
Abstract
Consumer concerns toward chemical preservatives have resulted in increased search for healthy green alternative. In this study, the antioxidant activity and antibacterial effects of Eucalyptus camaldulensis ethanolic leaf extract against Listeria monocytogenes, a serious foodborne pathogen, was evaluated. Total phenolic and flavonoid contents of the extract were 11.10 mg garlic acid equivalent/mg extract and 15.05 mg quercetin equivalent/mg extract, respectively. The minimum inhibitory concentration (MIC) and minimum bactericidal concentration of the extract was 64–128 μg/mL and 256–512 μg/mL, respectively. Time-kill assay revealed growth inhibitory effects after 4-h treatment of the bacteria with the extract. A reduction of ≈2–3 log colony-forming units per milliliter was observed against the tested food and environmental isolates after challenging the pathogens with the extract at MIC for 6 h. Sub-MICs of the extract significantly inhibited motility and listeriolysin O production up to 80%, with 60% inhibition of biofilm formation (p < 0.05). Antioxidant assay revealed free radical scavenging activity with 50% inhibitory concentration (IC50) of 57.07 μg/mL for 2,2-diphenyl-1-picrylhydrazyl and 29.01 μg/mL for ABTS [2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid)] assay. Ferric reducing antioxidant power assay further showed a total antioxidant power equivalent to 92.93 μM ascorbic acid equivalent/mg extract. As the extract exhibited profound antilisterial activity and good radical scavenging ability, it might serve as a potential alternative source of biopreservative agent against L. monocytogenes.
Introduction
Listeria monocytogenes is a common foodborne pathogen that is ubiquitous to various environments. It is responsible for the invasive foodborne disease “listeriosis,” acquired through the ingestion of contaminated food products, especially ready-to-eat meat products, unpasteurized milk, cheese, ice cream, and seafood (Burall et al., 2017). In healthy individuals, listeriosis presents a self-limiting infection with occasional mild symptoms. However, in immunocompromised individuals and pregnant women, it results in life-threatening conditions such as meningitis, encephalitis, miscarriage, and stillbirth (Buchanan et al., 2017; Datta and Burall, 2018). Although it is a rare infection, prevalence of listeriosis is relatively high among pregnant women accounting for 15% of all reported cases of Listeria infection in the United States (Mateus et al., 2013; McKinney, 2016) with about one-third fetal loss or neonatal death (Silk et al., 2012).
Persistence of L. monocytogenes in food environments and its tolerance to various food grade antimicrobials makes it a safety and health threat. With an ability to survive at temperature ranges of <1–45°C, acidic and alkaline pH and high salt concentrations range of 7.5–10%, L. monocytogenes constitutes a major concern to food handlers and safety personnel. Resistance to antimicrobial sanitizers and preservatives has been reported among different serotypes (Henriques and Fraqueza, 2017; Vázquez-Sánchez et al., 2017). Previous researchers suggested that Listeria biofilm formation is a critical adaptive mechanism, responsible for its survival (Nakamura et al., 2013; Henriques and Fraqueza, 2017).
In recent times, the food industry has witnessed an increase in consumer awareness and general distaste for chemical preservatives. The demands for minimally preserved food with unaltered nutritional and sensory qualities as well as failures of some age-long trusted preservatives suggest an end to the era of synthetic chemical preservatives. Concerned with the effect of these changes in trend, food industries, scientist, and nutritionist are in urgent search for effective alternative antimicrobials that would replace existing chemical agents.
Natural products like essential oils, phenolic compounds, flavonoids, and crude extracts of plant parts have demonstrated antimicrobial and antioxidant properties and thus might serve as an alternative source of potential preservatives that would be acceptable to consumers. Various plant species, including plants of the family Myrtaceae, have shown antimicrobial activity against food spoilage and foodborne microorganisms (Gottardi et al., 2016; Odedina et al., 2016). Essential oils from plants of the genus Eucalyptus are rich sources of antimicrobial and antioxidant products (Lima et al., 2018; Nasr et al., 2018) used in the pharmaceuticals, cosmetics, and food industries (Marzoug et al., 2011).
Eucalyptus camaldulensis (river red gum), belonging to the family Myrtaceae, is one among more than 900 species of the genus Eucalyptus. Rich in phytochemicals, it is a source of antimicrobial agent effective against numerous clinical, spoilage, and foodborne microorganisms (El-Ghorab et al., 2003; Chaves et al., 2018). In vivo experiments demonstrated that leaf extracts of E. camaldulensis produced no toxic effects (El-Ghorab et al., 2003; Imam and Wudil, 2011). Hence, this study aims to evaluate the antimicrobial and antivirulence activities of E. camaldulensis against L. monocytogenes. The effects of the extract on membrane permeability and its antioxidant activities were also investigated.
Materials and Methods
Plant extraction
Classified reference voucher specimen of E. camaldulensis was deposited at the Herbarium of Faculty of Pharmaceutical Sciences, Prince of Songkla University, Thailand. The leaves were extracted as described in previous works from our laboratory (Limsuwan et al., 2009; Hiranrat et al., 2012).
Folin–Ciocalteu assay
The total phenolic contents of the extract were determined using Folin–Ciocalteu method (Kaur and Kapoor, 2002) at an absorbance of 700 nm. Total phenolic content was expressed as milligrams of garlic acid equivalent (GAE) per milligram of extract.
Total flavonoid
The total flavonoid contents (TFCs) of the extract were determined using aluminum chloride calorimetric method as previously described (Tohidi et al., 2017) with some changes. Measurements were taken at an absorbance of 510 nm, and the TFC was presented in milligrams of quercetin equivalents (QEs) per milligram of the extract.
Bacteria and culture conditions
Twenty isolates of L. monocytogenes were used in this study. A reference strain F2365 (a gift from Food Safety Laboratory, Cornell University) (Nightingale et al., 2007) and 19 isolates from ready-to-eat food and food-processing environments were obtained from the Department of Food Technology, Prince of Songkla University, Thailand (Vongkamjan et al., 2015). All the bacterial cultures were stored in tryptic soy broth (TSB; Difco, Le Port de claix, France) supplemented with 40% glycerol and kept at −80°C.
