
Abstract
The development of coresistance to disinfectants and heavy metals contributes to the fitness of Listeria spp. in foods or food processing environments, where life-threatening Listeria monocytogenes coexist and coevolve with other Listeria spp. Despite extensive research on L. monocytogenes, coresistance to disinfectants and heavy metals is less documented for other Listeria spp. In this study, we screened 30 L. monocytogenes and 27 Listeria innocua isolates recovered from 273 swine samples for resistance to quaternary ammonium compound benzalkonium chloride (BC) and to heavy metals cadmium (Cd) and arsenic (As). Moreover, we evaluated the potential mechanisms of resistance by detecting the efflux pump activity in BC resistance and the presence of resistance determinants. The average minimum inhibitory concentrations of BC in L. innocua (10.7 ± 2.0) were significantly higher than that in L. monocytogenes (6.9 ± 3.7) (p < 0.05). Resistance to BC and heavy metals was correlated, where all BC-resistant L. innocua and As-resistant L. monocytogenes isolates were coresistant to BC and Cd. Twenty percent and 66.7% of BC resistance in L. monocytogenes and L. innocua were related to reserpine–associated efflux pumps, whereas all cases of BC resistance were related to carbonyl cyanide 3-chlorophenylhydrazone–associated efflux pumps. The cadA1 and cadA2 genes were present in Cd-resistant isolates but not in Cd-sensitive isolates, and cadA3 was undetectable in all isolates examined. cadA4 conferring lower level of Cd resistance was copresent with arsA1 and arsA2 in the Cd-resistant and As-susceptible L. monocytogenes isolate LM3. Our findings suggest that swine serves as a reservoir for developing resistance to disinfectant and heavy metals in L. monocytogenes and L. innocua, which share common resistance mechanisms such as efflux pumps and resistance genes. This work provides new insight into the coresistance events of other Listeria as a potential contributor of the resistance in L. monocytogenes.
Introduction
In food and food-processing environments, which are recognized as important reservoirs of disinfectant, heavy metal, and antibiotic resistance for Listeria monocytogenes and other Listeria (Katharios-Lanwermeyer et al., 2012; Jamali et al., 2015; Camargo et al., 2017; Cherifi et al., 2018), Listeria species such as Listeria innocua and Listeria welshimeri are more frequently isolated than the life-threatening L. monocytogenes (Chen et al., 2009, 2010; Jamali et al., 2015). Some L. innocua strains have also been associated with cases of fatal bacteremia and meningitis, and some virulence genes have been reported in the so-called nonpathogenic Listeria spp. (Gourin et al., 1994; Perrin et al., 2003; Johnson et al., 2004; Favaro et al., 2014). However, these Listeria species are less investigated for coresistance compared with L. monocytogenes.
Coresistance to disinfectants and heavy metals has attracted the interest of researchers owing to the potential for coselection and coevolution of resistance (Katharios-Lanwermeyer et al., 2012; Kovacevic et al., 2013). For example, plasmid harboring genes conferring resistance to disinfectant benzalkonium chloride (BC) and heavy metal cadmium (Cd) in L. welshimeri and L. innocua could be readily transferred to L. monocytogenes through conjugation, and Cd resistance transfer can be effectively used as a surrogate for transfer of resistance to BC, since transconjugants selected on Cd were resistant to both Cd and BC (Katharios-Lanwermeyer et al., 2012). With such coselection mechanism, Cd and BC coresistance L. monocytogenes, which frequently exist in food processing plants (Katharios-Lanwermeyer et al., 2012), will be able to gain fitness and be enriched in these environments.
Genetic links between Cd and BC resistance elements were found to be a main contributor to this coresistance (Elhanafi et al., 2010; Xu et al., 2016). Moreover, efflux pumps are a primary mechanism of multidrug resistance (Webber and Piddock, 2003; Blair et al., 2015). In L. monocytogenes, gene encoding efflux pumps have been reported to confer resistance to substances like ethidium bromide, BC, or heavy metals (Aase et al., 2000; Mata et al., 2000; Godreuil et al., 2003). The cadA1–A4, arsA1–2, and bcrABC resistance determinants investigated in the current study all confer resistance through the efflux pump activity (Mullapudi et al., 2008; Elhanafi et al., 2010; Briers et al., 2011; Parsons et al., 2017). This suggests that efflux pump may participate in coresistance, and transfer of efflux pump-associated genetic elements may represent one important aspect of the mechanisms of transferring coresistance.
