
Abstract
Humans can contract anisakiasis by eating fish or squid containing live larvae of the third stage (L3) of the parasitic nematodes of the genus Anisakis, majorly from Anisakis simplex s.s. and Anisakis pegreffii, sibling species of the A. simplex s.l. complex. Most cases diagnosed molecularly are due to A. simplex s.s., although A. pegreffii has also been identified in human cases. Cathepsins are mostly lysosomal multifunctional cysteine proteases and can participate in the pathogenicity of parasites. Cathepsin B and L activities were investigated in the two sibling species of Anisakis mentioned. L3 and L4 of both species were collected during their in vitro development, and cathepsin activity was determined in the range of pH 4.0–8.5, using specific fluorogenic substrates. The activity detected with the substrate Z-FR-AMC (N-α-benzyloxycarbonyl-L-phenylalanyl-L-arginine-7-amido-4-methyl-coumarin) was identified as cathepsin L (optimum pH = 5.0, range 4.0–6.0, p < 0.001). Activity was highest in L3 freshly collected from fish, especially in A. simplex s.s., and decreased during development, which could be related to virulence, invasion of host tissues, and/or intracellular digestion. Cathepsin B-like activity was not identified with either of the substrates used (Z-RR-AMC [N-α-benzyloxycarbonyl-L-arginyl-L-arginine-7-amido-4-methyl-coumarin] and Z-FR-AMC). With Z-RR-AMC, cleaving activity was detected almost exclusively in L4 of A. simplex s.s. (p < 0.05) with optimum pH = 8.0 (range 7.0–8.5). Assays with class-specific protease inhibitors showed that this activity was mainly due to serine proteases [up to 90% inhibition with 4-(2-aminoethyl) benzenesulfonyl fluoride hydrochloride (AEBSF)], although metalloproteases (up to 40–45% inhibition with 1,10 phenanthroline) and slight cysteine protease activity (<15% inhibition with E64 [L-trans-epoxysuccinyl-leucylamido-(4-guanidino)-butane]; putative cathepsin B-like) were also detected. These results show differential serine protease activity between sibling Anisakis species, regulated by larval development, at least in A. simplex s.s. The higher cathepsin L and serine protease activities detected in this species could be related to its greater pathogenicity, reported in experimental animals, compared to that of A. pegreffii.
Introduction
Anisakiasis or anisakidosis is an infection caused by the larvae of the third stage (L3) of parasitic nematodes of the family Anisakidae. Humans can be infected on consuming fish or squid parasitized by these larvae and which is raw, undercooked or marinated, smoked, salted, etc. More than 20,000 cases have been described worldwide (Chai et al., 2005), perhaps underestimated (Bao et al., 2017), by Anisakis (>97% cases), Pseudoterranova (2–3%), Contracaecum, and Hysterothylacium. Although more than 90% of cases occur in Japan, anisakiasis also have been often reported in Asian countries such as South Korea (Lim et al., 2015) or European countries such as Spain or Italy (López-Serrano et al., 2000; Pampiglione et al., 2002).
The etiologic agents are usually the larvae of Anisakis simplex s.s. or Anisakis pegreffii, two species of the complex A. simplex s.l. Although cases involving the former appear more frequent in sympatric zones, implying greater pathogenicity, the lack of studies in humans advises caution, although studies on experimental animals seem to confirm this (Rello Yubero, 2003; Suzuki et al., 2010; Quiazon et al., 2011; Romero et al., 2013; Jeon and Kim, 2015).
Cathepsins are peptidases, usually from the papain family and of lysosomic origin, thus acting preferentially in an acidic medium. However, in contrast to the cysteine endopeptidases of vertebrates, the enzymes of helminths exhibit activity within a wide range of pH (pH 4.0–8.0) (Robinson et al., 2013). They are generally cysteine proteases although some aspartyl- or serine-type proteases have also been described.
Nematode cathepsins are involved in most of the functions performed by the proteases of parasites, including penetration of host tissues, immune response evasion, virulence, digestion, embryogenesis, moulting, and, particularly, intracellular digestion as a result of their lysosomic origin (Dalton et al., 1996; Hashmi et al., 2002; Guiliano et al., 2004; Robinson et al., 2008; Malagón et al., 2010, 2011, 2013). However, the cathepsins of parasites have not been sufficiently studied despite their being the key to the development of new chemotherapeutic treatments against parasitic nematodes or those causing plant diseases and also useful for diagnosis and development of vaccines (Britton and Murray, 2002; Sajid and McKerrow, 2002; Caffrey et al., 2013).