Determination of the minimum inhibitory and minimum bactericidal concentrations
The minimum inhibitory concentration (MIC) of the extract was determined using the standard broth microdilution method (CLSI, 2015). Concentrations of the extract ranging from 16 to 1024 μg/mL sodium nitrite and sodium benzoate (4–128 mg/mL) were evaluated on selected isolates. The MIC values were recorded as the lowest concentration that completely inhibited the bacterial growth, and the minimum bactericidal concentration (MBC) values as the lowest concentrations that showed no growth on tryptic soy agar (TSA) plates after incubation for 24 h. All the experiments were set up in triplicates for two independent studies.
Time-kill assay
Bacterial inactivation kinetics of the extract was evaluated using the time-kill assay (Mitsuwan et al., 2017). The reduction in colony-forming unit (CFU) per milliliter was monitored after treatment with extract concentrations at 0, 4, 8, 12, 16, and 24 h. The 96-well polystyrene microtiter plate was used for this experiment, and all the experiments were performed in triplicate and recorded as mean ± standard deviation (SD).
Motility assay
The tested isolates and strain F2365 were evaluated for motility at 30°C and 37°C according to the method previously described (Borges et al., 2012; Vazquez-Armenta et al., 2018). The effect of the extract on the motility of highly motile isolates (PSU-KV-108, PSU-KV-116, and PSU-KV-122) and strain F2365 were evaluated at 30°C. Results were recorded as mean diameter of motility ± SD of triplicate values from two independent trials.
Effects of E. camaldulensis on L. monocytogenes biofilm formation
The effects of the extract on biofilm formation were determined in a 96-well microtiter plate, using the crystal violet assay as described (Du et al., 2018). Absorbance was read at 570 nm and values were recorded as mean ± SD of triplicate values.
Listeriolysin O inhibition assay
Hemolytic activity of L. monocytogenes was investigated following a method described previously (Sansano et al., 2017) with slight modifications. Overnight cultures grown in TSB at 37°C were harvested (4500 g, 10 min), and washed three times with TSB. Pelleted cells were resuspended in TSB with or without the extract to a final concentration of 106 CFU/mL. After 18-h incubation, the cultures were centrifuged at 12,000 g, 10 min, and the cell-free supernatant (CFS) was collected for hemolysis assay. About 3% red blood cells (RBC) were obtained as described (Upadhyay et al., 2012) and treated with the CFS. After 30-min incubation, the absorbance was read at 600 nm using microplate reader (EnSpire Multimode Plate Reader; PerkinElmer). Percentage hemolysis was calculated using the formula (Du et al., 2018):
where ODs , ODn , and ODp are absorbance values for sample, negative control, and positive control.
Membrane permeability assay
The effects of the extract on bacterial membrane integrity were evaluated on isolates PSV-KV-033, PSU-KV-039, PSU-KV-148, and strain F2365 by crystal violet uptake assay as described (Devi et al., 2010) with slight modifications. Cultures treated with 1% dimethyl sulfoxide (DMSO) were used as negative controls. Experiments were performed in triplicate for two independent trials and the crystal violet uptake was calculated as follows:
2,2-diphenyl-1-picrylhydrazyl assay
Stock solution (0.076 mM) of 2,2-diphenyl-1-picrylhydrazyl (DPPH) was prepared by dissolving 0.003 g crystalline powder in 100 mL of 95% methanol. Standard solution of trolox and ascorbic acid was also prepared by dissolving 1 mg of crystalline trolox and 1 mg ascorbic acid powder in 1 mL 95% methanol. The DPPH assay followed the modified method of Kharat and Mendhulkar (2016). Radical scavenging activities were read at an absorbance of 517 nm and the percentage inhibition was calculated using the formula;
where AB control is the absorbance of the negative control, AB test is the difference of the absorbance of samples and corresponding blanks. The concentration required to scavenge 50% of DPPH (IC50) was calculated from a regression line of percentage inhibition against concentrations. The experiment was conducted in triplicate for two independent trials.
ABTS radical scavenging activity
The ABTS [2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid)] was performed as described by Li et al. (2012), with some modifications. The ABTS solution was prepared by reacting equal volumes of 1.8 mg/mL aqueous ABTS and 0.66 mg/mL potassium persulfate (K2S2O8), and then storing in the dark for 12 h at room temperature. The ABTS scavenging effects were measured at 734 nm. Trolox and ascorbic acid were used as standards. Percentage scavenging activity was calculated and expressed as IC50, using the formula:
Ferric reducing antioxidant power assay
The ability of the test extracts to reduce ferric ions was measured using the ferric reducing antioxidant power (FRAP) assay as described by Benzie and Strain (1999). FRAP reagent was prepared by mixing 300 mM sodium acetate buffer (pH 3.6), 10.0 mM tripyridyltriazine solution, and 20.0 mM FeCl3·6H2O solution in a ratio of 10:1:1. The reaction was measured at an absorbance of 593 nm, and a standard curve was prepared using different concentrations of ascorbic acid. The results were expressed as micromolar ascorbic acid equivalent/mg extract. Analyses were performed in triplicate on each extract.
Statistical analysis
All experiments were performed in triplicate for two independent studies. The results were statistically analyzed using analysis of variance. Data were tested using the Scheffe statistical test (SPSS 20). A 95% confidence limit (p < 0.05) was adopted throughout the study.