In this study, we investigated the resistance of L. monocytogenes, as well as L. innocua isolates isolated from food of swine origin in Guangzhou and Xiamen of South China to the quaternary ammonium compound (QAC) disinfectant BC and heavy metals Cd and arsenic (As), evaluated the effect of efflux pumps in mediating BC resistance, and determined the presence of QAC, Cd, and As resistance genes. Overall, the findings should help to understand the risk of L. innocua, which share common coresistance mechanisms with L. monocytogenes, to transfer resistance to L. monocytogenes to increase their fitness and colonization and threat to the safety of food supply.
Materials and Methods
Bacterial isolates
The Listeria spp., used in this study, were recovered from 273 swine samples, including raw pork (n = 100), haslet (n = 25), and cooked meat products (n = 20) from supermarkets and fresh markets in Guangzhou and raw pork (n = 113) and haslet (n = 15) from a large-scale pork-processing plant (>5000 pigs) in Xiamen in 2012, where a total of 30 L. monocytogenes and 27 L. innocua isolates were identified. Isolation and identification of L. monocytogenes and L. innocua were performed according to the National Standards of the People's Republic of China (GB/T 4789.30–2010). 16S rDNA polymerase chain reaction (PCR) was carried out to confirm the species of the isolates, with the universal primers 27F (5′-AGAGTTTGATCMTGGCTCAG-3′) and 1492R (5′-TACGGYTACCTTGTTACGACTT-3′) (Lane, 1991).
Serotyping by multiplex PCR
Multiplex PCR was conducted to distinguish five major serotypes of L. monocytogenes (1/2a 3a, 1/2b 3b, 1/2c 3c, 4a 4c, and 4b 4d 4e) as previously described (Doumith et al., 2004). In brief, one multiplex PCR assay for four marker genes, lmo0737, lmo1118, ORF2819, and ORF2110, and an internal amplification control for the genus Listeria, prs, was conducted.
BC, Cd, and As susceptibility tests
The minimum inhibitory concentration (MIC) of BC, Cd, and As was determined by the agar dilution method using Mueller Hinton (MH; Guangdong Huankai Microbial Sci&Tech, Guangzhou, China) agar (for BC susceptibility test) or IsoSensitest agar (ISA; Oxoid, Basingstoke, England) (for Cd and As susceptibility test) plates containing 1.2% defibrinated sheep blood as described previously with minor alterations (Mullapudi et al., 2008; Elhanafi et al., 2010). Briefly, a single colony from a BHI agar plate culture was suspended in 100 μL of tryptic soy broth. Five microliters of suspension was spotted in duplicate onto MH agar plates containing 0, 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 26, and 30 μg/mL BC and incubated at 30°C for 48 h or to ISA plates containing 0, 35, 70, 140, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1100, and 1200 μg/mL of cadmium chloride anhydrous (Sigma, St. Louis, MO) or 250, 500, and 1000 μg/mL of sodium arsenite (Sigma) and incubated at 37°C for 48 h.
Efflux pump inhibition with reserpine and carbonyl cyanide m-chlorophenylhydrazone
A single colony from a BHI agar plate culture was suspended in 100 μL of tryptic soy broth. Five microliters of suspension was spotted in duplicate onto MH agar plates containing 20 μg/mL reserpine (Sigma) or 100 μmol/L carbonyl cyanide m-chlorophenylhydrazone (CCCP; Aladdin, Shanghai, China) and different concentrations of BC as described above (Romanova et al., 2006). After 48 h incubation at 37°C, MICs of BC were observed and compared with the MICs measured in the absence of the inhibitors. Five microliters of suspension was spotted in duplicate onto agar plates containing reserpine or CCCP but without BC; if the spotted area is full of colonies after 48 h, it indicates that the inhibitors have no inhibitory effect on growth.