As cathepsins B- and L-like are those most frequently described in nematodes, the aim of the present study was to detect and partially characterize these two types in the two species of Anisakis, which are the most common etiologic agents of anisakiasis, while determining differential characteristics associated with each species.
Materials and Methods
Sample collection and culture in vitro
Anisakis L3 type I were collected from blue whiting (Micromesistius poutassou) from Spanish ports located on the Cantabrian Sea (Ondarroa, Bay of Biscay) and the western Mediterranean Sea (Villajoyosa and Gandía, eastern Spanish coast). The fish were transported and immediately processed to collect the Anisakis larvae (Molina-Fernández et al., 2018). Briefly, the larvae were collected at that time for the L3 sample from fish (L3–0h) and frozen at −20°C until use. Meanwhile, new L3 batches were prepared and cultured as described by Iglesias et al. (1997, 2001). The larvae were removed from the culture at different development times: at 24 h (L3–24h); at 24 h after moulting to L4 (L4–24h), and after 14 days of culture (L4–14d, 10 days after moulting to L4). After harvesting they were frozen at −20°C until use.
To obtain a sufficient sample of each of the species investigated, the fish were collected from two different geographical areas with predominance of one or other species, according to previous studies (Martín-Sánchez et al., 2005; Mattiucci et al., 2018; Molina-Fernández et al., 2018). Thus, fish from Ondarroa (northern Spain, zone FAO VIIIc) were used for the collection of larvae of A. simplex s.s. and those from Villajoyosa and Gandía (eastern Spain, zone FAO 37.1.1) for larvae of A. pegreffii.
Genetic identification
Genetic identification was carried out as described by Molina-Fernández et al. (2015, 2018). After larval DNA extraction, a polymerase chain reaction–restriction fragment length polymorphism (PCR-RFLP) of the ribosomal DNA (rDNA) region ITS1–5.8S-ITS2 was performed using the primers NC5 (forward) and NC2 (reverse) described by Zhu et al. (1998). RFLP was performed independently with two restriction enzymes, TaqI and HinfI FastDigest (Thermo Scientific). The band pattern generated was visualized by 3% agarose gel electrophoresis and compared to a control for each species to be identified according to D'Amelio et al. (2000) and Abollo et al. (2003). The larvae that showed a mixed banding pattern between A. simplex s.s. and A. pegreffii with one or other restriction enzyme were considered as L3 type I recombinant genotype larvae.
Preparation of protein extracts
Larval extracts were prepared extemporaneously in tris-HCl buffer at pH 7.8, with 20% glycerin w/v to stabilize the proteins and prevent their degradation. The larvae were homogenized by mechanical means and immersion in liquid nitrogen. Once homogenized, the crude extract was centrifuged at 4°C and 19,000 × g for 20 min (Malagón et al., 2010, 2011). The resulting supernatant was transferred to an Eppendorf and kept on ice until its use as a source of enzymatic activity, its protein concentration being determined (Bradford, 1976).
Cathepsin assays
For the determination of the cathepsin L- and B-like activity of the soluble extract, the procedure of Malagón et al. (2010) was followed. The activity was tested in the pH range of 4.0–8.5 with increments of 0.5. To achieve a pH-activity profile, it was decided to use a single buffer for the entire pH range. This buffer was tris-maleic 0.2 M, adjusting the pH with HCl or NaOH depending on the case, with 2 mM dithiothreitol (DTT). The maximum ionic strength of the buffer was equivalent to 0.6 M, so NaCl was added to equalize the ionic strength at the different pHs, when necessary.