Results and discussion
Antibacterial effects of E. camaldulensis on L. monocytogenes
This study presents the antibacterial effects of E. camaldulensis leaf extract on recalcitrant foodborne pathogenic L. monocytogenes. The results revealed good antilisterial effects with MIC ranging from 64 to 128 μg/mL and MBC from 256 to 512 μg/mL for all the tested isolates (Table 1). In contrast, the antilisterial activities of sodium nitrite and sodium benzoate used commercially as food preservatives were relatively high, with MIC values of 8–32 mg/mL and 32–64 mg/mL, respectively, with MBC >128 mg/mL. The antibacterial activity of these plants reflects the abundance of active phytochemicals such as terpenoids, flavonoids, phenols, saponin, and tannins (Ishnava et al., 2013). Microbial growth during food storage is a major cause of food spoilage (Petruzzi et al., 2017). Thus, application of antimicrobial extracts of E. camaldulensis as a natural preservative agent will retard spoilage, extend shelf-life, and improve food quality and safety. Plants from the family Myrtaceae, such as Rhodomyrtus tomentosa (Odedina et al., 2015), Myrtus communis (Amensour et al., 2010), and Psidium guajava (Olatunde et al., 2018) have been reported to exhibit antilisterial activities. The antibacterial activities of essential oil (Ghaffar et al., 2015) and flavonoid compounds (Takahashi et al., 2004) from Eucalyptus leaves have been reported previously. Ethanolic extracts of E. camaldulensis leaf exhibited antimicrobial activity against S. aureus (Nasr et al., 2018).
Antibacterial Activity of Eucalyptus camaldulensis Ethanolic leaf Extract on Listeria monocytogenes
The experiments were performed in the 96-well polystyrene microtiter plate, wells containing extract or salt concentrations were seeded with 100 μL of 106 CFU/mL bacterial suspension in MHB and incubated for 18–24 h at 37°C. For MBC, 10 μL aliquots from wells showing no visible growth were drop plated on TSA and incubated at 37°C for 24 h. The MBC values were recorded as the lowest concentrations that showed no growth on TSA plates. All experiments were set up in triplicates for two independent studies.
CFU, colony-forming unit; MIC, minimum inhibitory concentration; MBC, minimum bactericidal concentration; MHB, Mueller-Hinton broth; TSA, tryptic soy agar.
Time-kill kinetic of E. camaldulensis on L. monocytogenes
The time-dependent killing activity of E. camaldulensis extract against representative isolates from the food-processing environment (PSU-KV-032), ready-to-eat food (PSU-KV-120), and strain F2365 were demonstrated (Fig. 1). Time-kill assay revealed growth inhibitory effects after 4-h treatment of the bacteria with the extract. Approximately 2–3-log reduction in CFU/mL against all the tested food and environmental isolates was observed after challenging the pathogen with the extract at MIC for 6 and 8 h. The results suggested that the extract at MIC, 2MIC, and 4MIC were bactericidal to the test organisms, but at 1/2MIC, a bacteriostatic effect was observed. The effects of the extract on L. monocytogenes were both concentration and time dependent.
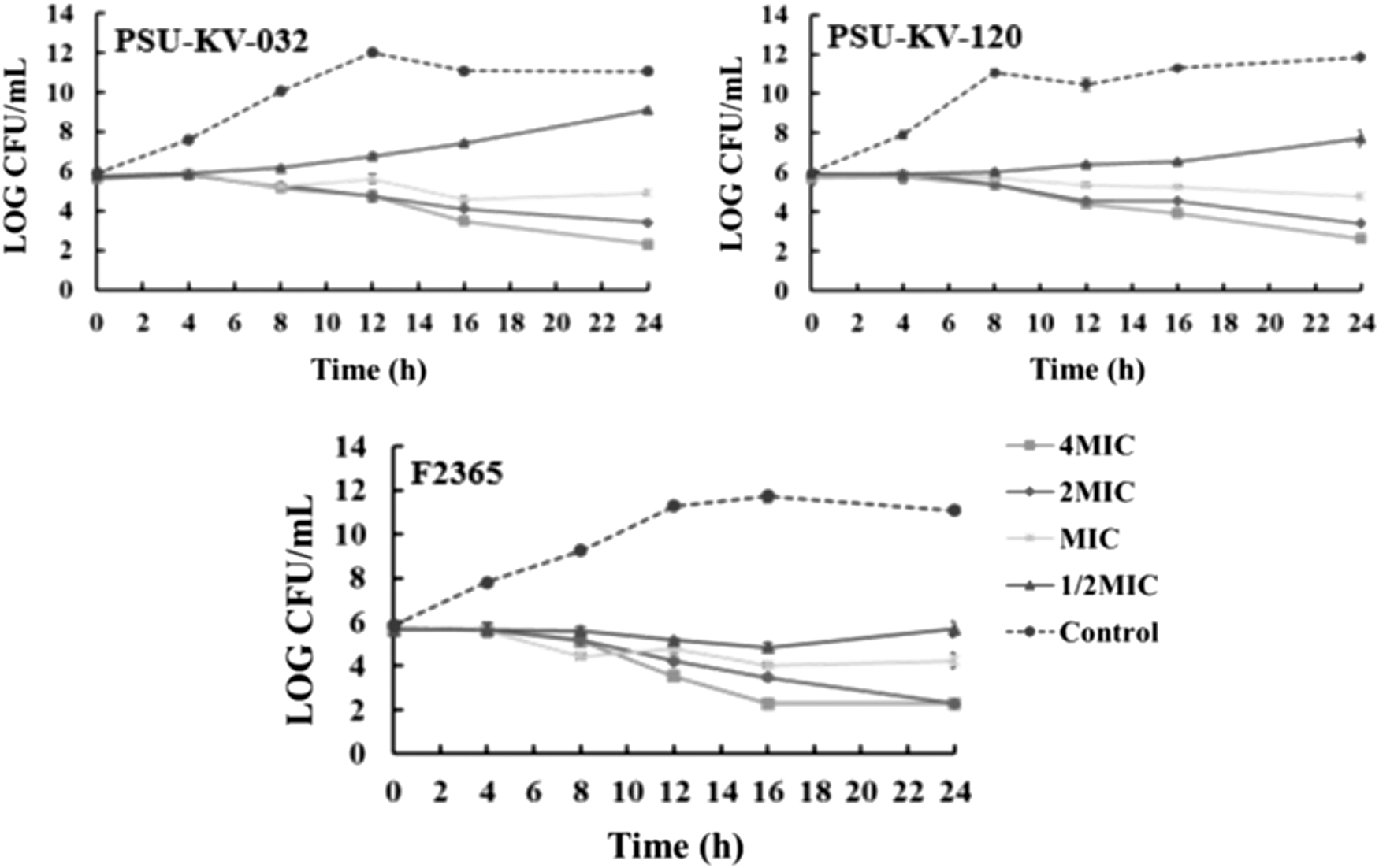
Time-kill curve of Eucalyptus camaldulensis extract on Listeria monocytogenes isolates performed using 96-well microtiter plate. Exponential phase cultures of PSU-KV-032, PSU-KV-120, and strain F2365 were adjusted to a 0.5 McFarland standard and 100 μL of 106 CFU/mL was treated with 100 μL of TSB supplemented with 4MIC, 2MIC, MIC, and 1/2MIC. The cultures were incubated statically at 37°C. One hundred microliters from individual concentrations was serially diluted at 0, 4, 8, 12, 16, and 24 h, and 10 μL of dilutions were plated on TSA. Viable bacteria cells were counted, and results were recorded as log CFU/mL. TSB supplement with 1% DMSO were maintained as negative control. The experiments were performed in triplicate and recorded as mean ± SD. CFU, colony-forming unit; MIC, minimum inhibitory concentration; TSA, tryptic soy agar; TSB, tryptic soy broth. SD, standard deviation; DMSO, dimethyl sulfoxide.