Detection of QAC, Cd, and As resistance genes
PCR amplification was used to screen for the QAC resistance genes qacA/B, qacC/D, qacE, qacEΔ1-sul, qacF, qacG, qacH, qacJ, and bcrABC; Cd resistance genes cadA1, cadA2, cadA3, and cadA4; and As resistance genes arsA1 and arsA2. The primers are listed in Table 1 (Ploy et al., 1998; Mereghetti et al., 2000; Sidhu et al., 2002; Bjorland et al., 2003; Zhou and Lu, 2005; Gillings., 2010; Mullapudi et al., 2010; Lee et al., 2013). PCR products were sequenced at Beijing Genomics Institute (Shenzhen, China) and analyzed using the BLAST algorithm (
Primers Used for Polymerase Chain Reaction Amplification of the Quaternary Ammonium Compound, Cadmium, and Arsenic Resistance Genes
Statistical analysis
Statistical analysis was conducted to compare the frequency of cadA2 gene with chi-square tests (other Cd and As resistance genes are not subjected to statistical analysis because of the limited number of isolates with these genes) and compare the BC, Cd, and As MICs with Welch's t-test through SAS (version 9.13; SAS Institute, Cary, NC). Differences with p < 0.05 were considered as statistically significant. Pearson correlation coefficient between the MICs of BC, Cd, and As was analyzed using Pearson Correlation Coefficient Calculator (
Results
MICs for BC, Cd, and As
In this study, we designated isolates with BC MIC >10 μg/mL as resistant according to previous studies (Romanova et al., 2002; Sidhu et al., 2002). In L. monocytogenes, two distinct populations of susceptible (25/30, 83.3%) or resistant (5/30, 16.7%) were identified. In L. innocua, the majority of the isolates showed a MIC of 10 μg/mL (Table 2 and Fig. 1A, D); thus, isolates with a BC MIC >10 μg/mL were scored as resistant (9/27, 33.3%). The MICs of BC are significantly higher in L. innocua than in L. monocytogenes (p < 0.05) (Table 2 and Fig. 1A, D).
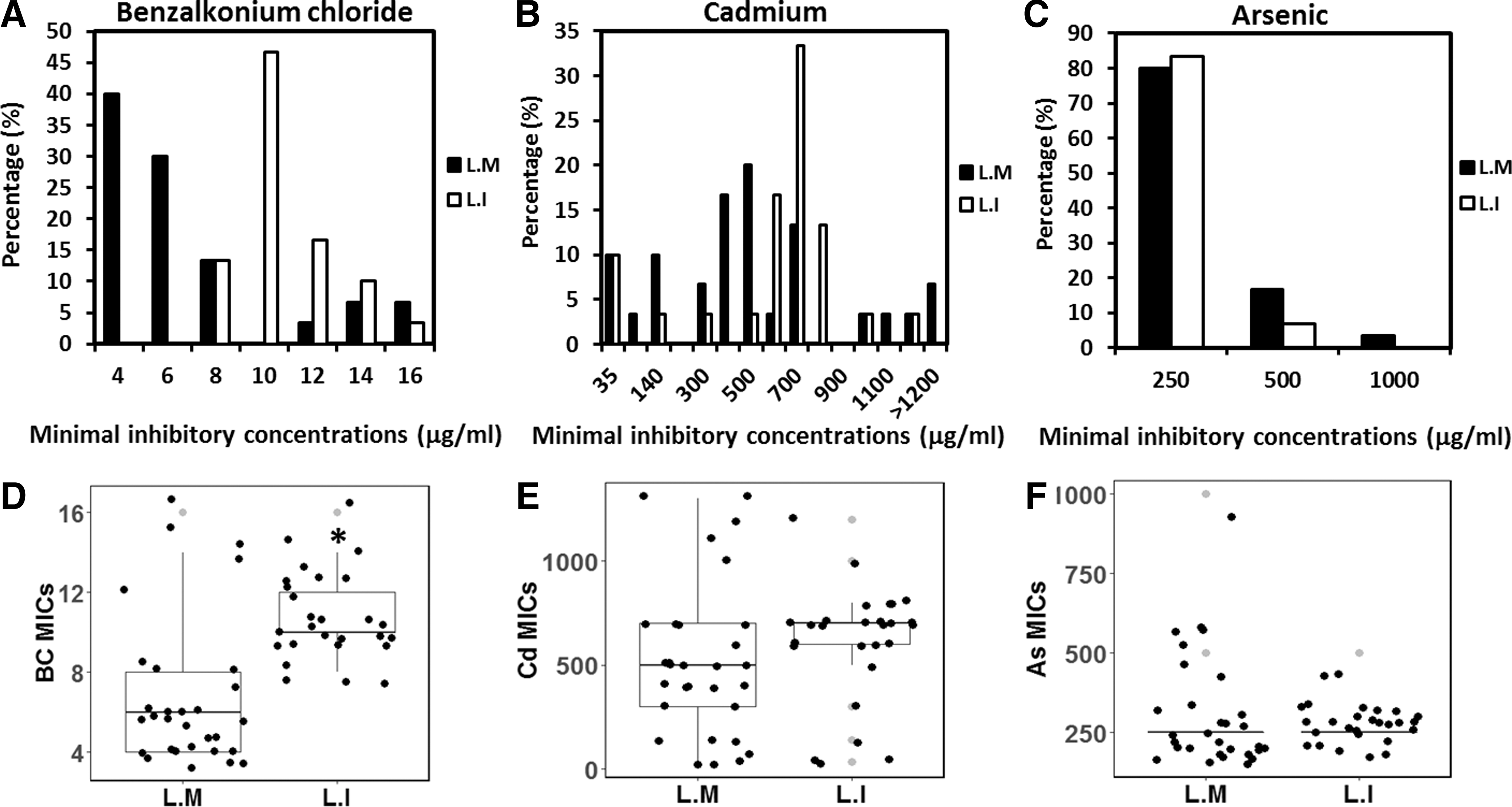
Distribution of MICs of BC
Profiles of Benzalkonium Chloride, Cadmium, and Arsenic Resistance and the Resistance Determinants
