
Abstract
Clostridium perfringens (CP) is a foodborne pathogen. The bacterium can also inhabit human gut without symptoms of foodborne illness. However, the clinical symptoms of long-term inhabitation have not been known yet. Therefore, the objective of this study was to elucidate the relationship between intestinal CP and other internal organs. Phosphate-buffered saline (PBS) and CP were orally injected into 5-week-old (YOUNG) and 12-month-old C57BL6/J (ADULT) mice. Gene expression levels related to inflammation (tumor necrosis factor-α [TNF-α], interleukin [IL]-1β, and IL-6) and oxidative stress (superoxide dismutase [SOD]1, SOD2, SOD3, glutathione reductase [GSR], glutathione peroxidase [GPx]3, and catalase [CAT]) responses were evaluated in the brain, small intestine, and liver. In addition, apoptosis-related (BCL2-associated X [BAX]1 and high-mobility group box-1 [HMGB1]) and brain disorder-related genes (CCAAT-enhancer-binding protein [C/EBP]-β, C/EBPδ, C/EBP homologous protein [CHOP], and amyloid precursor protein [APP]) as brain damage markers were examined. The protein expressions in the brain were also measured. Gene expression levels of inflammation and oxidative stress responses were higher (p < 0.05) in brains of CP-YOUNG and CP-ADULT mice, compared with PBS-YOUNG and PBS-ADULT, and the gene expression levels were higher (p < 0.05) in brains of CP-ADULT mice than CP-YOUNG mice. Apoptosis-related (BAX1 and HMGB1) and brain disorder-related genes (C/EBPβ, C/EBPδ, CHOP, and APP) were higher (p < 0.05) in brains of CP-challenged mice, compared with PBS-challenged mice. Even oxidative stress response (GPx and SOD2), cell damage-related (HMGB1), and β-amyloid proteins were higher (p < 0.05) in brains of CP- than in PBS-challenged mice. C/EBP protein was higher (p < 0.05) in CP-YOUNG, compared with PBS-YOUNG mice. However, these clinical symptoms were not observed in small intestine and liver. These results indicate that although asymptomatic intestinal CP do not cause foodborne illness, their inhabitation may cause brain inflammation, oxidative stress, apoptosis, and cell damage, which may induce disorders, especially for the aged group.
Introduction
C lostridium perfringens (CP) is major foodborne pathogen, and the symptoms caused by foodborne CP infection are abdominal cramps, diarrhea, vomiting, and fever (Johnson et al., 2007; Lentino, 2016). CP can also exist in intestines without pathogenic symptoms. However, the clinical impact of the asymptomatic inhabitation on other organs has not been studied, especially for the long-term inhabitation in intestines.
Recently, a number of researchers have provided scientific evidences, supporting the relationship between the intestinal microbiota and human health. The association between gut microbiota and diseases such as inflammatory disorders and even neuropsychiatric disorders has been discussed by many scientists (Guarner and Malagelada, 2003; Burisch et al., 2014; Wang and Kasper, 2014; Boulangé et al., 2016; Schneiderhan et al., 2016; Shen and Wong, 2016). In a study by Mitsuoka (1992), the prevalence of bifidobacteria in human feces decreased, but CP levels rapidly increased in the aged. One of the diseases caused in the aged is a brain disorder, and the causes of the brain disorder have not yet been fully clarified. In addition, many scientists suggested the linkage between gut microbiota and brain, called “gut–brain axis (GBA)” (Rhee et al., 2009). Taken together, there is a possibility that intestinal CP is related to brain disorders and disease in other organs as well.
Therefore, the objective of this study was to elucidate if commensal CP in intestines is related to any clinical symptoms in human body, especially in the brain.
Materials and Methods
Animals
Five-week-old (YOUNG) and 12-month-old (ADULT) C57BL/6J male mice were purchased from Japan SLC, Inc. (Hamamatsu, Japan), and were randomly divided into the phosphate-buffered saline (PBS)- and C. perfringens (CP)-challenged groups (five mice/group). Thus, the challenge groups were PBS-YOUNG, PBS-ADULT, CP-YOUNG, and CP-ADULT. YOUNG mice (two to three mice/cage) and ADULT mice (one mouse/cage) were separately housed under controlled temperature (21°C ± 1°C), humidity (50–60%), and on a 12-h dark/light cycle. For the 1-week acclimation, the mice received sterile standard chow and acidic water (pH 3, adjusted with 1 M HCl) ad libitum. All animal procedures were approved by the Animal Ethical Committee of Korea University Institutional Animal Care and Use Committee (KUIACUC-2016-172).
