
Abstract
Zoonotic pathogen Escherichia albertii has been identified as the cause of several human disease outbreaks; however, factors such as the general symptoms and incubation period of E. albertii infection have yet to be defined. Therefore, we aimed to determine the unique aspects of E. albertii outbreaks in Japan and to examine the genetic characteristics of the causative pathogen. We studied all known E. albertii outbreaks that occurred in Japan up until 2015, which consisted of five confirmed outbreaks and one putative outbreak (Outbreaks 1–6). Outbreaks were re-examined based on personal communications between researchers in prefectural and municipal public health institutes, and through examination of any published study conducted at the time. Draft genome sequences of outbreak-associated E. albertii isolates were also generated. The most common symptom displayed by patients across the six episodes was watery diarrhea (>80%), followed by abdominal pain (50–84%) and fever (37.0–39.5°C) (26–44%). The estimated average incubation period of E. albertii infection was 12–24 h. We assumed that most of the outbreaks were foodborne or waterborne, with restaurant foods, restaurant water, and boxed lunches being the suspected transmission vehicles. Three of the six outbreak-associated E. albertii isolates possessed intact ETT2 regions, while the remaining isolates contained disrupted ETT2-encoding genes. Virulence gene screening revealed that more than half (44/70) of the tested genes were present in all 5 strains examined, and that each of the strains contained more than 1 gene from 14 out of the 21 groups of virulence genes examined in this study. The five E. albertii strains were classified into four of the five known phylogroups. Therefore, we determined that multiple E. albertii genotypes in Japan have the potential to cause outbreaks of diarrhea, abdominal pain, and/or fever following infection of a human host.
Introduction
Several outbreaks of gastrointestinal illness caused by Escherichia albertii, a pathogen first described in 2003 (Huys et al., 2003), have been reported (Sharma et al., 2007; Ooka et al., 2013; Asoshima et al., 2014; Murakami et al., 2014a). As such, key aspects of E. albertii outbreaks need to be recorded. From a public health perspective, it is important to establish the common symptoms and incubation period of E. albertii infection because this information may help to identify a causative agent in cases of foodborne disease (Centers for Disease Control and Prevention, 2015). Likewise, the unique aspects of E. albertii outbreaks should be documented to aid in disease prevention. However, the common symptoms, incubation period, and specific vehicles of pathogen transmission to humans have not been definitively identified for E. albertii infections. Moreover, the question of whether E. albertii-associated illness results from foodborne infection or foodborne toxicoinfection remains unresolved.
It is also important to determine the genetic characteristics of outbreak-associated pathogens to determine their propensity for disease and to monitor any developing changes in pathogenicity. Genome-wide phylogenetic analysis of E. albertii has identified five confirmed phylogroups (Ooka et al., 2015). Moreover, genes coding for a second, active type 3 secretion system (T3SS), designated ETT2 (for Escherichia coli T3SS 2) (Ooka et al., 2015), a cytolethal distending toxin (Pickett et al., 1996), a Shiga toxin (Stx2f) (Ooka et al., 2012), and invasion protein IbeA (Fiedoruk et al., 2014) have been recorded in E. albertii strains. However, little information is available about genetic aspects of outbreak-associated stains of E. albertii (Ooka et al., 2015; Gangiredla et al., 2018). Therefore, this study aimed to both determine the unique aspects of E. albertii outbreaks in Japan and examine the genetic characteristics of the outbreak-associated strains.
Materials and Methods
Outbreaks and sample collection
We examined all E. albertii outbreaks reported in Japan before 2015, which consisted of five outbreaks and one putative outbreak (Outbreaks 1–6, Table 1). Outbreak 1 was the only outbreak for which E. albertii was identified as the causative bacterium in real time (Table 1). In the other four outbreaks and one putative outbreak, E. albertii was misidentified as E. coli during the original investigations. Local governmental public health institutes later identified the causative pathogens as E. albertii. All six outbreaks were initially reported by medical doctors to health centers run by local governments as suspected food poisoning episodes in accordance with the Japanese Food Sanitation Act. These episodes were then investigated by food hygiene inspectors associated with the health centers in accordance with the Manual for Food Poisoning Survey produced by the Japanese Ministry of Health, Labour, and Welfare (Ministry of Health Labour and Welfare Japan, 2013). Patient specimens were collected by the food hygiene inspectors and then screened for causative foodborne pathogens at local governmental public health institutes. In this study, these episodes were re-examined based on personal communications between researchers in prefectural and municipal public health institutes, and through examination of any published study conducted at the time (Table 1). The study protocols were approved by the National Institute of Infectious Diseases for Public Health Ethics Committee (Approval No. 576).