Fluorogenic substrates Z-FR-AMC (N-α-benzyloxycarbonyl-L-phenylalanyl-L-arginine-7-amido-4-methyl-coumarin), to determine cathepsin B- and L-like activity, and Z-RR-AMC (N-α-benzyloxycarbonyl-L-arginyl-L-arginine-7-amido-4-methyl-coumarin) to determine cathepsin B-like activity were used. These substrates show fluorescence when AMC becomes free as a consequence of the hydrolysis of the Arg-AMC bond. The emitted fluorescence was detected with a fluorometer with λex = 355 nm and λem = 460 nm. Before use, the substrates were dissolved in DMSO (dimethyl sulfoxide) at 10 mM and frozen at −20°C. The final concentration of DMSO in well was 1% in all the assays. The measurements were made in black microplates, and each well contained a volume (final concentration) of 100 μL of tris-maleic buffer (100 mM) with DTT (1 mM), 10 μL of extract (10 μg of protein), 30 μL of substrate (15 μM), and bidistilled water to complete a final volume of 200 μL. The reaction was initiated by the addition of the substrate. The measurements were made every 60 s for 60 min and, to standardize the process, the most stable zone of the curve was selected for each test, corresponding to 10–30 min.
For the inhibition assays, 0.01 mM E64 [L-trans-epoxysuccinyl-leucylamido-(4-guanidino)-butane], an irreversible inhibitor of cysteine proteases, was added to the reaction mixture. When this did not inhibit 100% activity, pepstatin A (0.02 mM) for aspartic proteases, 1,10-phenanthroline (2 mM) for metalloproteases, and 4-(2-aminoethyl) benzenesulfonyl fluoride hydrochloride (AEBSF, 2 mM) for serine proteases were also used as class-specific inhibitors. Enzyme activity was expressed as a variation (Δ) of fluorescence relative units (FRU) × min−1 × mg−1 protein.
Statistical analysis
The software SPSS 22.0 for Windows was used for the study. As the residuals of the dependent variable did not follow a normal distribution in any case using the Shapiro–Wilk test, the nonparametric Kruskal–Wallis test was performed. The slope of activity for each substrate was used as a dependent quantitative variable, and the variables “larval stage,” “species,” and “pH” were independent. Since it is a weak method, multiple post hoc comparisons were made by pairs when p < 0.1, using the Mann–Whitney U test with the Bonferroni correction. The significance level was set at 0.05.
Results and Discussion
Genetic identification
A total of 101 larvae of Anisakis type I from blue whiting from the port of Ondarroa (northern Spain) were analyzed using the PCR-RFLP technique, identifying 89.1% as A. simplex s.s., 3.0% as A. pegreffii, and 7.9% as recombinants of the two species. A further 55 larvae of Anisakis type I from the Mediterranean ports of Villajoyosa and Gandía (eastern Spain) were identified as 90.9% A. pegreffii, 1.8% A. simplex s.s., and 7.3% as recombinants. These data coincide with the known distribution of these species, A. pegreffii being more prevalent in the western Mediterranean and A. simplex s.s. in the northeastern Atlantic, with sympatry to the south and west of the Iberian Peninsula, although these zones were not sampled in this study (Martín-Sánchez et al., 2005; Mattiucci et al., 2018; Molina-Fernández et al., 2018, 2019).
Enzymatic activity
The superfamily of papain-like cysteine proteases, to which cathepsins B and L belong, is the best-described group of proteases and is regulated during helminth development (Robinson et al., 2008). Although mainly lysosomic, they have also been detected in the nucleus and cytosol and are secreted into the extracellular medium (Kirschke, 2013).
In the present study it was observed that the pH ranges in which activity of the extracts of A. simplex s.s. and A. pegreffii was detected did not overlap when a profile of activity-pH was carried out with the two substrates used. With substrate Z-FR-AMC, which is optimal for cleaving by cathepsins L, although it may also be cleaved by cathepsins B (Robinson et al., 2013), greater activity was observed at acidic pH, with statistically significant differences between pH values (p < 0.001), with pH 5.0 being the most favorable (Fig. 1).

Profile of cathepsin L-like activity measured by Z-FR-AMC cleavage in Anisakis spp. during its development in vitro, depending on pH. Each point is the mean of two to three experiments in triplicate. Upper panel: Anisakis simplex s.s. Bottom panel: Anisakis pegreffii. Z-FR-AMC, N-α-benzyloxycarbonyl-L-phenylalanyl-L-arginine-7-amido-4-methyl-coumarin.
