
Abstract
Salmonella enterica remains an important foodborne pathogen in all regions of the world, with Typhimurium as one of the most frequent serotypes causing foodborne disease. However, the past two decades have seen a rapid worldwide emergence of a new Salmonella serotype, namely monophasic variant of S. Typhimurium, whose antigenic formula is 1,4,[5],12:i:-. It has become one of the 2–5 most common Salmonella serotypes responsible for animal and human infections in different regions. The global epidemic of monophasic S. 1,4,[5],12:i:- has mainly been characterized by an increase in multidrug-resistant S. 1,4,[5],12:i:- isolated in Europe since 1997. The unexpected link to swine has escalated monophasic S. Typhimurium infections to the status of a global public health emergency. The large-scale application of whole genome sequencing (WGS) in the last 10 years has revealed the phylogenetic associations of the bacterium and its antimicrobial resistance (AMR) genes. Local and global transmission reconstructed by WGS have shown that different clones have emerged following multiple independent events worldwide, and have elucidated the role of this zoonotic pathogen in the spread of AMR. This article discusses our current knowledge of the global ecology, epidemiology, transmission, bacterial adaptation, and evolution of this emerging Salmonella serotype.
Introduction
Non-typhoidal Salmonella,
Since the first S. 1,4,[5],12:i:- identified in the late 1980s from poultry in Portugal (Machado and Bernardo, 1990), it has become one of the most common Salmonella serotype and was strongly associated with the swine food chain, especially in Europe and the United States of America, suggesting a link of human infections with this serotype to pork and pork products (de la Torre et al., 2003; Hauser et al., 2010; Elnekave et al., 2018). In addition, most emerging infectious diseases are driven by socioeconomic, environmental, and ecological factors, such as the use of antimicrobials, resulting in the emergence of multidrug-resistant (MDR) bacteria. With high transmission capacity, MDR, S. 1,4,[5],12:i:- is increasingly becoming a global hazard and challenge.
Epidemiological success depends on selective advantage of epidemic clones, resulting from their unique genotypes. Bacterial adaptation to survive in changing conditions may involve mechanisms such as virulence factors, fitness factors, and AMR. The objective of this review is to investigate the epidemiological and genetic characteristic and mechanisms of antibiotic resistance in S. 1,4,[5],12:i:-, and to reveal the evolution path and bacterial adaptation of this dominant serotype.
Phenotypic and Genotypic Characteristics of S. 1,4,[5],12:i:-
Serological and genetic characterization of S. 1,4,[5],12:i:-
Serotyping of Salmonella is based on antigenic variability of lipopolysaccharides, flagellar proteins, and capsular polysaccharides (Grimont and Weill, 2007). Most Salmonella strains are motile, with H antigen expressed, by means of peritrichous flagella. The expression of H antigen is regulated by expression of fliC, fljBA, and hin genes (Switt et al., 2009). The majority of the Salmonella serotypes are biphasic, which could produce both phase 1 flagellin (FliC) and phase 2 flagellin (FljB) under phase switching by the H segment (Yamamoto and Kutsukake, 2006). For example, some nonbiphasic Salmonella have been described as S. Typhimurium-like strains, such as 1,4,[5],12:-:1,2, or 1,4,[5],12:i:-, or 1,4,[5],12:-:- (European Food Safety Authority, 2010).
Widely spread S. 1,4,[5],12:i:- strains are monophasic and lack expression of phase 2 flagella. The latter characteristic may be due to different mutations (including point mutations) and partial or complete deletions in fljB and adjacent genes (Zamperini et al., 2007; Soyer et al., 2009). An inverted IS26 sequence in the fljBA operon plays an important role in the form of various deletion types in fljBA (Laorden et al., 2010; Lucarelli et al., 2012; Garcia et al., 2013, 2016; Boland et al., 2015). As more isolates were investigated from different regions, some clades lacked part of the fljBA locus, resulting from deletions ranging in size different from the types above. However, to a less frequent extent, other mechanisms can lead to the monophasic status such as mutations and deletions of fljA and hin. Furthermore, the invertible promoter that controls the expression of fljB and fliC may become blocked in one position, thereby allowing only the expression of the fliC gene (Switt et al., 2009).
However, monophasic strains, in which the phase-2 flagellar antigen is not detected serologically, but can be detected by PCR, have also been observed, and are defined as “inconsistent/atypical” S. Typhimurium variants (Hopkins et al., 2010; Bugarel et al., 2012; Boland et al., 2015). Isolates harboring an fljB coding sequence have been reported in several countries (Zamperini et al., 2007; Hauser et al., 2010; Barco et al., 2011, 2012, 2014a; Bugarel et al., 2012; Hopkins et al., 2012; Wasyl and Hoszowski, 2012; Mandilara et al., 2013; Prendergast et al., 2013; Arguello et al., 2014; Ido et al., 2014). This phenomenon may be caused by the following: (i) deletions in part of fljB (Soyer et al., 2009); (ii) absence of the hin gene (Barco et al., 2014a); (iii) nonconservative point mutations identified in fljA and hin genes (Barco et al., 2014a); and (iv) serotype inconsistent strains in replicates (Soyer et al., 2009).