Known Human Outbreaks (Including One Putative Outbreak) Caused by Escherichia albertii in Japan Between 2003 and 2015
Genome sequencing
Draft genome sequences of the E. albertii isolates from Outbreaks 1, 2, 4, 5, and 6 (Table 1) were generated using the Illumina HiSeq platform (Illumina, San Diego, CA). Sequence data for an isolate from Outbreak 3 (Table 1) were obtained from a previous study (Ooka et al., 2013). Sequence read assembly was performed as previously described (Ooka et al., 2015) (Supplementary Table S1), and the resulting genome sequences were deposited in the DDBJ/GenBank/EMBL databases (see Supplementary Table S2 for accession numbers). A neighbor-joining tree (Fig. 1) was constructed using the concatenated nucleotide sequences of 111 single-copy genes from the genomes of 35 E. albertii strains, 44 E. coli strains, 5 Escherichia fergusonii strains, and 15 strains belonging to cryptic Escherichia clades with a low probability of recombination.
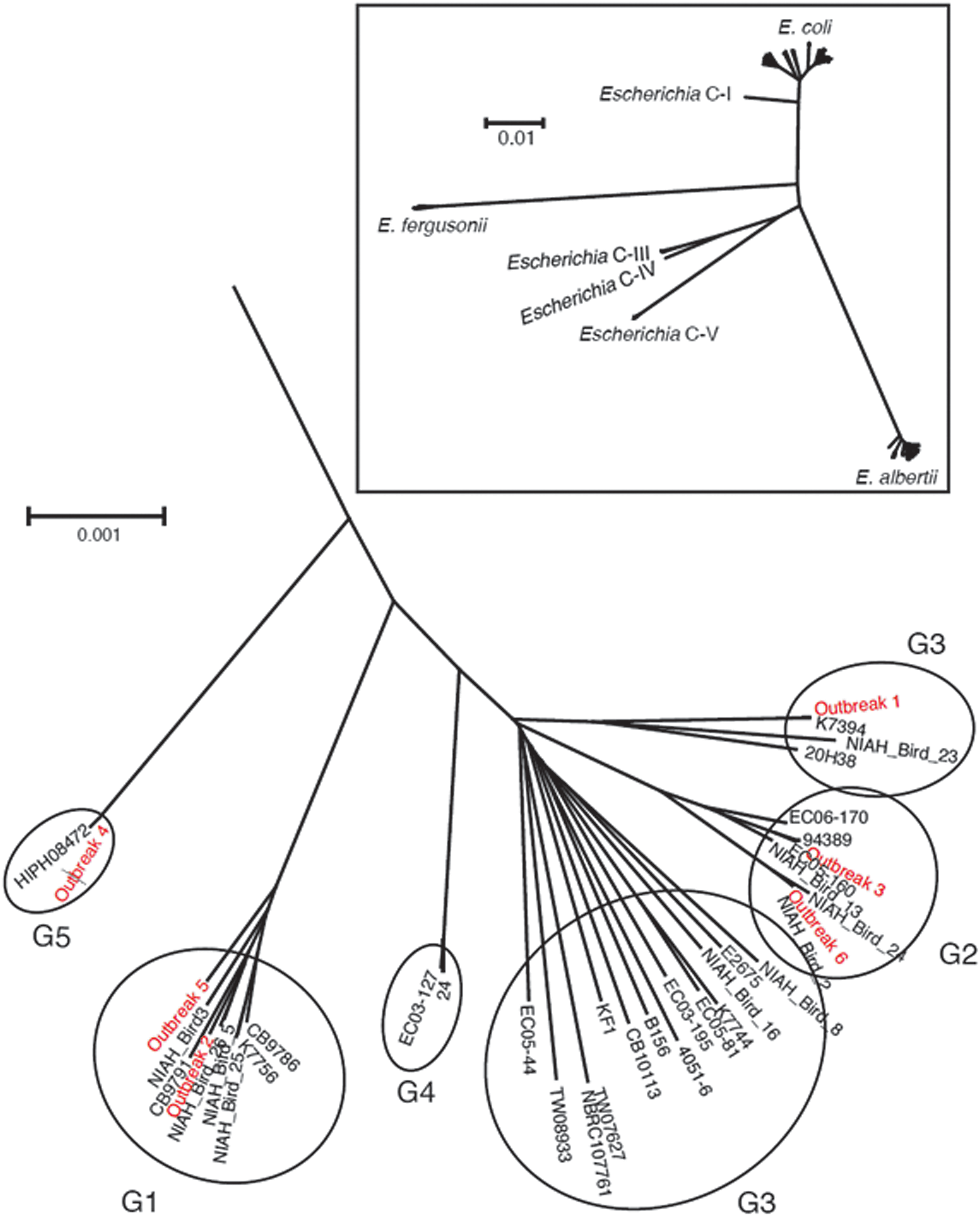
Genome-wide phylogenetic analysis of Escherichia albertii isolates and strains belonging to other Escherichia species and clades. The five E. albertii phylogroups are indicated by G1–G5. The tree was originally published in a previous study (Ooka et al., 2015), and the five outbreak strains were added in this study.