G, Guangzhou; X, Xiamen.
C, cooked meat; R, raw pork.
Bold font represents resistance.
Cd-R gene, Cd resistance gene, white and black boxes represent negative and positive PCR results, respectively, for cadA1, cadA2, cadA3, and cadA4.
As-R gene, As resistance gene, white and black boxes represent negative and positive PCR results, respectively, for arsA1 and arsA2.
L. M, Listeria monocytogenes.
L. I, Listeria innocua.
As, arsenic; BC, benzalkonium chloride; Cd, cadmium; MICs, minimum inhibitory concentrations; PCR, polymerase chain reaction.
For Cd and As, isolates with MIC >35 or >500 μg/mL were considered to be Cd- or As resistant, respectively (Mullapudi et al., 2008; Lee et al., 2013). The resistance to Cd was high for both L. monocytogenes (27/30, 90.0%) and L. innocua (24/27, 88.9%). Two (6.6%) L. monocytogenes isolates with serotypes 1/2c 3c or 1/2a 3a had a MIC over 1200 μg/mL (Table 2 and Fig. 1B). By contrast, the prevalence of As resistance was relatively low in both species, with only 1 (3.3%) L. monocytogenes isolate showing resistance (Table 2 and Fig. 1C).
Five (16.7%) L. monocytogenes and 7 (25.9%) L. innocua isolates were coresistant to BC and Cd (Table 2). The MICs of BC and Cd showed moderate positive correlation in both L. monocytogenes (R = 0.67) and L. innocua (R = 0.50). There's only weak correlations between the MICs of either BC or Cd to As.
Serotypes of L. monocytogenes isolates
The number of L. monocytogenes with serotypes 1/2a 3a, 1/2b 3b, 1/2c 3c, 4a 4c, and 4b 4d 4e is 17, 5, 6, 2, and 0, respectively (Table 2). Since the number of L. monocytogenes isolates in each serotype is limited, it is not sufficient to analyze the serotype-specific distribution of resistance from a statistical point of view.
Efflux pump inhibition by reserpine and CCCP
All BC-resistant L. monocytogenes and L. innocua isolates became susceptible to BC with CCCP treatment, whereas only 1 (20.0%) L. monocytogenes isolate and 6 (66.7%) L. innocua isolates were rendered susceptible upon reserpine treatment (Fig. 2).
