
Abstract
Toxoplasma gondii is a worldwide prevalent, zoonotic parasite of major importance for public health, which can infect any warm-blooded animal species, including humans. Humans can get infected by consumption of meat from a chronically infected animal, by ingestion of sporulated oocysts (resulting from the sexual replication in felids), via contaminated water, soil, or vegetables, and by vertical transmission via the placenta. Infection through meat consumption is estimated to be one of the main sources of human toxoplasmosis cases in developed countries, and more specifically pork is considered to be responsible for 41% of foodborne human toxoplasmosis cases in the United States. To better assess the role of pork as a source of T. gondii infection in humans in Belgium, parasites were isolated from pigs to compare with human clinical isolates in a molecular epidemiological study. A positive result was obtained by magnetic capture-quantitative polymerase chain reaction for T. gondii in 14 out of the 92 hearts sampled during 2016 and 2017 from pigs raised in organic farms. From 9 of these 14 samples, parasites were isolated by mouse bioassay, demonstrating the presence of viable T. gondii in animals intended for human consumption. When genotyped and compared with 15 human isolates obtained during 2015 and 2016, a highly related structured population was demonstrated. Overall, these findings demonstrate the presence of infectious T. gondii in pigs intended for human consumption. Therefore, a potential transmission of T. gondii strains from pigs to humans could occur. However, both species could also be infected via a common source of infection such as oocysts. Furthermore, Belgium does not have an official surveillance program for T. gondii in human cases or food-producing animals; as a consequence, the detection of the infection source of a patient is very rare. Overall, this study reinforces the identification of pork as a potential risk for the consumers.
Introduction
T
The main infection routes described in the literature are consumption of meat from a chronically infected animal, ingestion of sporulated oocysts (resulting from the sexual replication in felids) via contaminated water, soil, or vegetables, and vertical transmission through the placenta (Cook et al., 2000; Tenter et al., 2000; Hide, 2016).
The consumption of raw or undercooked T. gondii-infected meat is considered a main route of transmission for humans (Cook et al., 2000): in developed countries, 50% of the infections are estimated to be meat borne (Scallan et al., 2011), and pork is considered to account for 41% of foodborne human toxoplasmosis cases in the USA (Batz et al., 2012). Besides the wide spectrum of intermediate hosts and the various routes of transmission, T. gondii shows a very structured genetic population in Europe and North America with three highly prevalent lineages also referred to as archetypal strains (types I, II, and III). The T. gondii strains circulating in Europe are primarily archetypal, and the type II strains are the most predominant in humans and livestock species (Dubey, 2010). However, no data are available on the genetic profile of the strains circulating in the Belgian human population or in any of the main livestock species.
The aim of this study was to genetically compare T. gondii isolates originating from humans and pigs to better estimate the role of pork as a source of infection for humans and the possible flow of strains from pigs to humans.
Materials and Methods
Porcine T. gondii isolates
To collect the isolates circulating in pigs, a study was designed with visits to a Belgian slaughterhouse where pigs originating from organic farms are slaughtered. Based on the preliminary results of a T. gondii seroprevalence study in Belgium, in which the prevalence in organic farms was consistently higher than in intensive production systems (Jennes et al., 2017), all the samples were collected from a slaughterhouse specialized in organic pigs located in Malmedy (Wallonia, Belgium). The farms were selected randomly based on the slaughtering schedule established by the slaughterhouse. A total of 92 pigs representing 17 Belgian organic farms were sampled during 2016. First, the pigs were serologically screened for Toxoplasma antibodies, and then confirmation of the presence of the parasite in heart was done by magnetic capture-quantitative polymerase chain reaction (MC-qPCR), where after the parasite was isolated by mouse bioassay.