The cleaving of substrate Z-FR-AMC occurred between pHs 4.0 and 6.0 (maximum at 4.5–5.5) and was almost undetectable at higher pH, always being inhibited by E64, a specific cysteine protease inhibitor. When considered together with the observed lack of fluorescence with substrate Z-RR-AMC (or very low levels not inhibited by E64) at this pH range, it must be supposed that cathepsin B activity is not measured with Z-FR-AMC—as reported by Dalton et al. (1996) for Schistosoma mansoni and by Malagón et al. (2010) for Hysterothylacium aduncum, suggesting that all the fluorescence detected in this pH range results from cathepsin L-like activity in the species of Anisakis studied.
The evolution of the activity during development varied according to species. In A. simplex s.s. maximum activity was detected in L3–0h, decreasing gradually in each developmental stage (p < 0.006). In A. pegreffii, although showing the same trend, there was an upsurge in activity in L4–24h, which then decreased dramatically to a value 14 times lower in L4–14d (Figs. 1 and 2). Despite this differential behavior, comparison of the different developmental stages between the two different species only showed significant differences between the most developed stage L4–14d (p < 0.005), although, overall, A. simplex activity was greater compared with A. pegreffii (p = 0.06). The highest activity in infective L3–0h may be related to the greater virulence observed in A. simplex s.s. when involved in the processes taking place during host tissue invasion, as occurs in other helminths (Stack et al., 2008; Xue et al., 2019), since, in this stage, the larva is prepared to invade either another paratenic host or its definitive host.
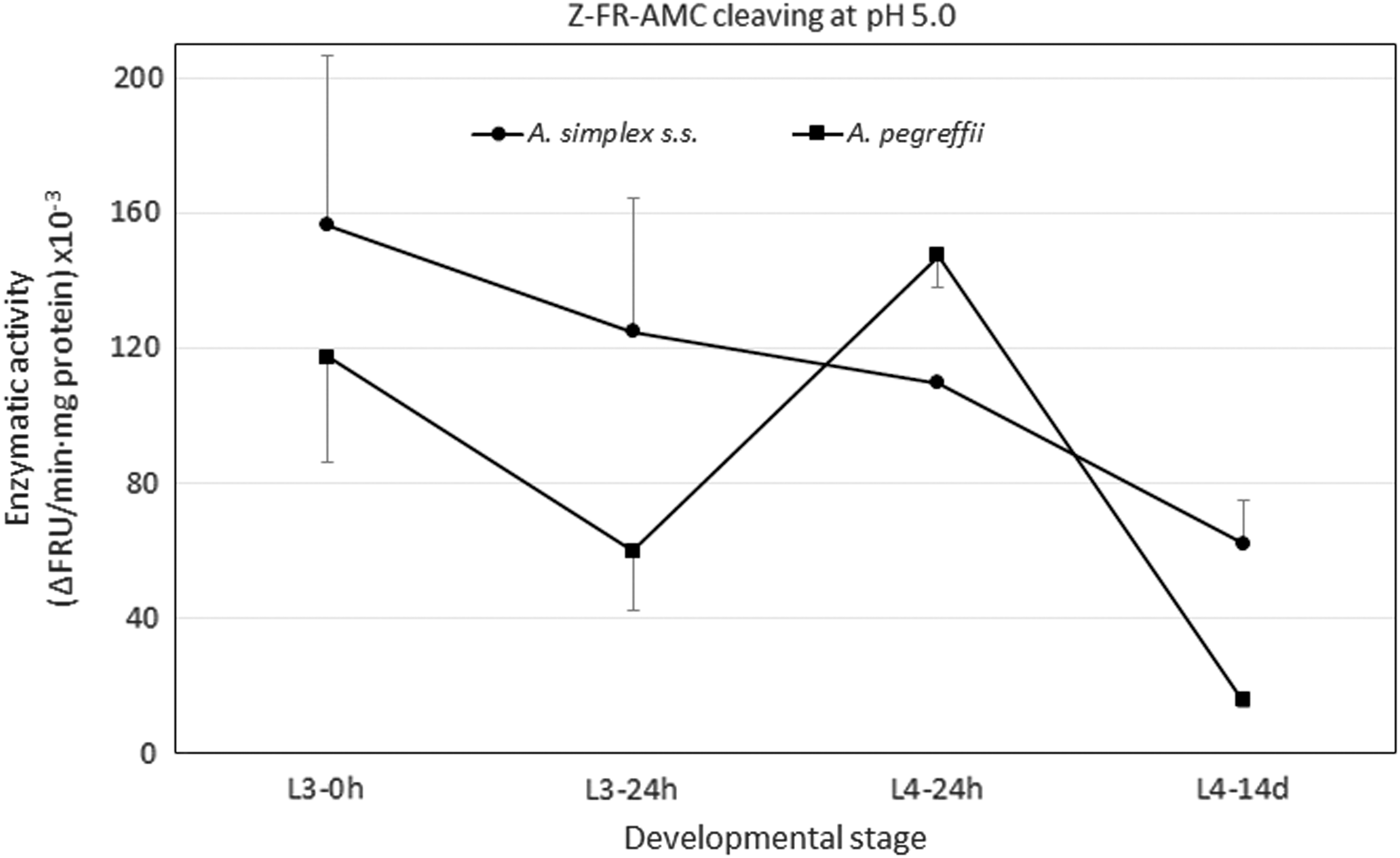
Profile of cathepsin L-like activity as measured by cleavage of substrate Z-FR-AMC at pH 5.0 in Anisakis simplex s.s. (•) and in Anisakis pegreffii (▪) during its in vitro development. Each point is the mean ± standard deviation of three experiments in triplicate.