In summary, the evolution of S. 1,4,[5],12,i:- may represent a single emergence event, followed by subsequent rearrangements and/or additional deletions in the region surrounding the fljBA operon, resulting in different S. 1,4,[5],12,i:- lineages.
Relationship of S. 1,4,[5],12:i:- to S. Typhimurium
It is generally believed that S. 1,4,[5],12:i:- are monophasic variants of S. Typhimurium 1,4,[5],12:i:1,2, whose flagella are encoded by fljB. Previous studies have confirmed this presumption in several aspects, including antibiotic-resistant pattern (de la Torre et al., 2003; Lucarelli et al., 2010), S. Typhimurium-specific genes (Echeita et al., 2001; Amavisit et al., 2005; Hopkins et al., 2010; Trupschuch et al., 2010; Ido et al., 2011), pulsed-field gel electrophoresis (PFGE)/multiple locus variable-number tandem repeat analysis (MLVA)/multilocus sequence typing (MLST) phylogeny (de la Torre et al., 2003; Zamperini et al., 2007; Dionisi et al., 2009; Soyer et al., 2009; Tavechio et al., 2009; Hopkins et al., 2010; Barco et al., 2015), microarray analysis (Garaizar et al., 2002; Hauser et al., 2010), and whole-genome sequencing (WGS) analysis (Petrovska et al., 2016; Ngoi et al., 2018).
Despite the similarity between this new serotype and S. Typhimurium, they show distinct epidemiological, genomic, and phenotypic features. For example, S. 1,4,[5],12:i:- was less heterogeneous compared with S. Typhimurium in respect of PFGE and MLVA profiles, and slight, but consistent, difference existed on biochemical test and antimicrobial susceptibility profiles between this new serotype and S. Typhimurium (Agasan et al., 2002; Soyer et al., 2009; Barco et al., 2015).
In summary, S. 1,4,[5],12:i:- was proved to be a monophasic variant of S. Typhimurium, according to similarity of its genetic characteristics, which causes a lack of flagellar antigen in the second phase. The detailed relationship between S. 1,4,[5],12:i:- and S. Typhimurium from different sources and geographical locations is likely to become clearer with the development of WGS and public database for S. 1,4,[5],12:i:-.
Identification methods for S. 1,4,[5],12:i:-
Accurate identification of S. 1,4,[5],12:i:- is important to avoid unnecessary regulatory actions by misclassification. In fact, serotyping alone could lead to misidentifications (Seixas et al., 2016). Different PCR protocols focused on genes encoding for flagellar phase 2 have been developed (Echeita and Usera, 1998; Barco et al., 2012; Bugarel et al., 2012; Prendergast et al., 2013; Maurischat et al., 2015). Tennant et al. (2010) described a simple, yet effective, PCR method that targeted two genes: fljB encoding flagellar phase 2, and the STM-specific position of an IS200 element within the fliA–fliB intergenic region. This protocol is recommended by the European Food Safety Authority as a PCR protocol to confirm the lack of the second phase antigen (European Food Safety Authority, 2010). However, for the “inconsistent” monophasic strains, the PCR methods are insufficient to distinguish the monophasic strains from S. Typhimurium. Therefore, a comprehensive and accurate identification method for S. 1,4,[5],12:i:- is needed.
WGS data can be used to describe the phylogeny of S. Typhimurium and reveal the detailed mutations around the fljB locus. With the convenient application of WGS technology and the decline in price, WGS is rapidly replacing current serology methods for subtyping and molecular typing methods for surveillance purposes (Petrovska et al., 2016; Mastrorilli et al., 2018). Therefore, WGS will probably become a new gold standard for the identification of S. 1,4,[5],12:i:-.