Pulsed-field gel electrophoresis analysis and virulence gene screening
Pulsed-field gel electrophoresis (PFGE) was carried out as previously described (Murakami et al., 2014b). The presence of virulence genes within each of the sequenced genomes was determined by comparison against the whole-genome sequence database GenEpid-J (Suzuki et al., 2016), and by using the ResFinder and VirulenceFinder tools available from the Center for Genomic Epidemiology (
Results
As shown in Supplementary Figure S1, the PFGE profiles (PFPs) of isolates from patients included in the same outbreak were identical, confirming that these episodes were outbreaks and not sporadic cases (data for Outbreaks 1, 2, 5, and 6, refer to Table 1). The PFPs of isolates associated with Outbreak 3 (available at
Outbreak 1 began on November 28, 2015, when students housed in a high school dormitory in Hiroshima Prefecture, Japan, began complaining of diarrhea and/or fever. Of the 83 students in the dormitory, 43 were affected. One additional patient not housed at the dormitory also exhibited symptoms (Table 1). All information pertaining to Outbreak 2 is listed in Table 1. In Outbreak 3, 48 out of 94 patrons (in two groups), who ate at a restaurant on May 29, 2011, exhibited symptoms ∼19 h after dining (Table 1). One group was a party of high school students. E. albertii was isolated from 29 of the 44 fecal specimens obtained from the affected patients, and the average incubation period was 19 h (Supplementary Fig. S2). Although a transmission vehicle was not identified, eae, which was harbored by the E. albertii isolate associated with the episode, was detected in well water from the restaurant. Therefore, the well water was considered the likely vehicle of pathogen transmission. In the case of Outbreak 4 (putative outbreak), two patients developed diarrhea after having dinner together. This was the first time the two patients had eaten together for a week. The symptoms of patient 1 began on October 20, 2008, and included diarrhea, abdominal pain, headache, and fever (37.7°C). The patient had seven episodes of diarrhea on day 1, four on day 2, and one on day 3. The patient ate raw vegetables, raw chicken meat, raw chicken egg, grilled chicken skewers, and fried chicken meat ∼13 h before the onset of symptoms. Patient 2 also suffered from diarrhea (about 30 times) and a low-grade fever (37°C) on day 1 of onset. Patient 2 ate the same meal as patient 1, except for the fried chicken, about 12 h before the onset of symptoms (Table 1). However, no fecal specimen from patient 2 was examined, meaning that the episode was designated a putative outbreak (Etoh et al., 2009; Murakami et al., 2014a). Outbreak 5 affected students and teachers from a high school in Fukuoka Prefecture (n = 409), who stayed at a campground in Oita Prefecture from July 18 to 20, 2005. The patients ate boxed lunches either from home or from caterers, as well as meals that they cooked themselves at the campground. Most campers (348/409) drank tap water from faucets at the campground, which was unchlorinated and derived from a sump pit. As shown in Supplementary Table S3, between day 1 of the camp and 3 days after the camp ended, a total of 273 of the 409 campers developed symptoms of E. albertii infection (Table 1). As shown in Supplementary Figure S1, the PFP of one patient isolate was identical to those of two isolates from the sump water, suggesting that contaminated sump water was responsible for at least some of the infections. Finally, boxed lunches were suspected to be the cause of Outbreak 6. A total of 31 people consumed the boxed lunches, 20 of whom developed diarrhea, abdominal pain, and/or fever (Table 1).