Depletion of gut commensal microflora and CP challenge
The mice were treated with a mixture of antibiotics in drinking water for 10 d before CP challenge as described by Fagarasan et al. (2002). Seven CP strains (NCCP10347 [type A], NCCP10858 [type B], NCCP10846 [type B], NCCP10920 [type D], NCCP10976 [type A], KCCM40946 [type A], and KCCM12098 [type A]) were used. Each CP stock culture (300 μL) was inoculated into cooked meat medium (CMM; Oxoid Ltd., Basingstoke, United Kingdom), and anaerobically incubated at 37°C for 24 h. Then, 3 mL of each CP culture was added to 30 mL Duncan and Strong medium (Duncan and Strong, 1968) and anaerobically incubated at 37°C for 48 h. Each CP culture was harvested by centrifugation (1,912 × g, 4°C, and 15 min), resuspended in 1 mL PBS, and then seven CP strains were mixed and centrifuged (1,912 × g, 4°C, and 15 min), followed by resuspension in 7 mL PBS. The aliquots (200 μL) of the CP mixture (9 log colony-forming unit [CFU]/mL) or PBS were administered to YOUNG (CP-YOUNG) and ADULT (CP-ADULT) mice by oral gavage three times a week for 8 weeks to make CP inhabit the gut.
Body weight and microbiota in feces
The mice were weighed, and their feces were collected on the arrival day (day 0), after antibiotics treatment (day 10), and on days 17, 24, 31, and 38. The collected feces samples were evaluated for total anaerobic bacteria and CP counts in brain–heart infusion broth (BHI; Difco™; Sparks, MD) and tryptose sulfite cycloserine (TSC) agar (Oxoid Ltd.), respectively. After incubation at 37°C for 24 h under anaerobic conditions, the colonies on BHI and TSC agar were counted.
Euthanasia
After 8 weeks of CP and PBS oral administration, the mice were killed for further experiments. Before killing, the mice were fasted for 18 h with only sterile tap water provided. They were then anesthetized with ethyl ether, and the mice killed by blood collection. Liver, spleen, small intestine, and large intestine were quickly removed and weighed to calculate relative weights (organ weight/body weight). The brain, intestine, and liver samples were then taken and placed on dry ice before storage at −70°C to minimize RNA and protein degradation. These organs were used for measuring gene and protein expression levels.
Gene expression of ageing factors
The frozen samples (10 mg) were placed in 2-mL microcentrifuge tubes, containing one stainless steel bead (5-mm diameter; Qiagen) on dry ice, followed by the addition of 1 mL PBS to lyse the tissue. The samples were sonicated using a tissue lyser (TissueLyser LT; Qiagen) for 5–10 min at 50 Hz until they were completely homogenized. The samples were centrifuged at 15,814 × g for 5 min to remove tissue lysates, and the supernatants were transferred to new microtubes and stored at −20°C until analysis. The RNAs in tissues of the mice were extracted using the RNeasy mini kit (Cat. No. 74106; Qiagen) and RNeasy lipid tissue mini kit (Cat. No. 74804; Qiagen) according to the manufacturer's instructions. The complementary DNA (cDNA) was synthesized from the messenger RNA (mRNA) using the QuantiTect reverse transcription kit (Qiagen) according to the manufacturer's instructions. The Rotor-Gene SYBR Green polymerase chain reaction kit (Qiagen) using Rotor-Gene Q (Qiagen) was used for the quantification of mRNA expression. To characterize the mRNA gene expression changes in YOUNG and ADULT mice after CP challenge, the expression levels of inflammatory cytokines-related genes (tumor necrosis factor-α [TNF-α], interleukin [IL]-1β, and IL-6) and oxidative stress-related genes (superoxide dismutase [SOD]1, SOD2, SOD3, glutathione reductase [GSR], glutathione peroxidase [GPx]3, and catalase [CAT]) were measured in the brain, intestine, and liver. The primer sequences used in this study are described in Table 1. β-actin gene was used as the reference gene for normalization, and relative gene expressions were calculated against the gene expression levels of PBS-YOUNG. The relative target gene expression was calculated, using the 2−ΔΔCt method. The threshold was set to 0.1, and Ct values were calculated as described by Pfaffl et al. (2002).