Epidemiologic Characteristics of S. 1,4,[5],12:i:-
Distribution of S. 1,4,[5],12:i:- in humans
In the past 20 years, S. 1,4,[5],12:i:- has become a dominant serotype for animal and human infections through rapid evolution and spread and one of the most common Salmonella serotypes responsible for human infections in different countries (European Food Safety Authority, 2010; Hopkins et al., 2010; Elnekave et al., 2018). In European Union member states, monophasic S. 1,4,[5],12:i:- accounted for ∼8% of total human salmonellosis cases reported yearly from 2014 to 2016 and ranked third among the most reported Salmonella serotypes (Supplementary Fig. S1) (European Food Safety Authority, 2017). In Germany, S. 1,4,[5],12:i:- accounted for nearly half (42.2%) of all S. Typhimurium isolates from humans (Trupschuch et al., 2010). In human cases as well as in food and animal products, isolation rates of S. 1,4,[5],12:i:- have increased 10-fold between 2005 and 2010 in France (Bugarel et al., 2012). In the United States, S. 1,4,[5],12:i:- is the fifth most common Salmonella serotype in human salmonellosis according to the Laboratory-based Enteric Disease Surveillance system. Remarkably, the incidence rate of infection with Salmonella overall has increased by 33% since a trough in 2001, while incidence rates of S. 1,4,[5],12:i:- infection have increased more than five times over the same period (CDC, 2016). In our unpublished work based on a national surveillance system, S. 1,4,[5],12:i:- has overtaken S. Typhimurium and ranked the second most frequently encountered serotype in patients in Henan, China, just following Salmonella Enteritidis.
Global distribution of S. 1,4,[5],12:i:- before 2009 has been summarized by Switt et al. (2009). In this study, we review the peer-reviewed literature on monitoring of S. 1,4,[5],12:i:- from a variety of countries from 2008 to 2018 (Supplementary Table S1 and Fig. 1). S. 1,4,[5],12:i:- has been reported in five continents (Asia, Europe, North America, South America, and Oceania), and in 79% of reports, S. 1,4,[5],12:i:- could be isolated from humans. Most reports on S. 1,4,[5],12:i:- are in Europe. The proportion of monophasic serotype isolates has increased from 2005 (Bugarel et al., 2012).
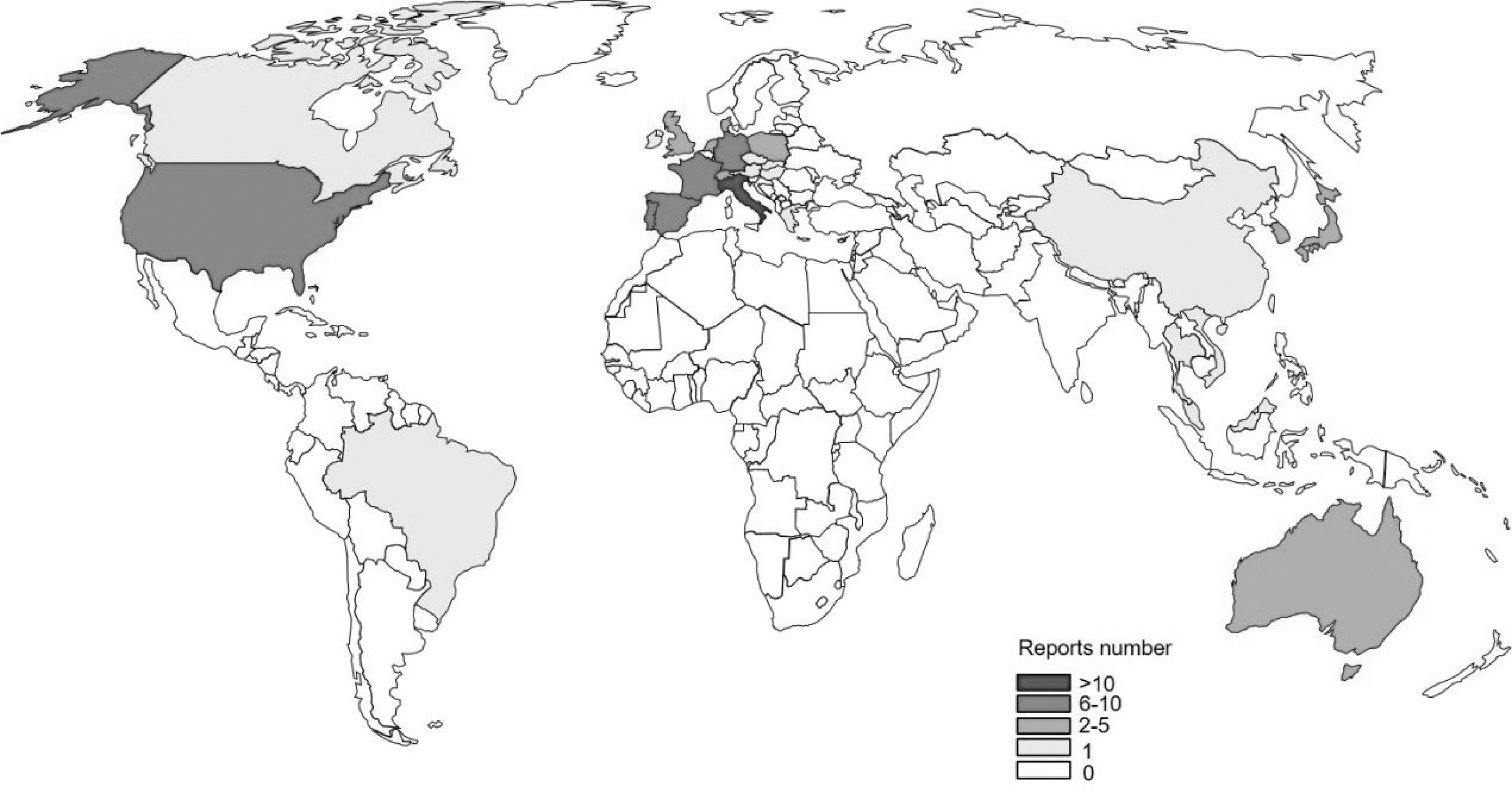
Peer-reviewed reports on monitoring of S. 1,4,[5],12:i:- around the world from 2008 to 2018.