However, it may also be related to intracellular digestion since, as several authors have suggested, L3 of Anisakis, unlike L4, are not able to ingest food through the digestive system but must obtain nutrients from the extracellular medium through the cuticle (Yasuraoka et al., 1967; Sommerville and Davey, 1976; Iglesias et al., 1997; Dávila et al., 2006). Recently moulted L4 clearly showed greater activity than those which had undergone a longer development time (Fig. 2), which may be related either to the remodeling of the cuticle, as observed in the filarial nematode Brugia pahangi (Guiliano et al., 2004), or to adaptation to a new acidic habitat in the stomach chambers of cetaceans (definitive hosts).
Maximum activity for cathepsins L with substrate Z-FR-AMC was at around pH 5 for the two species studied, as in other nematodes from the same superfamily Ascaridoidea such as the infective larva of Toxocara canis (Loukas et al., 1998) and H. aduncum (pH 5–5.5), at least in L3, L4, and adults (Malagón et al., 2010). As these cathepsins are usually of lysosomic origin, their optimal pH is generally 4.5–6.0 (Sajid and McKerrow, 2002; Malagón et al., 2010) although, as mentioned previously, they are also usually active and stable at neutral and even alkaline pH values, in contrast to those of mammals.
In this case, their exclusively acidic range of activity would imply their involvement in digestive processes (intracellular and/or intestinal), as reported for H. aduncum (Malagón et al., 2010), and perhaps also in processes related to attachment and moulting in an acidic medium associated with the gastric wall of the definitive host. Their relationship with moulting in nematodes has already been established and is considered a conserved function due to the high level of homology within this type of cathepsin (Britton, 2013). In addition, Xue et al. (2019) have linked the differential expression of cathepsin L genes with the development and pathogenicity of the nematode Bursaphelenchus xylophilus.
With substrate Z-RR-AMC, which is specifically cleaved by cathepsins B-like, activity was detected in both species within the range pH 5.0–8.5, with this activity concentrated within pH 7.0–8.5. Figures 3 and 4 show a good view of the effect of substrate for cathepsin B-like activity results in both species, being very low particularly in A. pegreffii. However, note that L4s showed notably higher activity (p < 0.05) with a maximum at pH 8.0 (Figs. 3 and 4). This appears to coincide with H. aduncum, in which a cathepsin B-like has been found with an optimum pH of 7.5 for cleaving Z-RR-AMC (Malagón et al., 2010). However, when the inhibition tests were carried out to determine the activity type of the Anisakis extracts, there was only a slight inhibition (<15%) with E64 under our experimental conditions, which shows that only a minimum part of this activity should be of a cysteine protease and, therefore, mostly it is not cathepsin B-like.