Primer Sequences of Inflammation and Oxidative Stress-Related Genes Used for Real-Time Polymerase Chain Reaction
CAT, catalase; GPx, glutathione peroxidase; GSR, glutathione reductase; IL, interleukin; SOD, superoxide dismutase.
Gene expression of brain damage markers
Tight junction protein-related genes (occludin, claudin, and TJP) were used to evaluate the changes in tight junction gene expression in the brain and intestine. In addition, cell damage-related (BCL2-associated-X [BAX]1 and high-mobility group box-1 [HMGB1]) and brain disorder-related (CCAAT-enhancer-binding protein [C/EBPβ], C/EBPδ, C/EBP homologous protein [CHOP], and amyloid precursor protein [APP]) gene expressions were measured in the brain. The primer sequences used in this study are described in Table 2.
Primer Sequences of Tight Junction-, Cell Damage-, and Brain Disorder-Related Genes Used for Real-Time Polymerase Chain Reaction
APP, amyloid precursor protein; BAX1, BCL2-associated X-1; C/EBP, CCAAT-enhancer-binding protein; CHOP, C/EBP homologous protein; HMGB1, high-mobility group box-1; TJP, tight junction protein.
Protein expression in the brain
Ten milligrams of frozen brain tissue samples were placed in 2-mL microcentrifuge tubes containing one stainless steel bead on dry ice, followed by the addition of 600 μL PRO-PREP™ solution to lyse the tissue for western blotting. The samples were sonicated, using a tissue lyser for 5–10 min at 50 Hz until tissue samples were completely homogenized. After sonication, the samples were centrifuged at 15,814 × g at 4°C for 5 min to remove tissue lysates, and the supernatants of the samples were transferred to new microtubes. The supernatants of the samples were stored at −20°C until analysis. For determination of total protein concentration levels, a DC protein assay I kit (Bio-Rad Laboratories, Inc., Hercules, CA) was used according to the manufacturer's instructions. Aliquots (17 μg) of extracted protein were separated, using electrophoresis (200 V, 30 min) on 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE), and transferred to polyvinylidene difluoride (PVDF) membranes (GE Healthcare Life Sciences, Marlborough, MA) at 60 V for 2 h and 30 min on ice. The protein probes were incubated with primary antibodies against β-actin (sc-81178, 1:1000), IL-6 (sc-57315, 1:500), occludin (sc-271842, 1:500), claudin (sc-166338, 1:500), TJP (ab191133, 1:1000), SOD (sc-130345, 1:500), GPx (sc-133160, 1:500), C/EBP (sc-7351, 1:200), β-amyloid (sc-28365, 1:100; all Santa Cruz Biotechnologies, Dallas, TX), and HMGB1 (ab18256, 1:2500; Abcam, Cambridge, United Kingdom) in Tris-buffered saline solution (TBST; 1 × Tris-glycine buffer and 0.1% Tween-20 in distilled water), containing 5% skim milk powder. The membranes were washed 3 times with TBST and incubated at room temperature for 1 h with a secondary antibody (goat anti-mouse IgG-HRP, sc-2005; Santa Cruz Biotechnologies), diluted 1:5000 in TBST containing 5% skim milk powder. After incubation, the membranes were washed three times with the TBST, and immunoreactive bands were visualized using the Amersham ECL prime western blotting detection reagent (GE Healthcare Life Sciences). The protein bands were captured using a biomolecular imaging system (LAS 3000; Fujifilm, Tokyo, Japan), and the intensities of the immunoreactive bands were quantified using the GelQuant software v.2.7 (DNR Imaging Systems Ltd., Jerusalem, Israel). β-actin was used as a reference protein for normalization of the data.
Statistical analysis
All values are presented as mean ± standard error. Statistical analysis was performed with the general linear model of the SAS® version 9.2 (SAS Institute, Inc., Cary, NC). Least squares means among the fixed effects were compared by a pairwise t-test at α = 0.05.