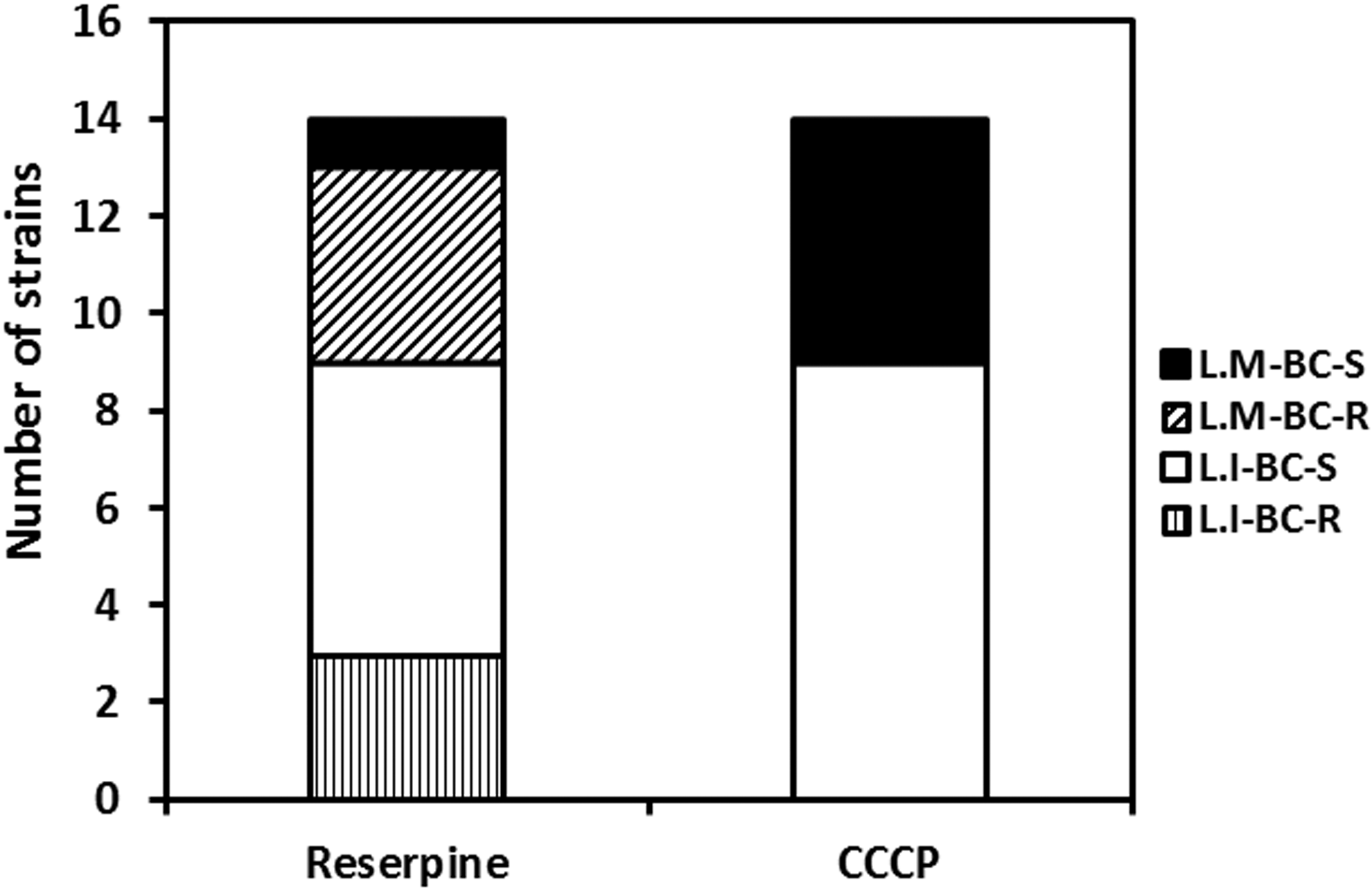
Susceptibility to BC of the resistance strains in the presence of reserpine or CCCP. L.M-BC-S, sensitive Listeria monocytogenes strains to BC in the presence of inhibitors; L.M-BC-R, resistance L. monocytogenes strains to BC in the presence of inhibitors; L.I-BC-S, sensitive Listeria innocua strains to BC in the presence of inhibitors; L.I-BC-R, resistance L. innocua strains to BC in the presence of inhibitors. BC, benzalkonium chloride; CCCP, carbonyl cyanide m-chlorophenylhydrazone.