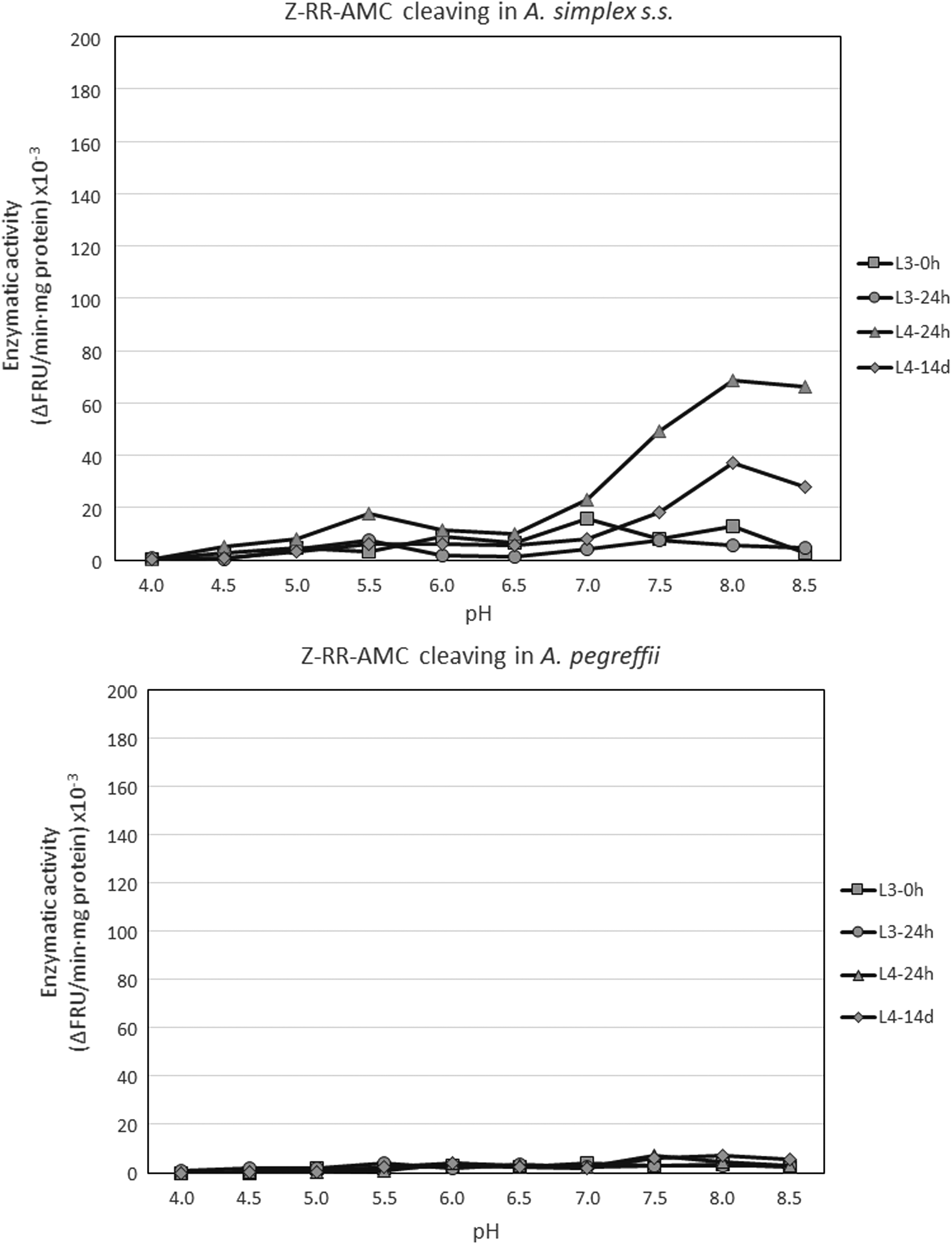
Profile of enzymatic activity measured by cleavage of Z-RR-AMC in Anisakis spp. during its development in vitro, depending on pH. Each point is the mean of two to three experiments in triplicate. Upper panel: Anisakis simplex s.s. Bottom panel: Anisakis pegreffii. Z-RR-AMC, N-α-benzyloxycarbonyl-L-arginyl-L-arginine-7-amido-4-methyl-coumarin.
