductivity of MRSA and L. monocytogenes ranged from 12.990–13.057 to 12.927–12.990 μs/cm, respectively, with no significant changes over 24 h (Fig. 4). Moreover, the difference of initial conductivities of the control and tested group was due to the low conductivities and buffer, which lead to difficulty in comparison. However, the conductivity of MRSA and L. monocytogenes increased significantly for the first 16 h (12.970 μs/cm) and 8 h (13.227 μs/cm), respectively, then remained stable until the experiment ended at 24 h.

Alteration of cell morphology
Untreated L. monocytogenes cells were ∼1–2 μm in length, intact, rod-shaped, and had a smooth surface (Fig. 5A). In contrast, L. monocytogenes cells treated with 1600 μg/mL PCEE exhibited deformation, were broken, or even perforated (Fig. 5B). Untreated MRSA cells were ∼1 μm in diameter, and of a spherical-shape with a smooth surface (Fig. 5C), whereas MRSA cells treated with 1600 μg/mL PCEE were collapsed and deformed (Fig. 5D).

Morphology of bacterium visualized by scanning electron microscopy.
Quantitative reverse transcription PCR
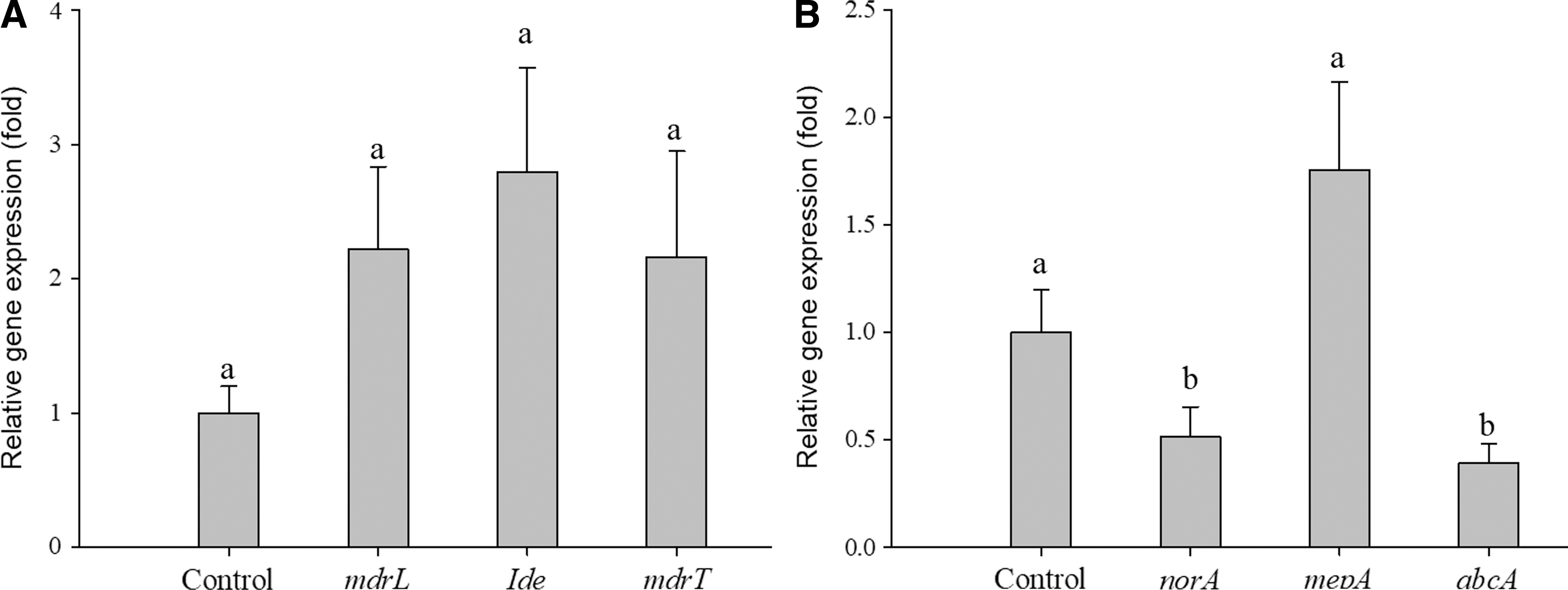
To further understand the interaction between PCEE and the responses of the bacteria, the gene expression of drug-resistant efflux pumps was examined. We found that L. monocytogenes mdrL, mdrT, and lde expression after PCEE treatment were 2.22-, 2.80- and 2.16-fold, respectively, of the control (Fig. 6A). This upregulation indicates a potential increase in the resistance of cells, but the differences were not statistically significant (p > 0.05). Low expression of MRSA abcA, norA, and mepA after PCEE treatment was observed (0.39-, 0.51-, and 1.75-fold, respectively) of the control. The significant downregulation of abcA and norA indicates that the resistance of MRSA was affected by the addition of PCEE, whereas mepA had a slightly higher expression, but the difference was not statistically significant (Fig. 6).
Discussion
We found that PCEE exhibited inhibitory effects on Gram-positive bacteria, yielding MICs of 50 μg/mL for MRSA and L. monocytogenes. However, there was no antibacterial effect on Gram-negative bacteria. Similar results were demonstrated that S. aureus and S. epidermidis were inhibited by derivatives of P. corylifolia, with MICs ranging from 0.009 to 0.147 mM (Yin et al., 2004). Gram-positive bacteria have a thick layer of peptidoglycan in cell wall, but this layer cannot prevent small molecular substances such as phenols, alcohols, and aldehydes from entering the cells. As a result, Gram-positive bacteria have a high sensitivity to these substances (Shimoda et al., 1995; Russell, 1999).