Serological screening
The cardiac fluid from the 92 pigs was evaluated using the modified agglutination test for the detection of antibodies against T. gondii as described by the manufacturer's instructions (Toxo-Screen DA kit; Biomerieux). In brief, the cardiac fluid samples were diluted in phosphate-buffered saline (PBS) at a 1/40 dilution, and 25 μL of each dilution was applied in a round bottom microtiter plate with 25 μL 2-mercaptoethanol at 0.2 mol/L. Subsequently, 50 μL of a 1/5 dilution of whole formalinized tachyzoites in bovine albumin borate saline (supplied in the kit) buffer was added, mixed for 1 min on a plate mixer at 300 rpm, and incubated overnight at room temperature. A negative and a positive control were both supplied by the manufacturer and consisted of goat serum. As an external control, positive and negative pig serum samples obtained from earlier experimental infections were tested on the same plate. The cutoff established to consider the animals as seropositive was 1:80; this high cutoff was established to avoid unnecessary animal testing and ensure the presence of the parasite in quantities that allowed the isolation of the strains for further molecular characterization.
Confirmation of the presence of the parasite by MC-qPCR
To confirm the actual presence of the parasite and determine the parasitic load, the magnetic capture real-time PCR was performed as described by Gisbert Algaba et al. (2017). Heart was selected as the target sample to detect and isolate the parasite as it was identified as one of the predilection tissues for T. gondii in experimentally infected pigs (Gisbert Algaba et al., 2018).
In brief, ±100 g of the heart samples were homogenized in the presence of lysis buffer (100 mM Tris HCL, 5 mM EDTA, 0.2% SDS [sodium dodecyl sulfate], 200 mM NaCl, and 40 mg/L proteinase k [30 mAnson-U/mg; Amresco, OH], pH = 8.0 ± 0.2) with a pedal homogenator (Labconsult, Brussels, Belgium) and lysed overnight at 55°C. Fat and cell debris were then removed by centrifugation, and any free biotin possibly present in the crude extract was removed by adding streptavidin-coated agarose beads (binding capacity >330 nmol/mL; Solulink, San Diego).
The specific biotin-labeled probes against T. gondii (Accession No. AF146527) and cellular r18S DNA were added to the biotin-free lysates and the samples were first denatured at 95°C followed by an incubation at room temperature to hybridize the probes with the complementary target sequences. After hybridization, the biotin-labeled probes bound to the target DNA were captured using streptavidin magnetic beads (binding capacity >2.5 nmol/mg; Solulink) and after washing and resuspension in elution buffer, the target DNA was released from the beads by exposure to ultraviolet light.
Finally, the qPCR was performed on the final DNA extract to detect the presence of T. gondii DNA and the results were analyzed using the Bio-Rad CFX manager software to obtain the crossing point (Cp) values.
All the samples with an exponential amplification curve crossing the threshold (Cp) were considered positive for T. gondii, samples with no amplification curve for the T. gondii target, but amplification of the NCIAC, were considered negative. As described in Gisbert Algaba et al. (2017), the limit of detection of the method is 65.4 parasites per 100 g of tissue. For each round of samples, a positive control with a known number of parasites (calibrator) was performed to correct for possible deviations due to manipulation errors.
Isolation of the strains, mouse bioassay
Only after confirmation of the presence of T. gondii DNA by MC-qPCR, 100 g of cut heart tissue were homogenized in a blender with 150 mL sterile saline (0.9% NaCl) supplemented with 0.25% trypsin (Gibco) and gentamycin (0.4 mg/mL). The homogenate was then transferred to a 2 L flask and incubated in a water bath at 37°C for 1 h 45 min under continuous stirring (magnetic stirrer). The digest was then filtered through gauze, transferred to a 500 mL centrifuge bottle, and centrifuged at 2100 g at room temperature for 20 min.
The supernatant was then removed and the pellet resuspended in 100 mL of saline with gentamycin and centrifuged at 2100 g at room temperature for 15 min. This step was repeated once more to remove most of the trypsin present in the pellet. Next, the final pellet was resuspended in 10 mL of PBS supplemented with gentamycin and intraperitoneally injected into five Swiss white female mice, 1 mL each.