Distribution of S. 1,4,[5],12:i:- in foods and animals
From an epidemiological point of view, S. 1,4,[5],12:i:- is strongly associated with the swine food chain, especially in Europe and the United States, suggesting a potential link between human infections with this serotype and pork and pork products in these areas (de la Torre et al., 2003; Mossong et al., 2007; Hauser et al., 2010; Lucarelli et al., 2010, 2012; Mourao et al., 2014). For example, most S. 1,4,[5],12:i:- strains (48%) in Germany were isolated from pork between 2006 and 2008, followed by cattle (13%), poultry (5%), and other sources (Hauser et al., 2010). In Poland, low occurrence of monophasic S. Typhimurium in comparison with other countries might be explained by low prevalence (0.6–4.8%) in pig production (Wasyl and Hoszowski, 2012). Even though strongly linked to pig production, this monophasic serotype was less frequently recovered from pig compared to S. Typhimurium in Belgium (Van Damme et al., 2018). In Japan and China, cattle were observed as an important animal reservoirs for S. 1,4,[5],12:i:- (Kurosawa et al., 2012; Ido et al., 2015; Yang et al., 2015), which was detected frequently less than S. Typhimurium. Besides pig and cattle as important sources of S. 1,4,[5],12:i:-, in the United States (Kawakami et al., 2016), S. 1,4,[5],12:i:- isolates from the outbreak were also isolated from frozen feeder mice (Cartwright et al., 2016). Even the community's private water system was identified as the source of an outbreak of S. 1,4,[5],12:i:- infection in a rural community in the United States (Kozlica et al., 2010).
In summary, although associated mostly with the swine food chain, S. 1,4,[5],12:i:- can be isolated from diverse sources, including humans, livestock like cattle and companion animals, food, and the environment. Therefore, with the wide spread in livestock, meat, and humans, S. 1,4,[5],12:i:- has become an important new foodborne pathogen.
Outbreaks caused by S. 1,4,[5],12:i:-
Worldwide distribution of S. 1,4,[5],12:i:- before 2009 has been reviewed by Switt et al. (2009), and in this study, we present some reports of more recent outbreaks of S. 1,4,[5],12:i:- (Table 1). This monophasic variant has also been reported as the cause of several outbreaks in Europe, including two outbreaks associated with salami and pork products in 2009 and 2010 in two regions of Northeast and Northwest Italy (Barco et al., 2014b; Andreoli et al., 2017); three outbreaks in France connected with imported beef and dried pork sausage in 2010 and 2011 (Bone et al., 2010; Gossner et al., 2012; Raguenaud et al., 2012); an outbreak associated with consumption of dried pork sausage in Spain in 2011 (Arnedo-Pena et al., 2016); an outbreak related to irrigation water channels in Central Italy in 2013–2014 (Cito et al., 2016); and an outbreak associated with roast pork in Spain in 2016 (de Frutos et al., 2018). In the United States, this monophasic variant strain has been involved in many outbreaks, including a waterborne outbreak in 2008 (Kozlica et al., 2010); an international outbreak associated with exposure to pet reptiles and frozen feeder rodents in 2008–2010 in the United States (Cartwright et al., 2016); a multistate outbreak associated with pork products in Washington State in 2015 (Kawakami et al., 2016); and a multistate outbreak linked to kosher chicken in 2018 (
Selected Peer-Reviewed Reports of Outbreaks of S. 1,4,[5],12:i:-
In summary, S. 1,4,[5],12:i:- associated foodborne outbreaks can occur in different regions and be linked to different foods, especially pork products. With the globalization of commodities and the increase of international trade in food products, foodborne outbreaks of S.1,4,[5],12:i:- are becoming an urgent challenge.