Outbreak strains showing no geographical or chronological relationships were classified into E. albertii clades G1, G2, G3, and G5 (Fig. 1). Although ETT2 is a cryptic second T3SS in the E. coli/Shigella lineage (Hayashi et al., 2001; Ren et al., 2004; Ideses et al., 2005), we found that three of the six E. albertii isolates (from Outbreaks 3, 5, and 6) possessed intact ETT2 regions (Fig. 2). The remaining isolates contained disrupted ETT2-encoding genes. Virulence gene screening revealed that more than half (44/70) of the tested virulence genes were present in all 5 outbreak strains, and that each strain contained more than 1 gene from 14 out of the 21 groups of virulence genes examined in this study. Only seven virulence genes were, respectively, harbored by a single strain (Supplementary Table S4).
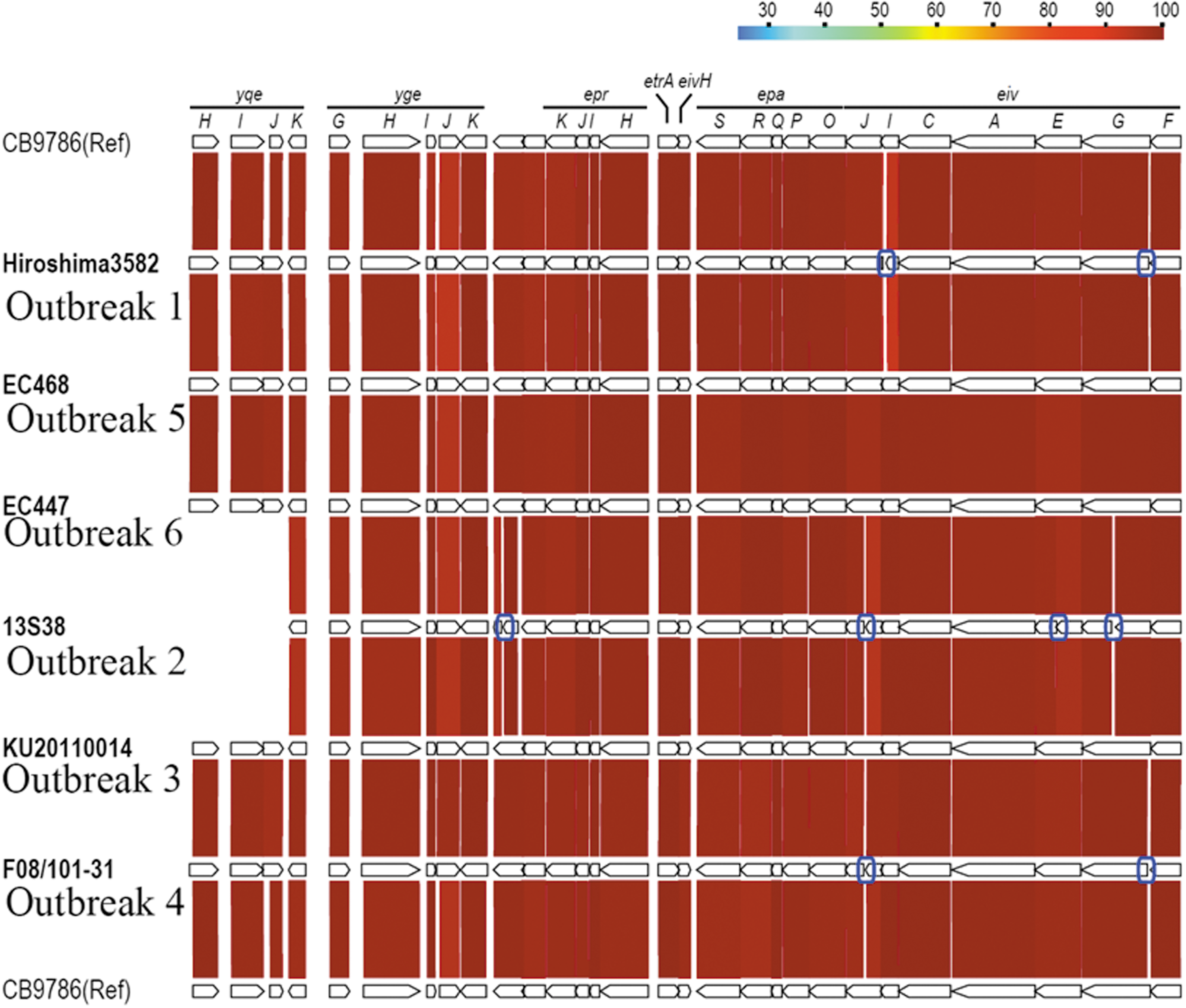
Structure of the Escherichia coli type three secretion system 2 (ETT2) region in Escherichia albertii. The gene organization within the ETT2 regions in the draft genome sequences of the six E. albertii isolates examined in this study is shown. For comparison, the ETT2 region of E. albertii strain CB9786 is also shown. Amino acid sequence identities between orthologous genes are presented, and the disrupted genes are indicated by blue boxes.