The mice were observed daily and a humane endpoint was defined to limit the suffering, in case of disease (the mice were euthanized, and the lungs and, if present, ascites from the peritoneal cavity were collected and tested in qPCR) to confirm the presence of T. gondii. Six weeks after the inoculation, the surviving mice were euthanized, their blood collected and tested by indirect immunofluorescence assay as described by Verhelst et al. (2015) (Ethics Committee Licence No: 20140704-02). The total number of animals used for this study were 105.
Human Toxoplasma isolates
The genotypes of 15 T. gondii isolates originating from cases of congenital toxoplasmosis in Belgium collected within the frame of the National Reference Center (NRC) for Congenital Infections during 2015 and 2016 were determined and analyzed at the Toxoplasma Biological Resource Center (CHU Limoges). Since no official surveillance program or compulsory notification for T. gondii in human cases exists in Belgium, all isolates were achieved on a voluntary basis and might not represent the population as a whole.
Genotyping
All the isolates were genotyped as described by Ajzenberg et al. (2010). In brief, T. gondii DNA was extracted from 200 μL of brain homogenate using the QIAamp DNA Mini Kit (Qiagen) according the to the manufacturer's instructions. Once extracted, a multiplex PCR was performed to amplify different microsatellite markers located in different genes (Supplementary Table S1). These markers are grouped in two different levels of discrimination: (i) typing markers (TUB2, W35, TgM-A, B18, B17, M33, IV.1, and XI.1) able to discriminate between the main T. gondii genotypes (I, II, and III) and (ii) fingerprinting markers (M48, M102, N60, N82, AA, N61, and N83) with a high power of discrimination able to differentiate between different strains.
A neighbor-joining tree was reconstructed from microsatellite data to examine the relationship between isolates obtained from humans and pigs originating from Belgium. The tree was constructed as described by Ajzenberg et al. (2016).
Results
The study yielded a total of 15 seropositive pigs (16.3%) representing 6 organic farms. The actual presence of the parasite in hearts was confirmed by MC-qPCR in 14 pigs (15.21%) representing 5 organic farms (Farms: A, D, G, L and O). In farms D and G, T. gondii was found in all the animals tested (D:1/1 and G:6/6), while in farms A, L, and O, only some of the tested animals were found positive (A: 1/3; L: 4/6; O: 2/3). Finally, nine isolates were successfully obtained by mouse bioassay demonstrating the infectivity and viability of the parasites present in these samples. The results are summarized in Table 1.
Overview of the Parasitic Load from Naturally Infected Animals
All mice were euthanized respecting the human endpoint and the strain could not be isolated for genotyping studies.
Cp, crossing point; MC-qPCR, magnetic capture-quantitative polymerase chain reaction; NA, negative result; Neg, negative result; np, not performed; P. load, log 10 (number of parasites per 10 g of tissue); Pos, positive result.
All the human (N = 15) and pig isolates (N = 9) were identified as archetypal type II strains (Supplementary Table S2). In addition, the fingerprinting analysis resulted in the identification of three different clusters within the pig isolates. Two different clusters were found inside one farm, and the other cluster identified represented all the genotyped samples from other farms (n = 2). The fourth cluster was found in two different human patients. All the data are represented in a neighbor-joining tree in Figure 1.