Effects of E. camaldulensis ethanolic extract on swarming motility of L. monocytogenes
L. monocytogenes exhibits a thermosensitive swarming motility mediated by the production of five to six peritrichous flagella (O'Neil and Marquis, 2006). These flagella consist of thousands of flagellin monomers that are modified by β-O-linked glycosylation (Schirm et al., 2004). Flagella formation and motility are thought to be responsible for L. monocytogenes biofilm formation and persistence within the food environment (Lemon et al., 2007). However, in contrast, Cabrita et al. (2015) noted that motility was not a critical feature for L. monocytogenes persistence in the food environment. The effects of inhibitory and subinhibitory concentrations of the extract on the motility of significantly motile isolates were determined at 30°C (Fig. 2). The results revealed a reduction in motility for most tested isolates. In addition, the tested concentrations significantly reduced the swarming motility of L. monocytogenes (p < 0.05), except 1/4MIC and 1/8MIC that showed no significant difference on isolate PSU-KV-116 (p < 0.05). The mechanism of Listeria motility inhibition by plant extracts and phenolic compounds is still unknown (Vazquez-Armenta et al., 2018). However, scanning electron microscopy revealed a loss of L. monocytogenes flagella corresponding to loss of motility and membrane damage at sublethal doses of olive leaf extract (Liu et al., 2017). A downregulation of flagellin-encoding gene (fliC) accompanied by loss of motility was reported in Escherichia coli after treatment with cranberry extract (Hidalgo et al., 2011).
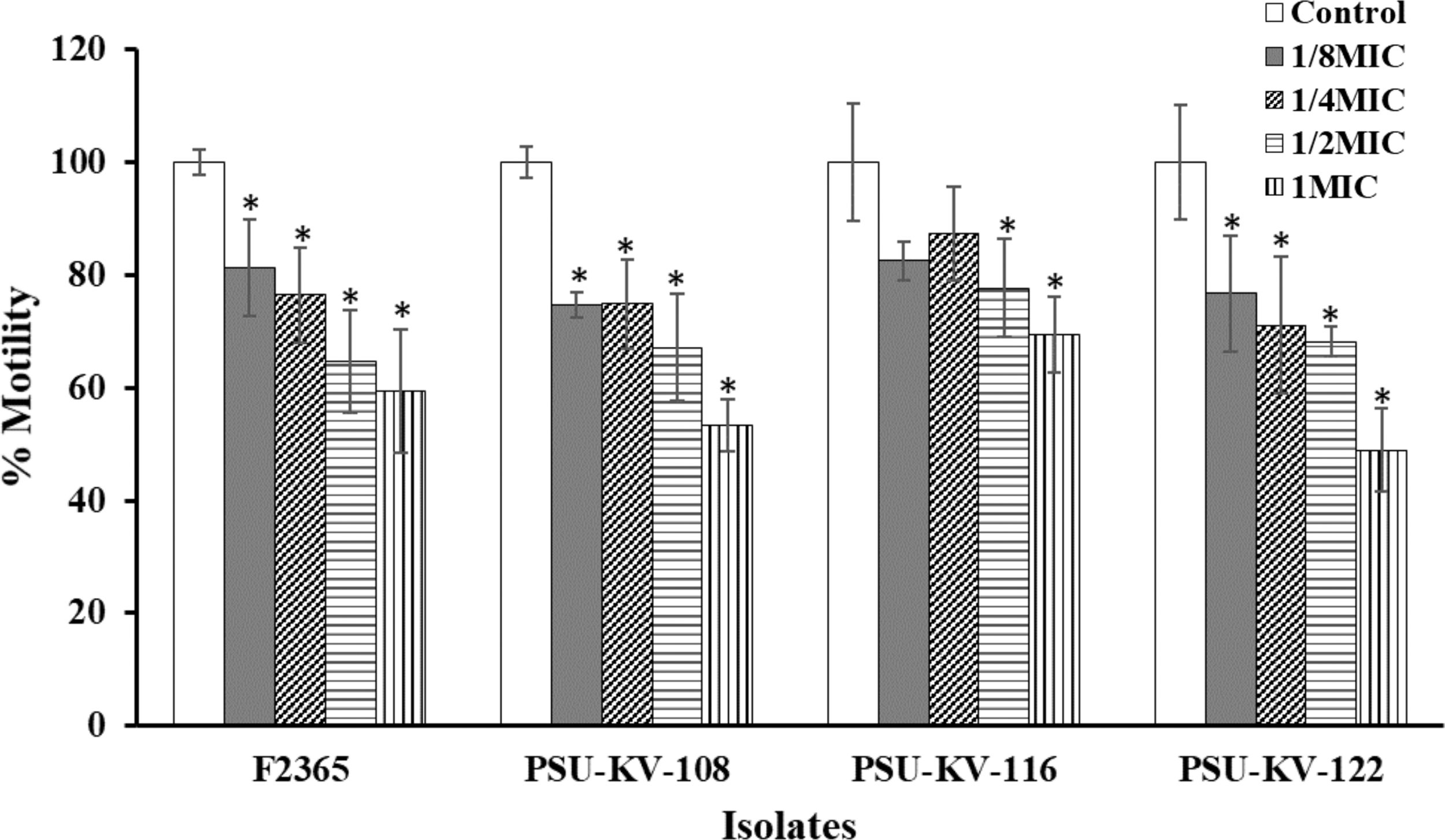
Inhibitory effect of E. camaldulensis extract on the motility of L. monocytogenes, measured on TSB +0.3% agar supplemented with inhibitory and subinhibitory concentrations of the extract. The plates were seeded with 10 μL of 106 CFU/mL bacterial suspension and incubated at test temperatures for 24 h. Motility medium supplemented with 1% DMSO was maintained as negative control. Result indicates values of the mean % motility ± SD for two independent experiments performed in triplicate. *Significant difference between treatment and control at p ≤ 0.05.