Prevalence of QAC, Cd, and As resistance genes
QAC family or bcrABC genes were undetectable in any of the isolates examined. Cd resistance genes cadA1, cadA2, and cadA4 were detected in 4 (13.3%), 18 (60.0%), and 1 (3.3%) L. monocytogenes isolates and in 2 (7.4%), 6 (22.2%), and 0 (0%) L. innocua isolates, respectively, whereas cadA3 was not detectable (Table 2 and Fig. 3). The frequency of cadA2 was significantly higher in L. monocytogenes than in L. innocua (p < 0.05; Table 2 and Fig. 3). Prevalence of cadA1 and cadA4 cannot be subjected to statistical analysis because of the limited number of isolates harboring these genes. All isolates harboring cadA1, cadA2, and cadA4 were resistant to Cd, and it is noteworthy that cadA4-harboring isolate LM3, which is of serotype 4a 4c, conferred low level of Cd resistance (MIC = 70 μg/mL) (Table 2). LM3 is also the only isolate that harbored arsA1 and arsA2 (Table 2).
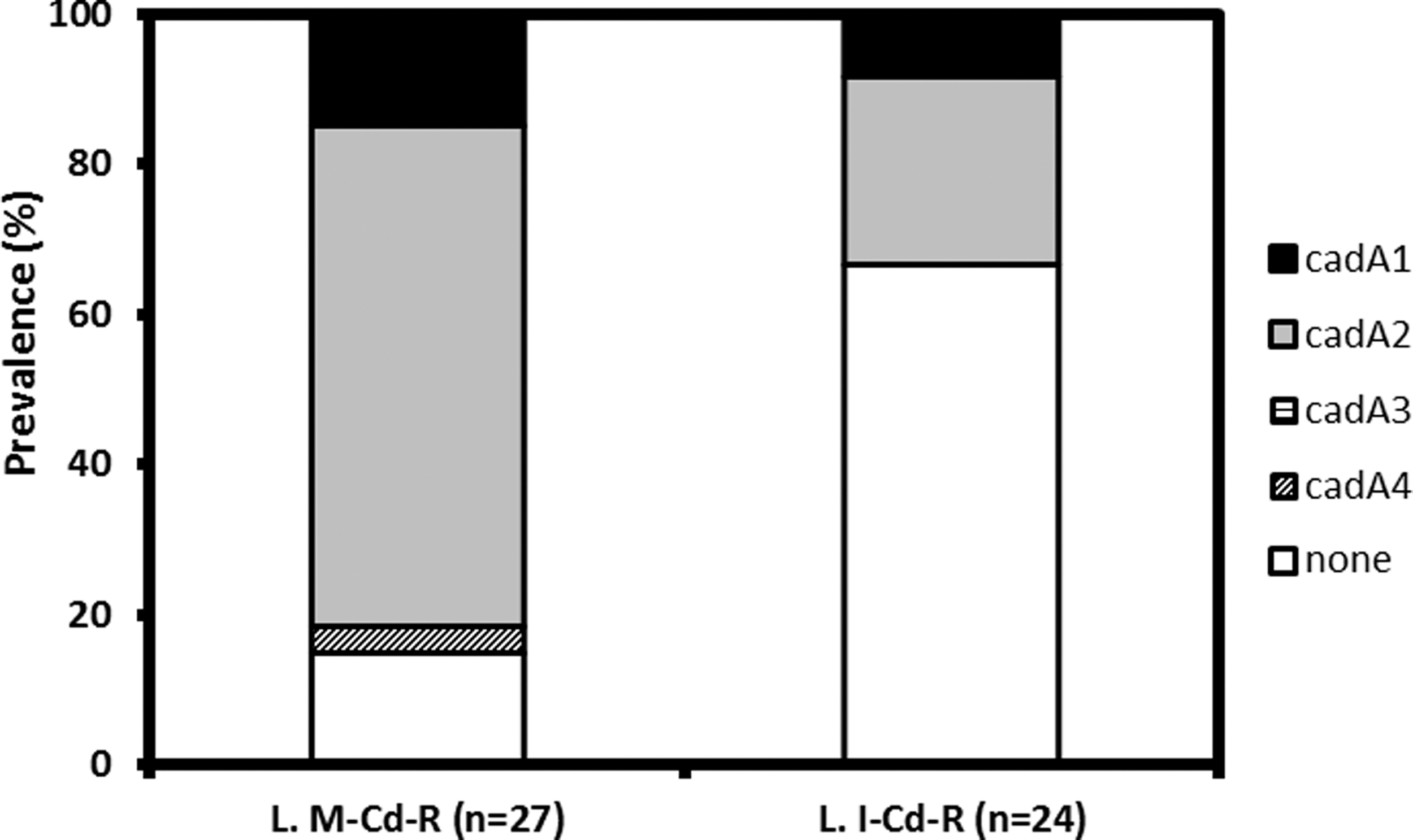
Prevalence of different Cd resistance determinants among Cd resistant Listeria monocytogenes and Listeria innocua strains. L.M-Cd-R, Cd resistance L. monocytogenes strains; L.I-Cd-R, Cd resistance L. innocua strains. Cd, cadmium.