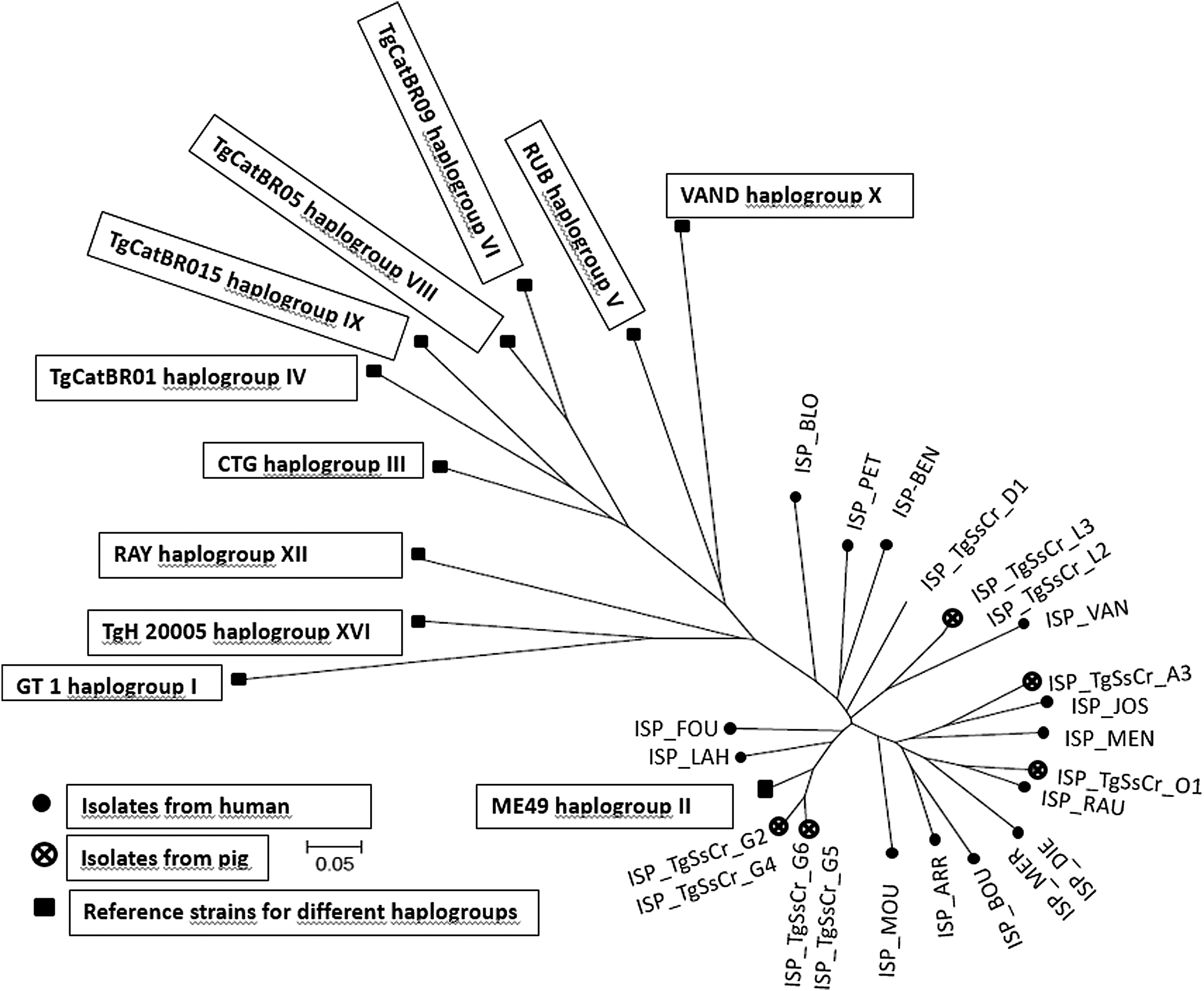
Neighbor-joining clustering of Toxoplasma gondii strains based on 15 microsatellite markers. Squares represent the reference strains for different haplogroups: GT1 (haplogroup I), ME49 (haplogroup II), CTG (haplogroup III), TgCatBr01 (haplogroup IV), TgCatBr05 (haplogroup VIII), TgCatBr15 (haplogroup IX), RUB (haplogroup v), VAND (haplogroup X), RAY (haplogroup XII), TgH20005 (haplogroup XVI), and TgCatBr09 (haplogroup VI); points represent the isolates from human; the crosses encircled represent the isolates from pig.