Extracts of plants, both crude extracts and essential oils, have been analyzed for their antimicrobial activity. Zuo et al. (2008) examined extracts of 19 plants and observed anti-MRSA activity with an MIC range of 1.25–3.07 mg/mL for all plants analyzed. Ethanol extracts of Garcinia cambogia, pomegranate, and Brassica inhibit MRSA ATCC 25923 with MICs of 100, 200, and 100 μg/mL, respectively, and inhibit MRSA hospital isolates with MICs of 50–400 μg/mL (Voravuthikunchai and Kitpipit, 2005). Jaradat et al. (2016) also found that thyme essential oil was effective against MRSA ATCC 25923 and field strain, yielding MICs of 250 and 500 μg/mL, respectively. Regarding PCEE in this study, interesting, both S. aureus and some field isolates of L. monocytogenes had higher MIC value comparing to L. monocytogenes 19115 and MRSA. Besides, the MIC for field isolates ranged from 6.25–100 μg/mL, lower than that of previously mentioned extracts or essential oils from other plants that display potential for further research or applications.
Damages to the cell membrane can be analyzed via the leakage of nucleic acids and proteins (Diao et al., 2014). The amounts of released nucleic acids increased significantly with increasing concentrations of PCEE for both L. monocytogenes and MRSA. Similarly, Lv et al. (2011) reported that a mix consisting of 10 essential oils caused damages to the cell membrane integrity of 4 foodborne bacteria. The OD260 increased with an increasing concentration of the essential oil mix; cinnamic aldehyde also acts on the cell membrane of E. coli and S. aureus, resulting in the release of nucleic acids (Shen et al., 2015). In our study, the increase in OD260 suggests that PCEE damaged the cell membrane of both L. monocytogenes and MRSA.
Regarding protein leakage, a significant increase was observed only with PCEE MBC-treated MRSA. Proteins are larger than nucleic acids, which may allow them to pass through the damaged membrane of the PCEE MBC-treated MRSA, whereas the other treatments did not cause such severe damage. Another possibility is that, based upon the low protein concentration measured, the sensitivity of detection was poor. However, all treatments displayed slightly increased protein concentration, indicating that damage may be inflicted by PCEE. PCEE is composed of phenol, hydrazine, aldehyde, and ketone, among others. The hydrophobic properties of these components can accumulate in the cell membrane and destroy the cell structure, causing the loss of internal substances and the reduction of enzymes, eventually killing bacteria (Sikkema et al., 1995). Therefore, PCEE could possibly change the membrane integrity of L. monocytogenes and MRSA, releasing nucleic acids and proteins, resulting in bacterial death. The cell membrane as an equipotential membrane acts as a permeability barrier for small ions, such as potassium and sodium (Singleton, 2004). Maintaining ion homeostasis is crucial for cells to maintain normal metabolism. Therefore, slight changes in the cell membrane can affect the metabolism of cells, resulting in death (Cox et al., 2001). Furthermore, electrolyte leakage can be used as an indicator of destruction of the permeability barrier (Diao et al., 2014). In this study, the extracellular conductivities of both L. monocytogenes and MRSA treated with 1600 μg/mL PCEE increased significantly, indicating that PCEE changed the cell membrane permeability, resulting in the release of electrolytes. This same antimicrobial mechanism was demonstrated in various studies (Shen et al., 2015; Zhao et al., 2015 and Zhang et al., 2016). Moreover, SEM images showed that PCEE caused morphological changes such as deformations, cracks, dents, and perforations of L. monocytogenes and MRSA cells. These results verified that the antimicrobial mechanism of PCEE was to cause damage to cell walls and membranes, resulting in cell contents leakage, which was consistent with our cell membrane integrity and permeability results.
Bacteria resistance to antimicrobial agents could be affected by several factors, including efflux pumps, cross-resistance, and mutations (Chuanchuen et al., 2001). When bacteria encounter stress, the efflux pumps would express to overcome the adversity. Rand et al. (2002) confirmed that nutritional restriction induces the reducing growth rate of E. coli due to downregulation of the multidrug efflux pump, acrAB-tolC. However, low glucose levels could stimulate cyclic adenosine monophosphate (cAMP) synthesis and increase the expression of acrAB through the cAMP receptor protein (Notley-MacRobb et al., 1997). By upregulating acrAB, bacteria would have a greater chance of survival in restrictive environments. The same situation was observed regarding regulation of the Pseudomonas aeruginosa mexA-mexB-oprM transporter (Evans and Poole, 1999; Adewoye et al., 2002). Therefore, PCEE may increase the gene expression of mdrL, mdrT, lde, and mepA and affect the permeability of bacterial cell membranes as the bacteria attempt to discharge PCEE. However, efflux pump inhibitors could deplete bacterial energy by interfering with proton dynamics or inhibiting adenosine triphosphate (ATP) binding, resulting in downregulation of efflux pump gene expression or inhibition of efflux pump activity in combination with hydrophobic regions of transport proteins (Markham et al., 1999; Marquez, 2005). Therefore, PCEE may decrease gene expression of norA and abcA by interfering with proton motive force and inhibiting ATP binding.
Conclusions
The PCEE exhibited good antibacterial activity against selected foodborne pathogens in this study. A possible mechanism is that the PCEE penetrates the membrane and then changes the metabolism of cell causing an energy depletion, either by inhibiting ATP or by disturbing the proton gradient, resulting in membrane damage. However, the direct damages to the cell wall and membrane cannot be ruled out. Further studies are needed for thorough understanding of the mechanism to expand the PCEE applications as an antibacterial agent.
Footnotes
Acknowledgment
This was supported by the research grant from Office of Research and Development (#8053), Fu Jen Catholic University.
Disclosure Statement
No competing financial interests exist.