Prevalent clones of S. 1,4,[5],12:i:-
In the past two decades, there were three distinct clones (i.e., Spanish, United States, and European) at different times and in different regions (Table 2). At present, the European clone has gradually overtaken the Spanish and U.S. clones as the most prevalent (Elnekave et al., 2018).
Characteristics of Different Clonal Groups of S. 1,4,[5],12:i:-
AR, antibiotic resistance; MLST, multilocus sequence typing; MDR, multidrug resistant.
This initial clone of S. 1,4,[5],12:i:- associated with pigs and pork products was identified as the Spanish clone from 1997, ascribed to phage type U302. It has been shown to differ from S. Typhimurium LT2 by five major deletions, designated clusters I–V (Mourao et al., 2014). The Spanish clone displays plasmid-mediated resistance to up to seven antimicrobial drugs: ampicillin, chloramphenicol, gentamicin, streptomycin/spectinomycin, sulfonamides, tetracyclines, and trimethoprim (ACGSSuTTp type). The genes are carried by a family of IncA/C plasmids that harbor (pUOSTmRV1 like) or do not harbor (pUO-STmR1 like) genes of pSLT, the specific virulence plasmid of S. Typhimurium (de la Torre et al., 2003; Antunes et al., 2011; Garcia et al., 2011, 2013, 2014). Most S. 1,4,[5],12:i:- isolates from the Spanish clone were detected in the fljBA operon, all starting at the same position, at the STM2758 gene, and followed by an IS26 insertion. Results indicated that transposition of IS26, presumably donated by pUO-STmR/RV1-like plasmids originally acquired by biphasic S. Typhimurium, was involved in the deletion of the fljBA operon and surrounding genes, and hence, is responsible for the monophasic phenotype displayed by the isolates (Garcia et al., 2013). It appears to be characterized by deletions of fljA, fljB, hin, and iroB and conservation of STM2757 (Garaizar et al., 2002; Laorden et al., 2010; Garcia et al., 2013). It was mainly reported in isolates of pig origin in Spain (1998–2000) (de la Torre et al., 2003), piggeries in Portugal (2006–2007) (Antunes et al., 2011), and food and veterinary practice in France (2001–2010) (Bugarel et al., 2012).
The second clonal lineage “U.S. clone,” emerged around 2004, was characterized with particular genomic deletion pattern surrounding the fljBA operon, but maintaining hin and iroB genes and the STM1053-1997 region (all absent from Spanish clone) (Soyer et al., 2009). Besides, MDR was rare in U.S. clone strains (Clade B-I ST19 without the ASSuT genotype), which was different from isolates in Spanish clone (Hauser et al., 2010). These data provided support that U.S. clone during 1999–2016 may be distinct from Spanish S. 1,4,[5],12:i:- isolates (Hauser et al., 2010). Besides, ∼2% isolates from food and veterinary in France (2001–2010) (Bugarel et al., 2012) and a Southern European clone (carrying a sul3-type I integron within IncR plasmids, ST19) described in Portugal were similar to U.S. clone (Mourao et al., 2014). By the application of WGS, the relationship between United States and European strains was compared. Salmonella 1,4,[5],12:i:- circulating in swine in the United States Midwest in 2014–2016 is likely to be part of an emerging MDR clade first reported in Europe (Elnekave et al., 2018).
The major European clone from 2005 (ASSuT phenotype, RR1–RR2/RR3 resistance regions, ST34) has spread in several countries across the European Union. It was characterized by chromosomally encoded resistance to ASSuT, and lacked the typical S. Typhimurium virulence plasmid (pSLT) (Hauser et al., 2010; Hopkins et al., 2010; Lucarelli et al., 2010, 2012; Majtan et al., 2011; Gallati et al., 2013; Arguello et al., 2014; Barco et al., 2014b; Garcia et al., 2014). The deletion profile of this clone was characterized by absence of the fljA, fljB, and hin genes and presence of STM2757, as in Spanish strains, but it harbored a conserved iroB (Lucarelli et al., 2012). Possession of Salmonella genomic island 3 (SGI3), which encodes resistance to heavy metals, including copper and zinc, is also characteristic of this clone. This clone was ascribed mainly to phage types DT193 and DT120 and ST34 (Hauser et al., 2010; Hopkins et al., 2010; Lucarelli et al., 2010; Bugarel et al., 2012; Mandilara et al., 2013). And more recently, it also was epidemic in the United States (Elnekave et al., 2018) and Canada (Switt et al., 2009; Mulvey et al., 2013), which in all cases, presented similar phenotypic and genotypic ASSuT markers. Even in Spain, the prevalent phage types of 1,4,[5],12:i:- from pigs, wild birds, rodents, and farm environmental samples changed to U311 (40.7%), DT195 (22%), and DT193 (8.4%) between 2008 and 2012. These phage types were more similar to those of European strains than to the well-characterized Spanish clone (Andres-Barranco et al., 2016). The European clone S. 1,4,[5],12:i:- was first reported in cattle in 1998 and became the predominant clone in many other countries such as Japan (2013–2017) (Arai et al., 2018) and the United States.