Discussion
In the present study, we were able to successfully isolate and genotype nine strains originating from pigs raised in five organic farms located in Belgium. All the isolates from pigs were identified as classical type II strains. Two different clusters were found on the same farm indicating that the animals had been exposed to different strains and/or infectious sources. Conversely, both of the isolates originating from another farm belonged to a single cluster, indicating a common source of infection (n = 2).
A previous study performed in Belgian foxes showed also a highly clonal population of T. gondii strains: 25 strains out of 26 were identified as type II and only 1 as a type III (De Craeye et al., 2011). In addition, studies performed in several European countries have consistently identified type II as most prevalent in livestock; such as 17 strains isolated from pigs in the Czech Republic (Slany et al., 2016) and 100% and 97% of the isolates from sheep in the Netherlands and France, respectively (Halos et al., 2010; Opsteegh et al., 2010). A recent study performed in industrial pigs intended for human consumption in Italy showed an overall presence of T. gondii DNA of 13.6%. From these isolates, 42.6% were classified as type II, while type I and III accounted for 28.6% of the isolates each (Vergara et al., 2018). Similarly, out of nine isolates successfully genotyped originating from pigs slaughter in Serbia resulted in seven type II and two type II strains (Kuruca et al., 2019). Altogether, these results confirm the highly structured T. gondii population found in central Europe.
In contrast, one study performed in north-western Italy showed that genotype I seemed to be the most prevalent in the analyzed samples from different species (Battisti et al., 2018). Moreover, similar studies performed in South America prove a much greater variability within the isolates genotyped (Alvarez et al., 2014; Miura et al., 2019).
All the human isolates analyzed in the current study were also identified as type II strains and two strains had identical genotyping results suggesting a common source of infection (Supplementary Table S2). These findings are in agreement with other studies carried out in France, Portugal, and Poland where different collections of T. gondii isolates were genotyped and the majority of the isolates were identified as type II (Dardé et al., 1992; Howe and Sibley, 1995; Ajzenberg et al., 2002; Djurković-Djaković et al., 2006). For example, in France, 84% the strains isolated between 1987 and 2001 were identified as type II; the other genotypes, type I and type III accounted for 8.14% and 2.33%, of the isolates, respectively (Ajzenberg et al., 2002). The results of these studies (high predominance of type II strains) contrast to the ones obtained, for example, in Denmark, where more than a third of the successfully genotyped strains were nontype II such as Africa 1, HG12-like, and atypical genotypes (Jokelainen et al., 2018), or in Portugal, where although most of the isolates were identified as type II (66.7%), 12.5% and 20.8% were identified as type I and mix/recombinant, respectively (Vilares et al. 2017). Strikingly, a study performed in Poland with isolates originating from humans identified 100% of their isolates as type III strains (Nowakowske et al., 2006). Type III strains have been previously associated with wildlife in European countries; therefore, the high prevalence of this genotype in Poland can indicate a potential different T. gondii population in Poland.
Conclusions
Overall, in the present study, 14 of the hearts originating from pigs raised in organic farms tested positive by MC-qPCR, demonstrating the presence of the parasite's DNA in animals intended for human consumption. These results demonstrate the higher risk that organic farms may pose to the consumer regarding T. gondii infections. Viable parasites were isolated in 9 out the 14 hearts demonstrating the presence of infectious T. gondii in pigs intended for human consumption. Also, the highly related population structure of all the strains isolated in humans and pigs during 2015, 2016, and 2017 indicate a common source of infection such as oocysts and/or transmission of T. gondii strains from pigs to humans. However, the latter is not yet clearly demonstrated. All these findings reinforce the identification of pork as a potential T. gondii source of infection for the consumers.
Footnotes
Acknowledgments
The authors thank the colleagues of the WIV-ISP institute for their technical assistance.
Funding Information
This study was granted by the Belgian Federal Public Service for Health, Food Chain Safety and Environment (grant RF 13/6274).
Supplementary Material
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.