Discussion
The prevalence of BC resistance in the current study (16.7% in L. monocytogenes and 33.3% in L. innocua) is comparable to that reported previously (10–46%) (Sidhu et al., 2002; Soumet et al., 2005; Mullapudi et al., 2008; Katharios-Lanwermeyer et al., 2012; Jiang et al., 2016). However, the prevalence of Cd resistance (90.0% in L. monocytogenes and 88.9% in L. innocua) was much higher than those previously reported (49–66%) (Mullapudi et al., 2008; Katharios-Lanwermeyer et al., 2012; Xu et al., 2014), suggesting that Listeria spp. isolated from different sources may possess diverse resistance to Cd, possibly due to exposure to different environmental stresses that select for Cd resistance. Resistance to As was less common compared with BC or Cd resistance, comparable to the result of a previous study (Mullapudi et al., 2008). It is noteworthy that 2 (6.6%) L. monocytogenes isolates had a Cd MIC higher than 1200 μg/mL, which has been rarely reported. This may be partially due to the fact that one breakpoint concentration like 35, 70, or 75 μg/mL was often used to determine Cd resistance (Mullapudi et al., 2008; Katharios-Lanwermeyer et al., 2012; Ratani et al., 2012; Lee et al., 2013). It is worthwhile to use a wider range of Cd concentrations to screen for Cd resistance. In addition, the cadA4-harboring isolate LM3 that showed a slightly higher MIC than those susceptible strains lacking any of the examined cad genes would be defined as susceptible if a MIC breakpoint of 70 μg/mL or higher was used, indicating that the MIC breakpoint need to be carefully selected.
The prevalence of L. monocytogenes with BC, Cd, and As resistance did not show a serotype-specific trend, in contrast to previous studies (Mullapudi et al., 2008; Xu et al., 2014). A larger sample size would contribute to figuring out the serotype-associated prevalence of resistance.
There are moderate positive correlations of BC and Cd MICs in both L. monocytogenes (R = 0.67) and L. innocua (R = 0.50) according to Pearson correlation coefficient analysis, in agreement with a previous study (Mullapudi et al., 2008), suggesting the coexistence of BC and Cd resistance determinants or the existence of a resistance system that mediates both BC and Cd resistance. BC and Cd resistance determinants have been reported to colocalize in the same plasmid in L. monocytogenes and other Listeria spp. like L. welshimeri and L. innocua (Mullapudi et al., 2008; Elhanafi et al., 2010; Katharios-Lanwermeyer et al., 2012; Xu et al., 2016). L. monocytogenes isolates of various serotypes and from diverse sources were found to harbor bcrABC genes together with cadA1 or cadA2 on pLM80-like plasmids (Dutta et al., 2013).
Efflux pumps represent a mechanism for multidrug resistance (Aase et al., 2000; Mata et al., 2000; Godreuil et al., 2003; Webber and Piddock, 2003; Blair et al., 2015). Reserpine is considered to inhibit members of the major facilitator superfamily (MFS), resistance nodulation division family (RND), and ATP-binding cassette superfamily (ABC) (Lee et al., 2003). CCCP has been shown to act on the efflux systems of the small multidrug resistance family, multidrug and toxic compound extrusion family, MFS, and RND (Bohnert and Kern, 2005; Rafii et al., 2005). In our study, CCCP-associated BC resistance was more prevalent than reserpine-associated BC resistance in both L. monocytogenes and L. innocua, suggesting that efflux pump related to CCCP play a major role in conferring BC resistance. Different types of efflux pump inhibitors can be used to identify the responsible efflux pump system and reveal the underlying resistance mechanisms.
None of the isolates analyzed was found to be positive for QAC family genes or bcrABC. In previous studies, QAC family genes are frequently detected in Staphylococcus aureus (Wu et al., 2008; Zmantar et al., 2011), but rarely documented in L. monocytogenes (Xu et al., 2014; Jiang et al., 2016). QAC genes in L. monocytogenes were suggested to be obtained by horizontal transfer from other species (Xu et al., 2014). In contrast, bcrABC, a BC resistance cassette found in either plasmid or chromosome, showed varied frequencies in Listeria spp. in different studies (Dutta et al., 2013; Jiang et al., 2016). Its absence from the present study suggests the variable genetic background of L. monocytogenes from different sources.