Effects of E. camaldulensis ethanolic extract on L. monocytogenes biofilm formation
Biofilm formation is an important microbial survival strategy. This sessile form of existence constitutes a food safety problem and serves as a source of recontamination of processed and finished products. Thus, biofilm inhibitory effect of sublethal concentrations of E. camaldulensis was further determined (Fig. 3). The results demonstrated that 1/2MIC and 1/4MIC inhibited biofilm formation. At 1/4MIC, biofilm formation for food isolate PSU-KV-148 and strain F2365 were significantly inhibited (p < 0.05). In contrast, no significant effect on isolates PSU-KV-033 and PSU-KV-039 was observed (p < 0.05). Previous researchers have suggested a correlation between flagella production, bacterial attachment, and biofilm formation (Lemon et al., 2007; Gorski et al., 2009). However, we observed that biofilm formation was higher at 37°C than 30°C (motility temperature) (Supplementary Figure S1). In addition, highly motile isolates did not produce a corresponding higher amount of biofilm (Supplementary Figure S2). The molecular mechanism governing biofilm production in L. monocytogenes remains uncertain (Schirm et al., 2004). However, population density-dependent cell–cell communication (quorum sensing) is thought to regulate biofilm formation (Rutherford and Bassler, 2012). Plant phytochemicals such as polyphenols and flavonoids inhibit biofilm by modulating bacterial cell–cell communication (Vikram et al., 2010), interfering with surface charge, hydrophobicity and motility (Monte et al., 2014), and downregulation of genes required for biofilm formation (Kim et al., 2016).
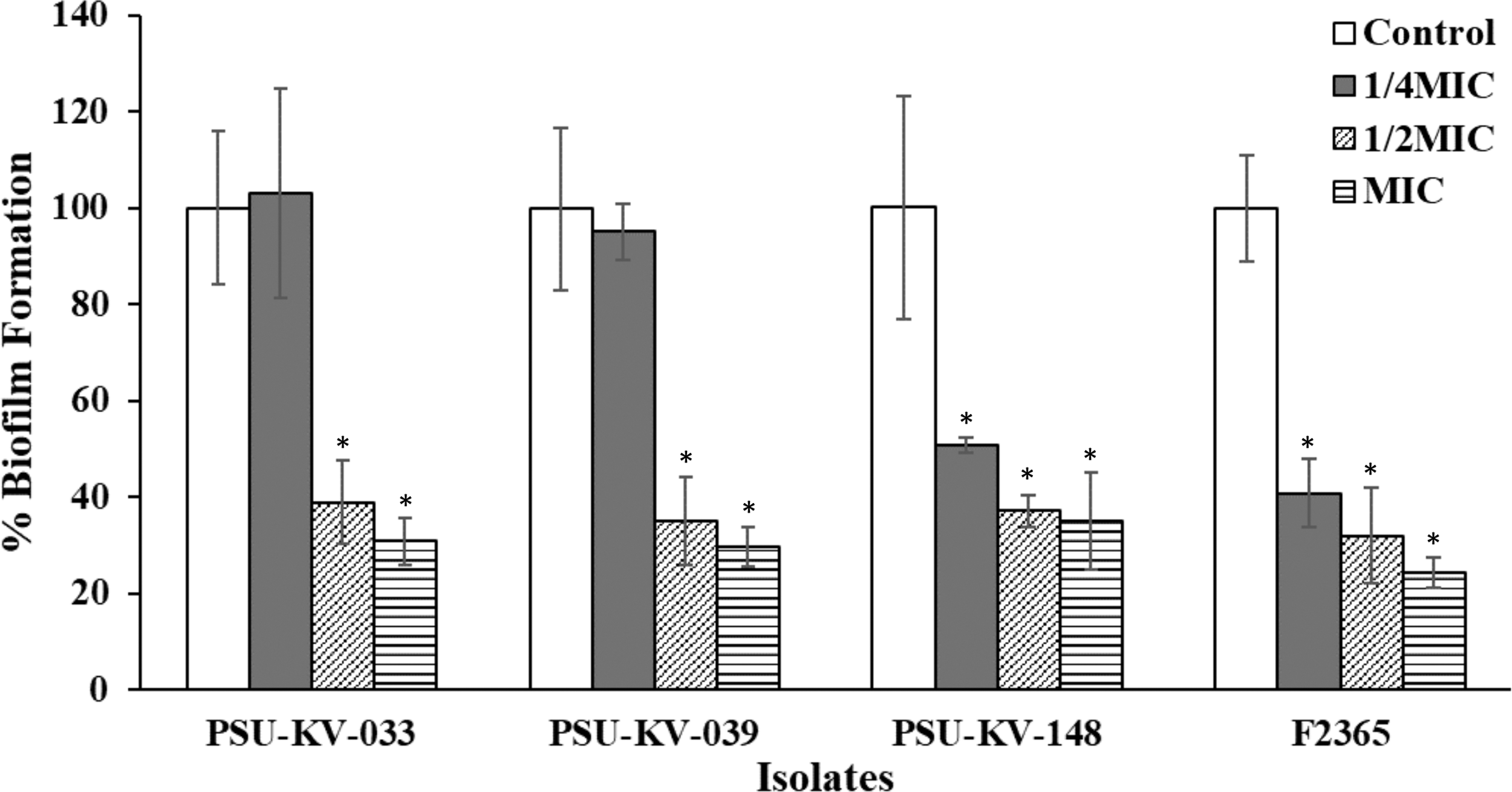
Biofilm inhibitory effects of subinhibitory concentrations of E. camaldulensis on L. monocytogenes after 24-h treatment. Overnight TSB cultures supplemented with 0.6% yeast extract were adjusted to 106 CFU/mL. One hundred milliliters of the 106 CFU/mL cultures was transferred into wells containing 100 μL of TSBYE supplemented with subinhibitory concentrations of the extract (MIC, 1/2MIC, and 1/4MIC). The plates were incubated at 37°C for 24 h. The effects of the extract on biofilm formation were determined by staining with 0.1% crystal violet. Untreated TSBYE cultures were maintained as negative control. Values indicate the mean ± SD for three independent experiments performed in triplicate. *Significant difference between treatment and control at p ≤ 0.05. MIC, minimum inhibitory concentration; TSBYE, tryptic soy broth yeast extract.
Effects of E. camaldulensis ethanolic extract on listeriolysin O production