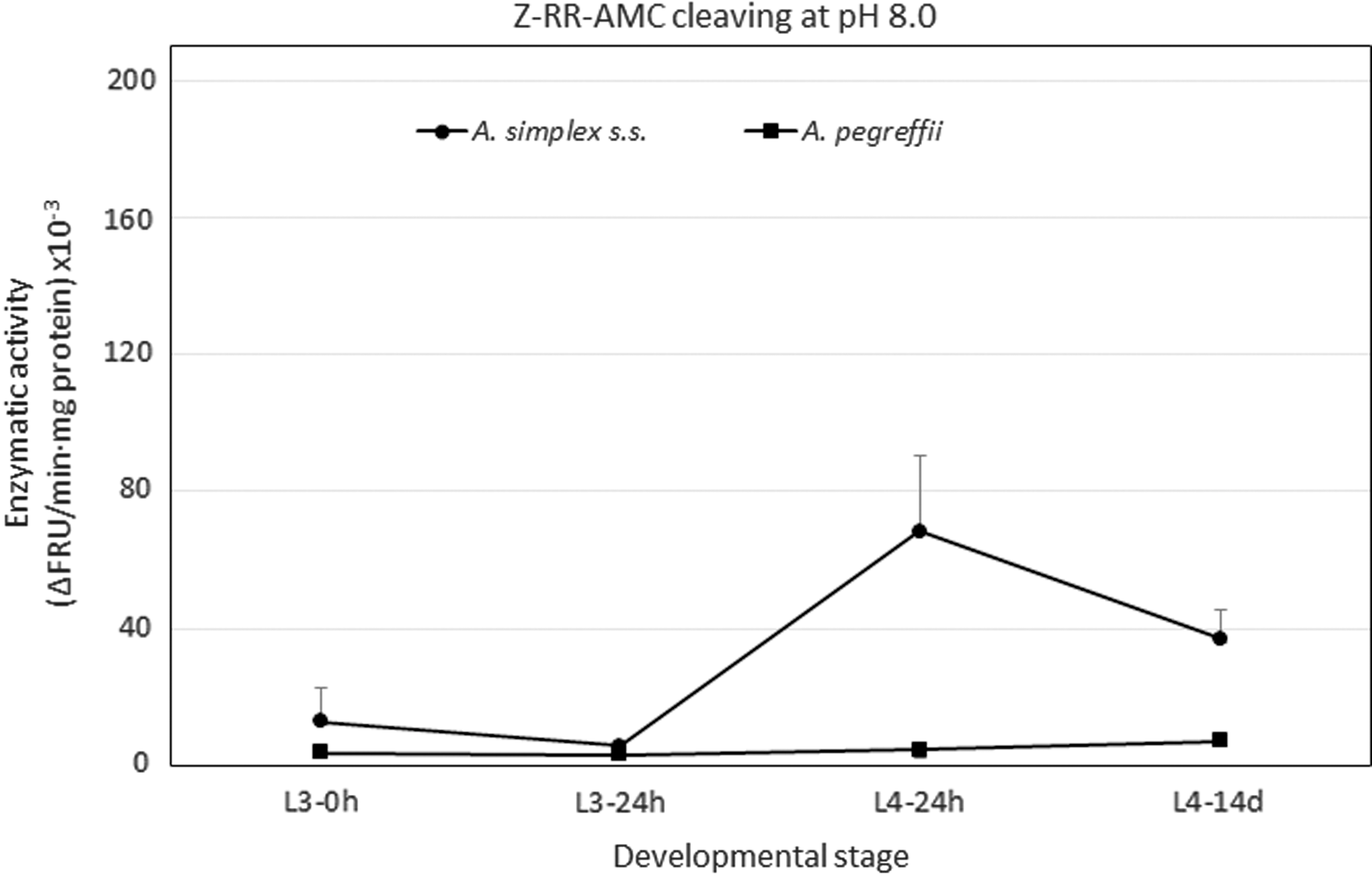
Profile of enzymatic activity measured by cleavage of substrate Z-RR-AMC at pH 8.0 in Anisakis simplex s.s. (•) and in Anisakis pegreffii (▪) during its in vitro development. Each point is the mean ± standard deviation of three experiments in triplicate.
Assays with other class-specific inhibitors revealed inhibition of up to 90% with AEBSF and 40–45% with 1,10-phenanthroline, showing that the activity detected was mainly due to serine proteases, with some participation by metalloproteases. Serine protease activity has been detected and identified in the excretory–secretory products of L3 of A. simplex with optimum pH of 7.5 (Matthews, 1982, 1984; Sakanari and McKerrow, 1990). Later, Morris and Sakanari (1994) isolated, purified, and characterized it as a trypsin-like serine protease, 89% homologous with pig trypsin, and able to cleave both Z-RR-AMC and Z-FR-AMC, the former more efficiently, which would explain the detection of activity with the former substrate and not with the latter. These authors reported that CaCl2 was necessary for the enzyme's stability but did not improve its activity.