Results and Discussion
Depletion of CP and physiological change
Before antibiotic treatments, 2.7 ± 0.5 log CFU/g of CP were detected in fecal samples of mice. After administering antibiotics for 10 d to deplete the gut commensal microflora, the CP cell counts in fecal samples were below the detection limit (1.4 log CFU/g), and the young (YOUNG) and adult (ADULT) mice were then orally injected with CP or PBS. After CP challenge for 1 week, CP cell counts in CP-challenged mice were 5.2 ± 2.0 log CFU/g in CP-YOUNG and 2.3 ± 0.9 log CFU/g in CP-ADULT, but no CP were detected in PBS-challenged mice (Table 3). This indicates that injected C. perfringens settled down in small intestine. After 24 d and until 38 d, CP were detected in PBS-challenged mice, but the concentrations were lower (p < 0.001) than CP-challenged mice in YOUNG group. It can be an evidence for supporting C. perfringens existence in intestine (Mitsuoka, 1992).
Clostridium perfringens Counts (Mean ± Standard Deviation) in Fresh Fecal Samples, Enumerated on Tryptose Sulfite Cycloserine Agar
CP-ADULT, Clostridium perfringens-challenged adult mice; CP-YOUNG, C. perfringens-challenged young mice; ND, not detected; PBS-ADULT, phosphate-buffered saline-challenged adult mice; PBS-YOUNG, phosphate-buffered saline-challenged young mice.
On day 59 after CP challenge, body weight differences were not significant between PBS- and CP-challenged YOUNG and ADULT groups. The final body weights were 26.52 ± 0.29 g and 25.36 ± 0.81 g for PBS-YOUNG and CP-YOUNG groups, and 31.03 ± 1.99 g and 28.18 ± 2.14 g for PBS-ADULT and CP-ADULT groups, respectively (Table 4). In addition, no clinical pathological symptoms such as loss of appetite, loose stool, and so on, were observed. After the mice were killed, the relative weights of the liver, spleen, small intestine, and large intestine were not significantly different between the PBS- and CP-challenged mice, regardless of age (Table 4). This indicates that CP challenge may not affect physical organ damage in mice.
Liver and Spleen Weights (Mean ± Standard Deviation) and Their Relative Weight (Mean ± Standard Deviation) of Young and Adult Mice
bw, body weight; CP-ADULT, Clostridium perfringens-challenged adult mice; CP-YOUNG, C. perfringens-challenged young mice; PBS-ADULT, phosphate-buffered saline-challenged adult mice; PBS-YOUNG, phosphate-buffered saline-challenged young mice.
Effects of CP on brain
Inflammation
TNF-α, IL-1β, and IL-6 are representative proinflammatory cytokines. The transcriptome levels of the cytokines such as TNF-α and IL-1β were not different between CP- and PBS-challenged mice, but the difference (p = 0.0062) was detected for IL-6 in ADULT groups (Fig. 1A). IL-6 protein expression was also examined, and more IL-6 protein was expressed (p = 0.0704) in CP-challenged mice than in the PBS-challenged mice in ADULT groups (Fig. 1B, Supplementary Fig. S1). Of the tight junction genes, occludin and claudin gene expressions were not different, only TJP gene expression levels were higher (p < 0.05) in CP-ADULT than in PBS-ADULT mice (Fig. 2A). In addition, in protein level, occludin and claudin expressions were not different between PBS- and CP-challenged mice, but TJP protein expression was higher (p < 0.05) in CP-challenged mice, compared with PBS-challenged mice, regardless of age (Fig. 2B, Supplementary Fig. S2). These results suggest that CP in the gut microbiota may increase proinflammatory cytokines, and it can cause low-grade inflammation as suggested by Minciullo et al. (2017). The inflammation over long term then possibly affects tight junction in brain.