Listeriolysin O is a cholesterol-dependent cytolysin, a family of pore-forming toxins commonly present in gram-positive bacteria. Listeriolysin O is a primary virulence factor of L. monocytogenes, necessary for intracellular survival and escapes from host acidic phagolysosomal compartments. In this study, listeriolysin O production ability of all the test isolates was evaluated. Eighty-five percent (17 isolates) showed 27–97% hemolysis on red blood cells after exposure for 30 min (data not given). The effect of subinhibitory concentrations of the extract on listeriolysin O production was further determined (Fig. 4). The results showed that subinhibitory concentrations of E. camaldulensis effectively inhibited listeriolysin O production after treatment for 18 h. MICs of the extract completely inhibited listeriolysin O production, probably because of growth inhibition at this concentration. Treatments indicated a consistent concentration-dependent effect that was significantly different from the control (p < 0.05). However, concentration of 1/16MIC showed no significant difference from the control against strain F2365. Previous researchers have reported listeriolysin O inhibitory activities of plant extracts and plant-derived compounds (Sansano et al., 2017; Du et al., 2018). However, a detailed understanding of the inactivation and inhibitory mechanism of listeriolysin O is still lacking. Repression of the L. monocytogenes listeriolysin O gene (hly) was observed after treatment with subinhibitory concentrations of tea tree oil (Liu et al., 2016). Fisetin, a natural flavonoid, was observed to inhibit hemolytic activity of listeriolysin O by competitively engaging loop 2 and loop 3 of listeriolysin O, thus preventing the binding of cholesterol and reduction of oligomerization (Wang et al., 2014).
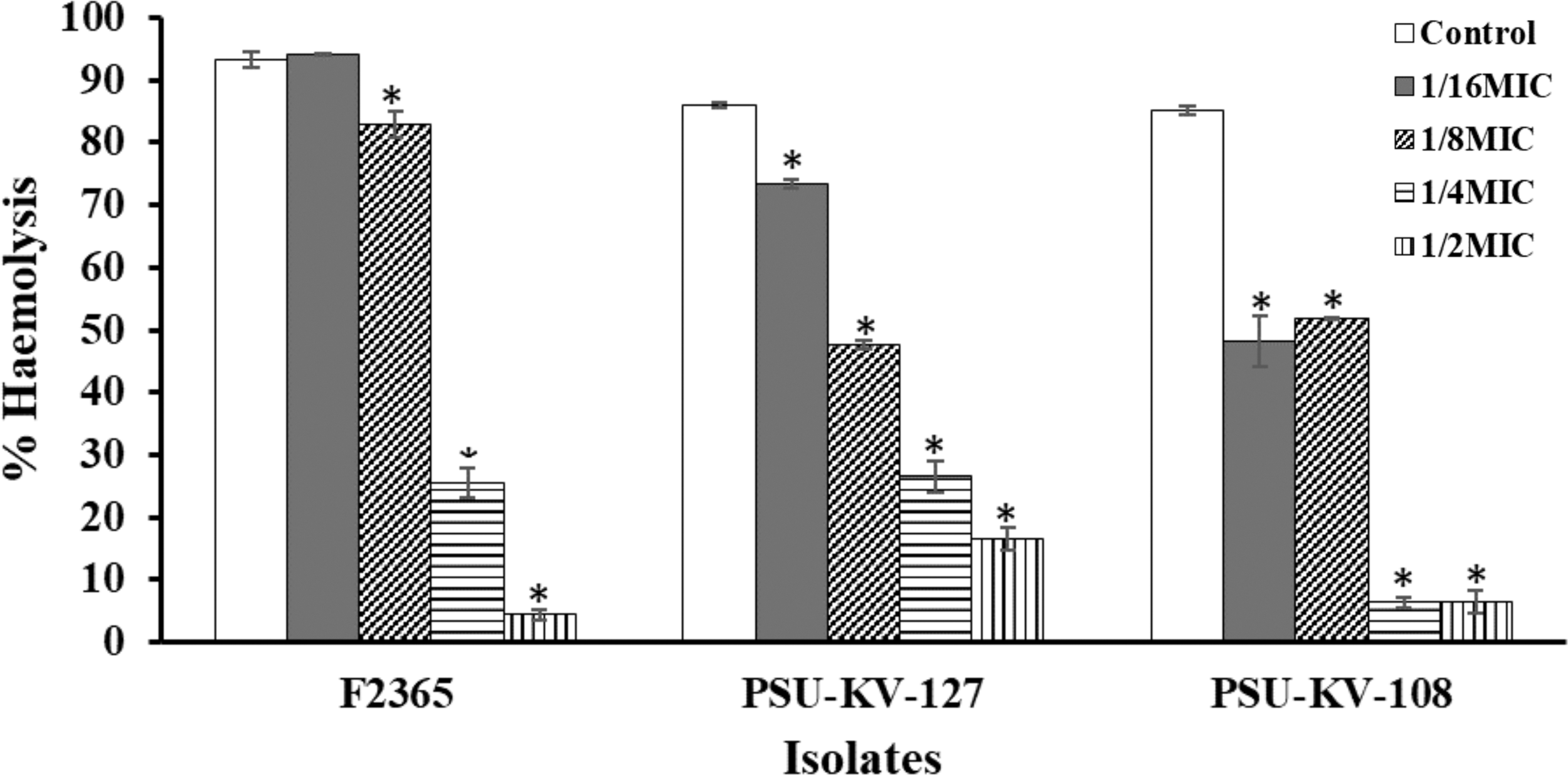
Effect of subinhibitory concentrations of E. camaldulensis on Listeriolysin O production. TSB cultures were supplemented with subinhibitory concentrations (1/2MIC, 1/4MIC, 1/8MIC, and 1/16MIC) of the extract and incubation for 18 h. The cultures were centrifuged at 12,000 g, 10 min, and the supernatant was used to treat 3% RBC (∼7.4 × 107 cells/mL). Hemolysis assay was conducted in microdilution plates by adding 100 μL CFS, 100 μL PBS, and 100 μL 3% RBC and incubating for 30 min. A positive control (100% hemolysis) 100 μL of 1% Triton X-100, 100 μL of PBS, and 100 μL of 3% RBC and negative control (0% hemolysis) 100 μL fresh TSB, 100 μL PBS, and 100 μL 3% RBC were included in the assay. Absorbance at 600 nm was read using microplate reader. Values were recorded as mean ± SD of triplicate values. *Significant difference between treatment and control at p < 0.05. RBC, red blood cells; CFS, cell-free supernatant; PBS, phosphate-buffered saline.