Besides the generally reported clones mentioned above, some endemic clones have also been revealed in several countries. The diverse Salmonella Typhimurium monophasic variant fljB+ population investigated in Belgium and characterized by numerous PCRs and MLVA profiles does not represent a single lineage, but rather a diversity of lineages originating from a few common ancestors (Boland et al., 2015). In Japan, pan-susceptible isolates were dominant (Ido et al., 2014). In the United Kingdom, a remarkable level of microevolution during clonal expansion of the epidemic was identified in the monophasic isolates from livestock and humans in 2005–2010 (Petrovska et al., 2016). This high level of microevolution was present with a large amount of genotypic variation that accumulated during clonal expansion, including multiple independent acquisitions of a novel prophage carrying the sopE gene and multiple deletion events affecting the phase 2 flagellin locus (Petrovska et al., 2016). Similarly, S. 1,4,[5],12:i:- isolates from Thailand contain a distinct deletion pattern in the fljBA region (Huoy et al., 2014). Similar microevolution of S. 1,4,[5],12:i:- has been described in human and animal S. Typhimurium isolates in Vietnam by WGS and phylogenetic reconstruction (Mather et al., 2018). This shows that a pandemic serotype, S. 1,4,[5],12:i:- ST34, has been introduced into Vietnam, and has reacquired a phase 2 flagellum and acquired an IncHI2 MDR plasmid.
Overall, the reported strains prevalent worldwide mainly comprise three cloned groups with independent evolution of multiclones in different regions. Some endemic clones have been found in some countries alone, suggesting that S. 1,4,[5],12:i:- isolates derived from various sources consist of multiple distinct clones. Higher heterogeneity and more clone distribution imply greater variability and adaptability, which may contribute to the success of this serotype. Microevolution associated with emergence and expansion of new epidemic clones of S. 1,4,[5],12:i:- could be determined by comparative WGS and phylogenomic analysis.
Mechanisms Involved in Adaptation to Survival of S. 1,4,[5],12:i:-
Virulence of S. 1,4,[5],12:i:-
Similar repertoire of virulence genes, such as gipA, sodC1, sopE1, and sspH1 located on prophages and spvC, pefA, and rck on a virulence plasmid in the comparison with S. Typhimurium, supports the notion that they have comparable virulence capacity in causing human salmonellosis (Hauser et al., 2010; Crayford et al., 2014; Yang et al., 2015; Seixas et al., 2016; Ngoi et al., 2018; Shippy et al., 2018). Several other virulent genes in S. 1,4,[5],12:i:- might result in different virulence level.
Toxin–antitoxin (TA) systems have been shown to be responsible for plasmid maintenance, stress management, bacterial persistence, and biofilm formation (Yang and Walsh, 2017). Widespread and virulent Salmonella serotypes, such as S. 1,4,[5],12:i:-, frequently harbor TA cassettes (Di Cesare et al., 2016). The distribution of type II TA families indicates that these elements are abundant in S. 1,4,[5],12:i:-, and the mcr-1 gene co-localizes with the hipA/hipB TA cassette only in the colistin-resistant Salmonella strain (Mastrorilli et al., 2018). These findings suggest an active role for type II TA in enhancing plasmid stability, and consequently the spread of mcr-1.
Biofilm formation in monophasic Salmonella is widely distributed in Portugal and could be one of the reasons for its dissemination in that country (Seixas et al., 2014b). Simulated gastrointestinal conditions have an effect on biofilm formation by S. 1,4,[5],12:i:- (Seixas et al., 2014a). Biofilm formation in Salmonella enables it to infect several host species and persist in the environment (Steenackers et al., 2012; Seixas et al., 2016).
SopE, one of the effector proteins translocated into host cells by the type III protein secretion system of S. Typhimurium, has been shown to bind to host RhoGTPases and activate RhoGTPase signaling, which could promote efficient bacterial entry into host cells (Rudolph et al., 1999). The sopE virulence gene was recently described in monophasic Salmonella isolates from Europe (Petrovska et al., 2016) and the United States (Elnekave et al., 2018). Expression of SopE may increase fitness of the pathogen, a possibility consistent with the observation that recent acquisition of the sopE gene by monophasic epidemic isolates was followed by an increase in sopE-positive isolates.