Discussion
In this study, the unique aspects of six E. albertii outbreaks in Japan were examined. The most common symptom among these six episodes was watery diarrhea (>80%), followed by abdominal pain (50–84%) and fever (37.0–39.5°C) (26–44%) (Table 1). In the largest outbreak examined in this study (Outbreak 5), nausea was a non-negligible symptom (104 of 273 cases, 38%), with a similar occurrence in Outbreak 2 (19 of 70 cases, 27%). The incubation periods reported by patients in Outbreak 3 were relatively varied (2–39 h), with a peak between 16 and 24 h (average of 19 h) (Supplementary Fig. S2). In comparison, the incubation periods of Outbreaks 4 and 6 were 12–13 h (n = 2) and 16–18 h (n = 20), respectively (Table 1). Therefore, the estimated average incubation period for E. albertii infection is in the range of 12–24 h. With regard to the vehicles of E. albertii transmission, restaurant foods, restaurant water, and boxed lunches were suspected in Outbreaks 2, 3, 4, and 6. In Outbreak 5, sump water at the campground was identified as one of the transmission vehicles (Table 1). The lack of isolates from the suspected contaminated food sources meant that we were unable to compare PFPs of the food isolates with those of the patient isolates as we were able to do with the waterborne outbreaks. Nevertheless, we predict that most of the outbreaks were foodborne or waterborne (Table 1). However, the unique aspects of this study are limited because data were restricted to that obtained from only a small number of publications describing the outbreaks, most of which focused on identification and examination of the causative pathogen. In addition, few official reports that were freely available were made at the time of the outbreaks, limiting the available epidemiological information.
There is also a lack of information on the differences in pathogenicity among E. albertii phylogroups and strains, and thus, this topic requires further study. Interestingly, we observed that different outbreak strains belonged to different clades (Fig. 1) and showed varied PFPs (Supplementary Fig. S1). The Outbreak 1 strain (phylogroup G3) was distinguished from the other strains by the presence of virulence gene ospG (protein kinase), while the Outbreak 4 strain (phylogroup G5) could be distinguished from the other strains by the presence of virulence genes astA (heat-stable enterotoxin), cma (lipid II-degrading bacteriocin colicin M), and stx2f (Shiga toxin).
Analysis of disease symptoms revealed that the presence of fever might be an occasional symptom of E. albertii infection. The E. albertii ETT2 appears to be distantly related to the Salmonella T3SS, which is encoded on Salmonella pathogenicity island 1 (Ren et al., 2004). The Salmonella T3SS plays a role in cellular invasion (Ren et al., 2004), resulting in fever following endotoxin-induced cytokine stimulation (Bhunia, 2008). While there are reports of a potentially functional ETT2 in E. albertii (Ooka et al., 2015), we found no remarkable differences in clinical symptoms between patients infected with strains containing intact or disrupted ETT2 regions. Further study is therefore needed regarding the underlying cause of fever in patients suffering from E. albertii infection.
Conclusions
Multiple E. albertii genotypes present in Japan are capable of causing outbreaks of diarrhea, abdominal pain, and occasionally fever in humans following the consumption of contaminated food or water. Moreover, the E. albertii outbreak strains examined in this study contain multiple virulence genes, some of which (e.g., LEE-encoded T3SS component genes) have the potential to cause severe disease in human patients.
Footnotes
Acknowledgments
We are grateful to the Director of the Hiroshima Prefectural Technology Research Institute, Public Health and Environment Center, the Director General of the Hiroshima Prefectural Western Center for Public Health, the Hiroshima Prefectural Government, and Dr. K. Ogata, Oita Pharmaceutical Association. We also thank Ms. Doi and Ms. Yamada of the National Institute of Infectious Diseases for their technical support, and Tamsin Sheen, PhD, from Edanz Group (
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by grants from the Japanese Society for the Promotion of Science (KAKENHI) (Grant No. JP19K10590), the Institute for Fermentation, Osaka (2016), the Toyo Suisan Foundation (2019), and the Research Program on Emerging and Re-emerging Infectious Diseases from the Japan Agency for Medical Research and Development (AMED) (Grant Nos. JP19fk0108033, JP19fk0108065, and JP19fk0108103).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.