Effects of E. camaldulensis ethanolic extract on L. monocytogenes cell membrane permeability
Bacterial membrane serves as a permeable barrier that protects cytoplasmic contents from the damage that might arise from the influx of extracellular molecules. Alteration in membrane integrity might disrupt its permeability machinery, and thus lead to cell leakage. In this study, crystal violet uptake assay was used to evaluate the effect of the extract on the membrane integrity of L. monocytogenes after 30-min treatment. The results suggest that E. camaldulensis extract compromised the cell membrane integrity of L. monocytogenes at all concentrations (Fig. 5). Crystal violet uptake for all tested isolates increased significantly in a concentration-dependent manner. The increased uptake of crystal violet reflects an increase in cell membrane permeability that could have resulted from the disruptive effects of the extract.
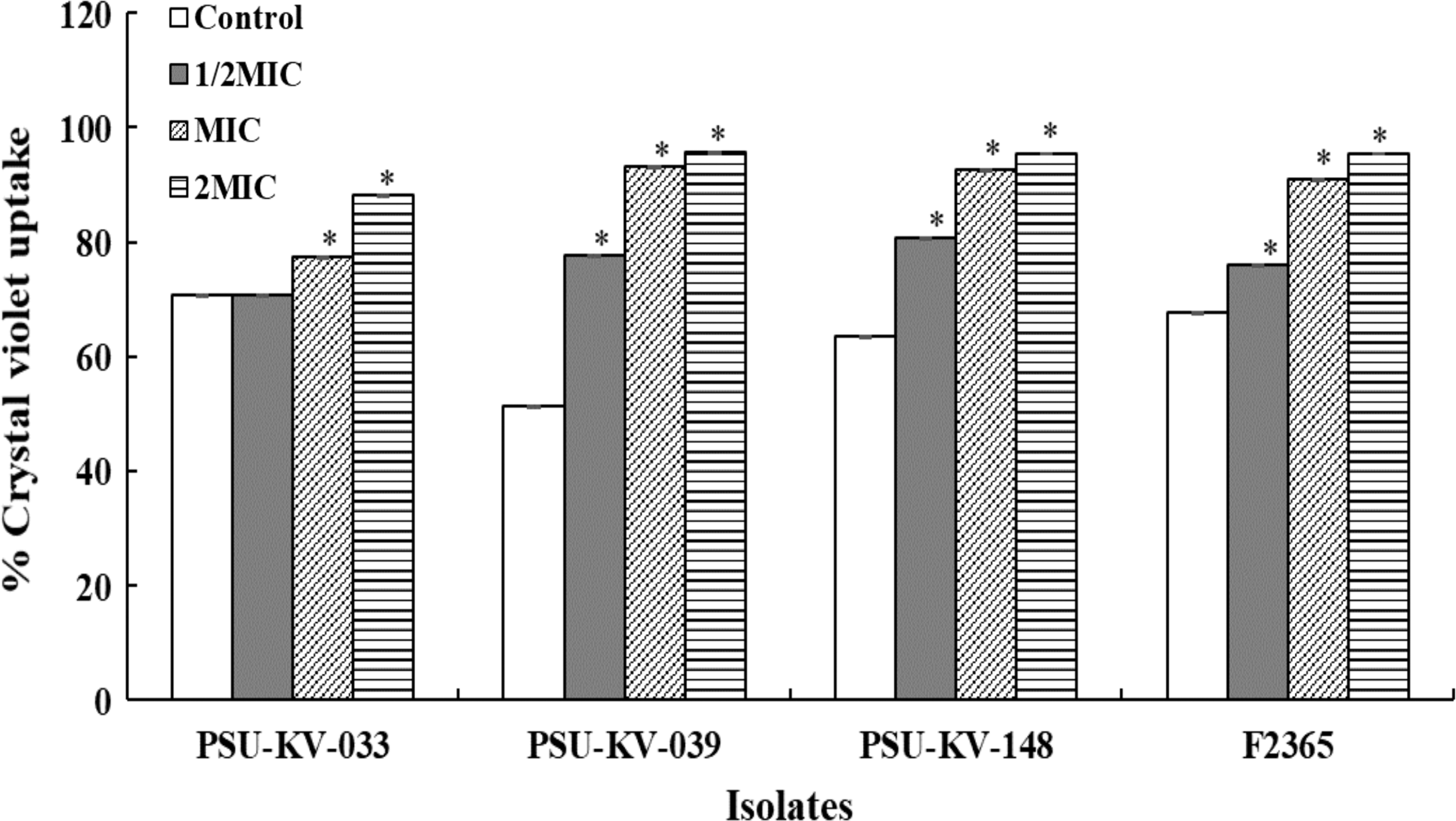
Effects of E. camaldulensis extract on membrane permeability of L. monocytogenes. Overnight cultures were harvested, washed, and resuspended in TSB supplemented with 2MIC, MIC, and 1/2MIC of extract to a final concentration of 1.5 × 108 CFU/mL. The bacterial suspensions were incubated for 30 min at 37°C. Cells were harvested at 8000 g, 5 min and the pellets were resuspended in PBS containing 10 μg/mL of crystal violet and incubated statically at 37°C for 10 min. Cell suspensions were centrifuged at 9500 g, 15 min, and the supernatants were collected and measured at OD590. The value indicates mean ± SD for three independent experiments performed in triplicate. *Significant difference between treatment and control at p ≤ 0.05.
Free radical scavenging activity of E. camaldulensis ethanol leaf extract
Free radical scavenging or quenching extends the shelf-life of food products. In this study, the free radical scavenging activities of E. camaldulensis was evaluated using the DPPH and ABTS assays (Table 2). The result showed an IC50 of 57.07 and 29.01 μg/mL for DPPH and ABTS, respectively. The IC50 of the E. camaldulensis methanolic leaf extract on DPPH was previously reported as 89.11 μg/mL (Ashraf et al., 2015) and 601.8 μg/mL and 1279.58 μg/mL for methanolic and ethanolic extracts, respectively (Upreti et al., 2018). ABTS assay demonstrated better activity with IC50 lower than ascorbic acid (40.52 μg/mL) and trolox (50.90 μg/mL). The low IC50 shown by ABTS might indicate the presence of lipophilic constituents not detected by the hydrophilic-based DPPH (Floegel et al., 2011; Kedare and Singh, 2011). Enzymatic oxidation resulting from the generation of free radical species is a major cause of food spoilage during storage (Petruzzi et al., 2017). Thus, the antioxidant results suggest that oxidative degradations such as lipid oxidation of food can be reduced by the applications of bioactive extracts of E. camaldulensis, leading to a delay of spoilage, extension of shelf-life, and maintenance of food quality and safety.