In the present study, the addition of CaCl2 20 mM reduced activity by 60–90% (results not shown). Although the cleaving of Z-RR-AMC may be at least partially due to this enzyme, the very low activity of L3 makes this idea questionable. This enzyme may have been preferentially secreted during the L3 stage, possibly to carry out extracorporeal digestion (Buzzell and Sommerville, 1985), which has been observed in nematodes (Feng et al., 2007), and later incorporate the resulting end products of digestion through the cuticle. This could explain the low level of activity detected in the somatic extracts from this stage. However, as L4 are now able to ingest food orally it would not need to secrete the enzyme and it could be accumulated for use in intestinal digestion. Of course, it may be another different serine-protease which appears to express itself differentially in the L4 stage of A. simplex s.s. and is almost undetectable under our experimental conditions in A. pegreffii.
Morris and Sakanari (1994) succeeded in partially characterizing a second serine protease in the somatic extracts of L3 of A. simplex, which was 85% homologous to a bacterial capable of degrading tissues. It should also be noted that Molina-Fernández et al. (2019) found a significantly greater proteolytic activity by serine proteases in all stages of A. simplex s.s., developed at 37°C, the same temperature as in the definitive host, than in A. pegreffii, although the opposite occurred in L3 collected from the intermediate/paratenic fish poikilotherm host. In addition, Cavallero et al. (2018) reported a greater presence of trypsin-like serine protease transcripts in A. simplex s.s. than in A. pegreffii, although with the proviso that the procedure followed may have been more efficient in the former than in the latter. Furthermore, Jasmer et al. (2015) reported the low expression of cathepsin B-like cysteine peptidases among the peptidases in the intestine of adult females of Ascaris suum, suggesting their possibly scant contribution to nutrient digestion. On the other hand, the lack of activity against the substrate Z-RR-AMC, used for the detection of cathepsins B-like, is not uncommon in nematodes, as in Dirofilaria immitis (Richer et al., 1992) or Ancylostoma caninum (Dalton et al., 1994). In Caenorhabditis elegans, CPR-6, a cathepsin B-like is almost not expressed in the larval stages and overexpressed in adults, showing 70% identity with that of A. suum (Britton, 2013). Consequently, this type of activity cannot be discounted in Anisakis.
Proteases in general and cathepsins in particular can be regarded as potential therapeutic targets in helminths due to their role in development, survival, and pathogenicity for the host (Xue et al., 2019). In fact, some proteases, including cathepsins, are currently being studied with a view to their use in experimental vaccines against trematodes, such as Fasciola hepatica or S. mansoni, or gastrointestinal nematodes such as Haemonchus contortus or Ostertagia ostertagi in animals and against the hookworms in humans, with encouraging results so far (Knox, 2012; Hotez et al., 2013; Figueiredo et al., 2015).
Conclusions
A cathepsin L-like activity has been detected in the two sibling species of the complex A. simplex s.l.. The activity of the L3 of A. simplex s.s. is higher than A. pegreffii L3, which could be related to the higher pathogenicity of the former, and it seems also to be involved in the digestion of nutrients. In addition, a cathepsin B-like specific substrate is mostly processed by serine protease activity, which has been detected to be significantly higher in A. simplex s.s. than in A. pegreffii; it could be related to the higher pathogenicity of the former.
Footnotes
Acknowledgments
The authors are grateful to Dr. Manuel Díaz López, from Department of Biology and Geology, University of Almería, for his invaluable advice, to Dr. Miguel Romero Pérez from Department of Pharmacology, Faculty of Pharmacy, University of Granada, for his help with fluorimetry, and to Esperanza Díaz Fernández for her technical assistance. Translation to English was by Robert Abrahams, BSc. This work has been funded by the Agencia Estatal de Investigación (Spanish State Research Agency) and European Regional Development Fund (ERDF), grant number CGL2013-47725-P.
Disclosure Statement
No competing financial interests exist.