Antibiotic resistance of S. 1,4,[5],12:i:-
The successful spread of this serotype is allied with an escalating incidence of AMR in S. 1,4,[5],12:i:- strains. The antibiotic resistance pattern for Salmonella could be regarded as an important indicator of clonal evolution and lineage classification. Antibiotic-resistant pattern also depends on antibiotic pressure at the farm generally. R type ASSuT is prevalent in European strains from humans, pigs, and pig products (de la Torre et al., 2003; Mossong et al., 2007; Hauser et al., 2010; Laorden et al., 2010; Lucarelli et al., 2010; Hopkins et al., 2012; Wasyl and Hoszowski, 2012; Mulvey et al., 2013; Petrovska et al., 2016; Seixas et al., 2016). The ACSSuT profile is specific for the so-called Spanish clone (Echeita et al., 1999; de la Torre et al., 2003), but MDR was rare in human strains of U.S. clone in the United States (Agasan et al., 2002; Soyer et al., 2009; Switt et al., 2009). The particular antibiotic resistance pattern in Europe could be explained by the fact that the main antibiotics used in pig breeding are tetracyclines, penicillins, and sulfonamides (data from the European database of sales of veterinary antimicrobial agents).
Salmonella isolates can carry a number of different AMR genes, which may be located on the chromosome or plasmids. Most isolates from European clones with the ASSuT pattern, regardless of their origin, harbor the same AMR genes, bla TEM, strA-strB, sul2, and tet(B), which are present on a chromosomal resistance island (Hopkins et al., 2010; Lucarelli et al., 2010; Antunes et al., 2011; Seixas et al., 2016). Two mutually exclusive resistance mechanisms located in different regions of the genome, RR1 and RR2 (Lucarelli et al., 2012) and RR3 (Garcia et al., 2016), confer the ASSuT phenotype in European isolates. These resistance regions RR1 and RR2 are both surrounded by IS26 elements and sequence comparative analysis shows 99% sequence identity with a region of plasmid pO111_1 from Escherichia coli (Lucarelli et al., 2012). As for RR3, a 28-kb region is embedded between the loci STM2759 and iroB, replacing the DNA located in between, including the fljBA operon. It encompasses the genes bla TEM −1, strA-strB, sul2, and tet(B) responsible for the R-type ASSuT located on IncH1 plasmids, which is an example of the stabilization of new plasmid-derived genetic material due to integration into the bacterial chromosome (Garcia et al., 2016). It is suggested that a common ancestor harboring an IS26 in this position, where IS26-mediated homologous recombination events occur, is responsible for acquisition of the new region present in the chromosome of S. 4,[5],12:i:-/ASSuT/STYMXB.0131 07-2006, and deletion of the second phase flagellar antigen genes (Garcia et al., 2016).
In addition to ASSuT, S. 1,4,[5],12:i:- could be resistant to further antimicrobial compounds (quinolone, cefotaxime, colistin, gentamicin, and chloramphenicol). Quinolones are key antibiotics for treating invasive Salmonella infections in humans. MDR analysis of S. 1,4,[5],12:i:- isolates from swine in the United States Midwest observed genetic polymorphisms, potentially leading to resistance to quinolones, for all three resistance mechanisms: mutations in the gyrA–gyrB and parC–parE gene pairs, acquired PMQR genes, and altered expression of efflux pumps and porin diffusion channels (Elnekave et al., 2018). A study on clinical isolates from Italy in 2013–2014 showed that S. 1,4,[5],12:i:- with sole resistance to other antimicrobials such as fluoroquinolones and nalidixic acid was detected in several strains from the European lineage (Rodriguez et al., 2012; Cito et al., 2016). Fluoroquinolone resistance-conferring genes qnrS1 (from three patients) and aac(6′)lb-cr (from one patient) were also detected in MDR S. 1,4,[5],12:i:- ST34 in Australia (Arnott et al., 2018).
Extended-spectrum β-lactamase (ESBL)-producing Salmonella is considered a serious concern to public health worldwide. The presence of ESBLs in monophasic S. Typhimurium isolates from Germany has been reported, and its emergence seems to have mainly resulted from the recent spread of isolates belonging to the predominant European clone ST34 (Rodriguez et al., 2012). This clone has acquired different IncI1 plasmids harboring the bla CTX-M-1 gene within different food-producing animals (mainly swine, but also cattle and sheep). In addition, an ST108 IncI1 plasmid encoding a bla CTX-M-1 gene had disseminated across multiple genera, S. 1,4,[5],12:i:-, E. coli, and Klebsiella pneumoniae resistant to cefotaxime on a United Kingdom pig farm; an example of horizontal gene transfer of the bla CTX-M-1 gene (Freire Martin et al., 2014). The bla CTX-M-14 gene was previously reported in a monophasic isolate collected from swine in Portugal (Clemente et al., 2013), and a CTX-M-14-producing isolate of this clone was also recovered from a human clinical sample (Fernandez et al., 2016).