Antioxidant Profile of E. camaldulensis Ethanolic Leaf Extract
ABTS, 2,2′-azino-bis-3(ethylbenzothiazoline-6-sulfonic acid); NA, not applicable; GAE, garlic acid equivalent; QE, quercetin equivalents; DPPH, 2,2-diphenyl-1-picrylhydrazyl; FRAP, ferric reducing antioxidant power; AAE, ascorbic acid equivalent; IC50, 50% inhibitory concentration.
Total phenolic, TFCs, and ferric reducing antioxidant power
Eucalyptus spp. are rich sources of phenolic and flavonoid compounds with excellent antioxidant activities. Total phenolic and flavonoid contents of the extract measured using the Folin–Ciocalteu and AlCl3 calorimetric assays yielded values equivalent to 11.10 mg GAE/mg extract and 15.05 mg QE/mg extract, respectively. The total antioxidant contents of E. camaldulensis leaf extract estimated by FRAP test demonstrated high FRAP of 92.93 μM ascorbic acid equivalent (AAE)/mg extract. The antioxidant power of the extract might be because of the phenolic and flavonoid contents of the extract. FRAP of E. camaldulensis leaf was previously reported as 34.81 ± 0.67 mg GAE/g of plant extract for hexane extract, 15.02 ± 0.35 mg GAE/g of plant extract for methanol extract, and 9.54 ± 0.04 mg GAE/g of plant extract for chloroform extract (Ashraf et al., 2015).
Conclusions
Control of foodborne and spoilage microorganisms is a crucial aspect of food safety. This study provides evidence on the antibacterial activity of the ethanolic extract of E. camaldulensis against foodborne L. monocytogenes. The extract demonstrated good antivirulence potentials against the pathogen. Subinhibitory concentrations of the extract inhibited motility, listeriolysin O production, and biofilm formation. In addition, a membrane disruption effect was observed as demonstrated by the increase in dye uptake. Furthermore, the plant extract also demonstrated good antioxidant properties. Thus, E. camaldulensis ethanolic leaf extract might serve as an alternative source of antibacterial and antioxidant biopreservative agent against L. monocytogenes.
Footnotes
Acknowledgments
The study was funded by the Thailand's Education Hub for ASEAN Countries (Grant No. TEH-AC 013/2017) and TRF Senior Research Scholar (Grant No. RTA 6180006), the Thailand Research Fund.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.