None of the isolates studied was positive for cadA3, similar to the results of Ratani et al. (2012). In L. monocytogenes, the prevalence of cadA2 was higher compared with cadA1 and cadA4, in contrast to the results of Ratani et al. (2012) in which cadA1 was found to be more prevalent than cadA2 (53% vs. 27%). In our study, none of the isolates harbored both cadA1 and cadA2, which was also reported in the previous study (Ratani et al., 2012), but conflicts with the results of other studies demonstrating that 30% and 2.9% of the Cd-resistant isolates harbored both cadA1 and cadA2, respectively (Mullapudi et al., 2010; Xu et al., 2014), suggesting that different selective pressures in different environments may differentially shape the genetic background of L. monocytogenes. Besides the present study, detection of the cadA1–A4 genes in L. innocua has only been reported in one current study (Korsak et al., 2019). In this study, we found 2 (7.4%) and 6 (22.2%) of the L. innocua isolates harbored cadA1 and cadA2 genes, respectively.
The cadA2 gene was more frequent in L. monocytogenes serotypes 1/2b 3b and 1/2c 3c than in serotype 1/2a 3a, although no statistically significant differences were obtained due to the limited number of each serotype analyzed. This finding is consistent with previous studies (Mullapudi et al., 2010; Ratani et al., 2012). Cd resistance was largely plasmid associated (Harvey and Gilmour, 2001), and cadA1 and cadA2 were associated with plasmid-borne transposon Tn5422 (Lebrun et al., 1994a, b) and large plasmids like pLM80 (Kuenne et al., 2010), respectively. Serotype-associated prevalence of Cd resistance determinants may be related to the serotype-specific trend of harboring plasmids (Harvey and Gilmour, 2001; Mullapudi et al., 2010).
It is also noteworthy that the cadA1 and cadA2 genes were only detected in Cd-resistant isolates, and the only cadA4 gene detected was harbored by LM3, which grew at 35 but not at 70 μg/mL Cd, showing a slightly higher MIC than other susceptible isolates (Table 2). This is consistent with prior studies demonstrating that strains harboring only the cadA4 Cd resistance cassette grew at 35 but not at 50 or 70 μg/mL Cd, probably due to the less conservation of cadA4 sequence compared with that of cadA1, cadA2, and cadA3 (Lee et al., 2013; Parsons et al., 2017), suggesting that the breakpoint of resistance MIC should be carefully defined.
Only one isolate, L. monocytogenes isolate LM41 was resistant to As, although neither of the As resistance determinants arsA1 or arsA2 was detected, suggesting the existence of novel As resistance determinants. In a study on L. monocytogenes serotype 4b isolates from sporadic human listeriosis patients, all 23 As-resistant isolates were positive for arsA2, and a subset (13/23) was also positive for arsA1, while none of the As-susceptible isolates harbored either arsA1 or arsA2 (Lee et al., 2013). However, in our study, the only L. monocytogenes isolate LM3 that harbored arsA1 and arsA2 was As susceptible. This same isolate was also positive for cadA4, in accordance with a previous study indicating that all isolates harboring both arsA1 and arsA2 also harbored cadA4 (Lee et al., 2013). The underlying association requires further investigation.
Conclusions
Our findings suggest that foods of animal origin such as swine are a reservoir of different Listeria species (L. monocytogenes and L. innocua), which share common resistance mechanisms such as efflux pumps or genetic resistance elements, highlighting the importance to continuously monitor the resistance among not only L. monocytogenes but also other Listeria spp. Future study with larger sample size would help to validate the findings of the current study and give more insight into the issues discussed here with a statistical point of view.
Footnotes
Acknowledgments
This work was supported by the National Natural Science Foundation of China (Grant No. 31571934), the Science and Technology Planning Project of Guangdong Province, China (2014A020214001), the Construction of the First Class Universities and Special Development Guidance Special Fund (K5174960), and the Science and Technology Planning Project of Guangdong Province, China (2014A020214001).
Disclosure Statement
No competing financial interests exist.