The plasmid-mediated colistin resistance gene, mcr-1, has been detected around the world (Campos et al., 2016; Doumith et al., 2016; Li et al., 2016) since its emergence in China in late 2015 (Liu et al., 2016). mcr-1 has been located on different plasmids in pig-associated MDR, copper-tolerant S. 1,4,[5],12:i:- ST34 clones from human and pork products since at least 2011 (Campos et al., 2016; Carroll et al., 2018). Using genomic surveillance, the presence of novel colistin resistance gene mcr-3 indicated that MDR S. 1,4,[5],12:i:- ST34 has established endemicity in Australia (Arnott et al., 2018).
Overall, the published data available on the AMR genes found in S. 1,4,[5],12,i:- led us to understand the evolution of MDR S. 1,4,[5],12,i:- strains and the natural history of the AMR gene clusters found in these strains. The acquisition of the resistance genes encoding ASSuT in the S. 1,4,[5],12,i:- isolates occurred with the loss of phase 2 flagella expression. It represents an example of the stabilization of new plasmid-derived material into the bacterial chromosome. In addition, S. 1,4,[5],12:i:-, through lacking phase 2 flagella, evolved to acquire the ASSuT resistance genes, which would help the bacteria to spread predominantly in European pig farms and industrial chains.
Heavy metal tolerance of S. 1,4,[5],12:i:-
Heavy metal tolerance might be another important reason for the prevalence and success of different clones of S. 1,4,[5],12:i:-. The resistance pattern to heavy metals could be used as another typing method besides WGS or antibiotic resistance pattern. Heavy metal compounds for growth promotion and therapeutic treatment, like zinc and cooper, have been used in pig and poultry production to promote animal growth and increase feed efficiency (Mastrorilli et al., 2018). In fact, the emergence of S. 1,4,[5],12:i:- clones causing human infections has been associated particularly with pig production with higher copper concentrations in feed (European Food Safety Authority, 2010; Hauser et al., 2010; Hopkins et al., 2010; Antunes et al., 2011; Wales and Davies, 2015). A time-dependent appearance of heavy metal tolerance genes in Italy around 2011 probably evolved under the selective pressure exerted by the rise in heavy metal concentrations in pig farming environments (Mastrorilli et al., 2018). Furthermore, genes conferring heavy metal tolerance may coexist with antibiotic-resistant genes on the same genetic element (e.g., plasmid), which could further promote co-dissemination and resistance (Yang et al., 2018). Therefore, toxic concentrations of heavy metal in the food–animal farm environment might aid the persistence of pathogens harboring metal and antibiotic resistance genes.
A widespread presence of heavy metal tolerance gene cassettes has been observed. Most of the strains possessed genes expected to confer resistance to copper (pcoA–pcoD) and silver (silA–silE) (Mastrorilli et al., 2018). The acquisition of these heavy metal tolerance gene clusters possibly promote diverse environmental adaptation: (i) bacterial motility in environments enriched with this metal (Mastrorilli et al., 2018); (ii) survival in diverse metal-contaminated settings (Mourao et al., 2016); and (iii) survival in anoxic conditions (Mourao et al., 2015).
Besides, it has been shown that copper is involved in macrophage defense against Salmonella infections (Osman et al., 2010; Achard et al., 2012), highlighting a mechanism for sil/pco genes to provide an additional fitness benefit to pathogenic organisms. The presence of additional heavy metal resistance genes may further contribute to the natural ability of S. 1,4,[5],12:i:- to escape the metal-mediated antimicrobial response of human macrophages (Kapetanovic et al., 2016).
Conclusions
S. 1,4,[5],12:i:- has successfully spread worldwide with high infection rates and broad antibiotic resistance, and diverse clones have been found in different areas with pig observed as an important reservoir. Moreover, the antibiotic resistance and heavy metal tolerance enable the serotype to adapt better to the high-abundance residues of antibiotics and heavy metals in the environment, which might elucidate the recently successful propagation and dissemination of this important pathogen. More study is still needed to reveal the association among different S. 1,4,[5],12:i:- clones, and the path of the evolution and transmission of this pathogen.
Footnotes
Supplementary Material
Supplementary Data
Supplementary Figure S1
Supplementary Table S1
Funding Information
The project was supported by the National Natural Science Foundation of China (31871899) and the National Key Research and Development Program of China (2017YFC1601502 and 2017YFC1601004).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.