Relative mRNA
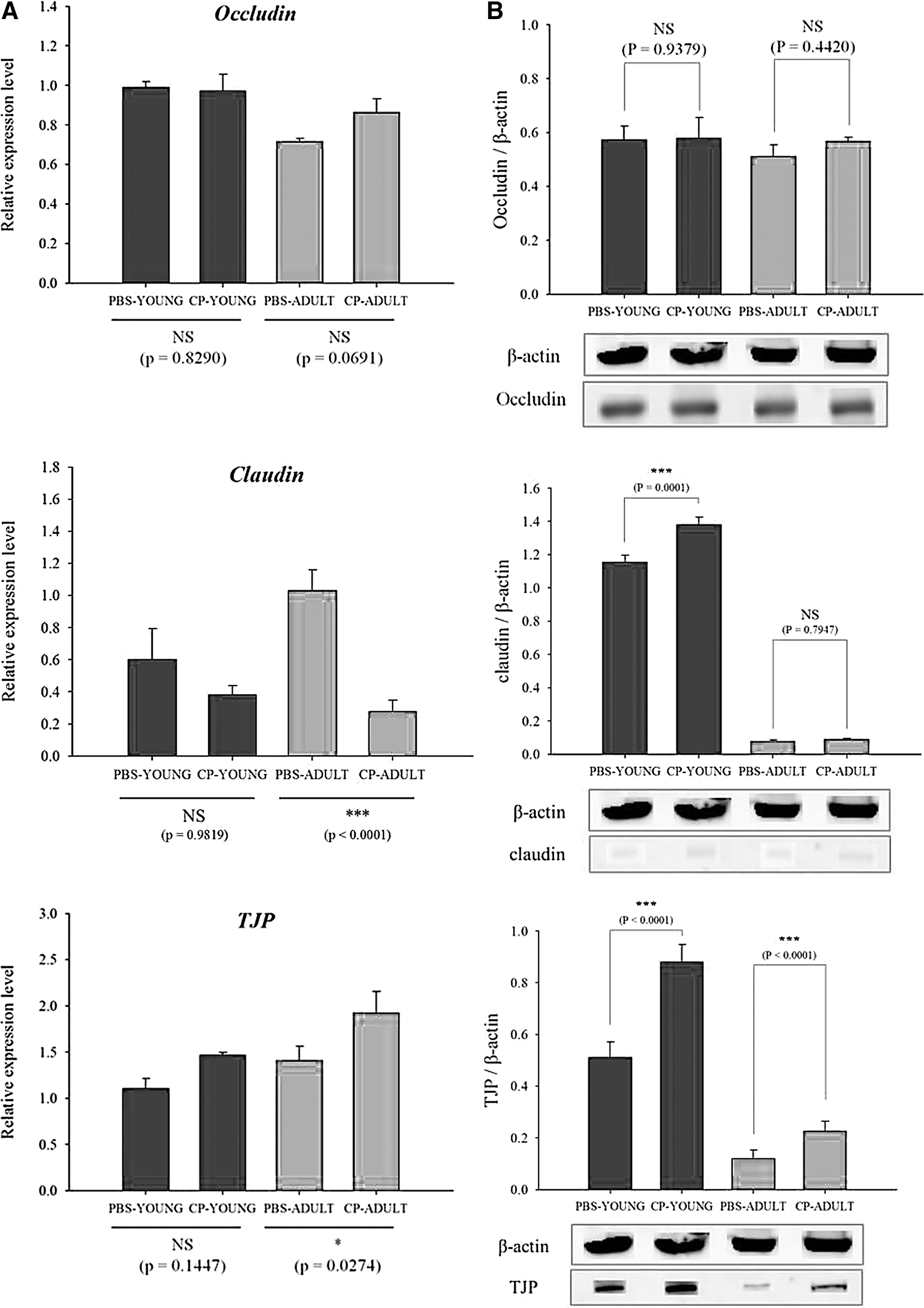
Relative mRNA
Oxidative stress response
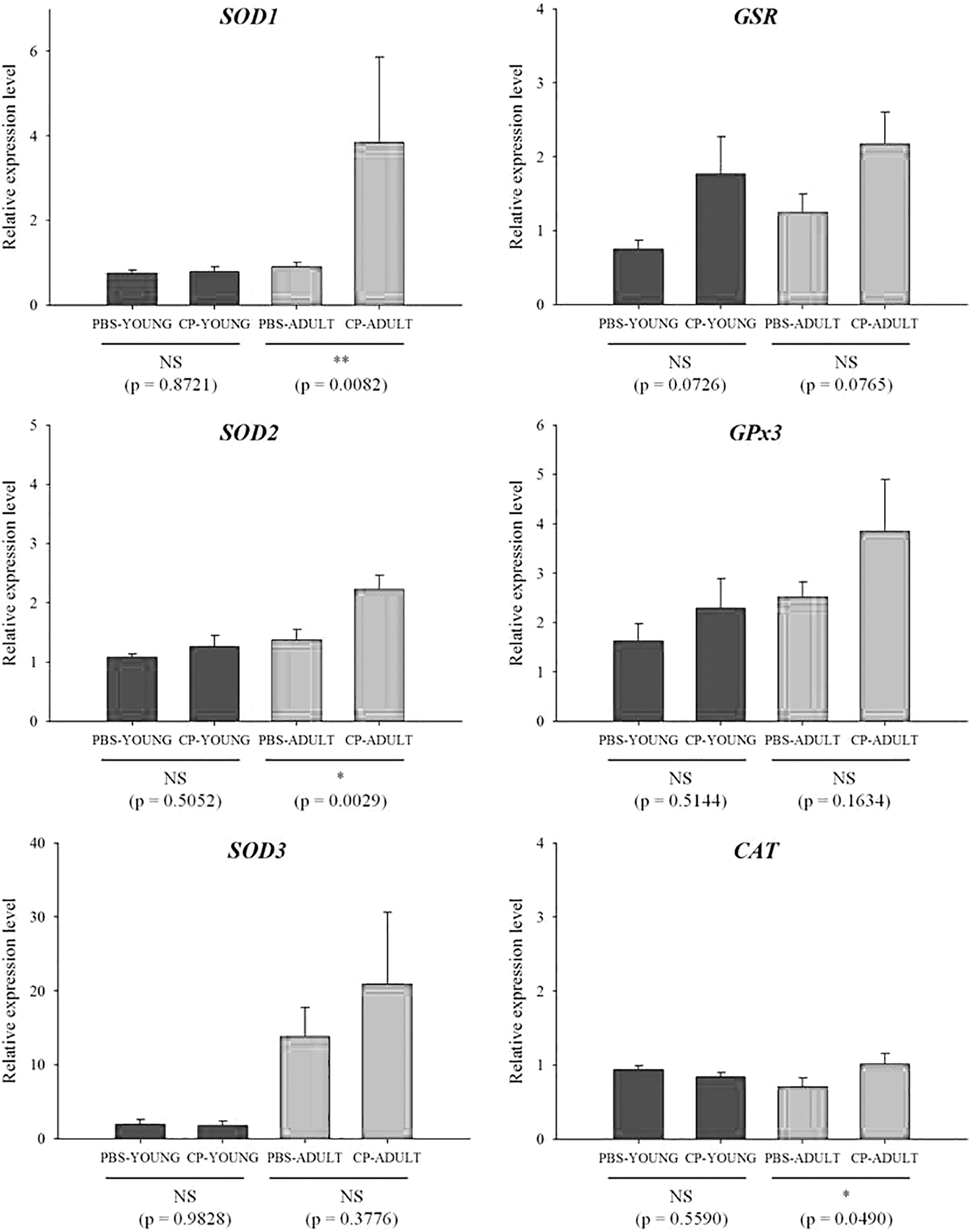
Reactive oxygen species (ROS) can increase dramatically when the host is exposed to stressful environments (Devasagayam et al., 2004). ROS play roles not only in apoptosis, but also programmed cell death by oxidative stress (Rada and Leto, 2008; Sosa Torres et al., 2015). Thus, the gene expression levels related to major antioxidant enzymes such as SOD, CAT, GSR, and GPx, which neutralize ROS, were measured. SOD1, GSR, GPx3, and CAT gene expressions were generally higher in CP-challenged than in PBS-challenged mice, especially in CP-ADULT (Fig. 3). In addition, at the protein level, GPx and SOD2 were higher (p < 0.05) in CP-ADULT than in PBS-ADULT mice (Fig. 4, Supplementary Fig. S3). According to the study by Van Raamsdonk and Hekimi (2008), oxidative damage initiated by ROS is a major contributor for aging. Taken together, these results suggest that inflammation and ROS production in the brain can be derived from the presence of CP in the intestine, especially in adult mice, which could induce aging and apoptosis in the brain. Thus, brain cell apoptosis was then measured.
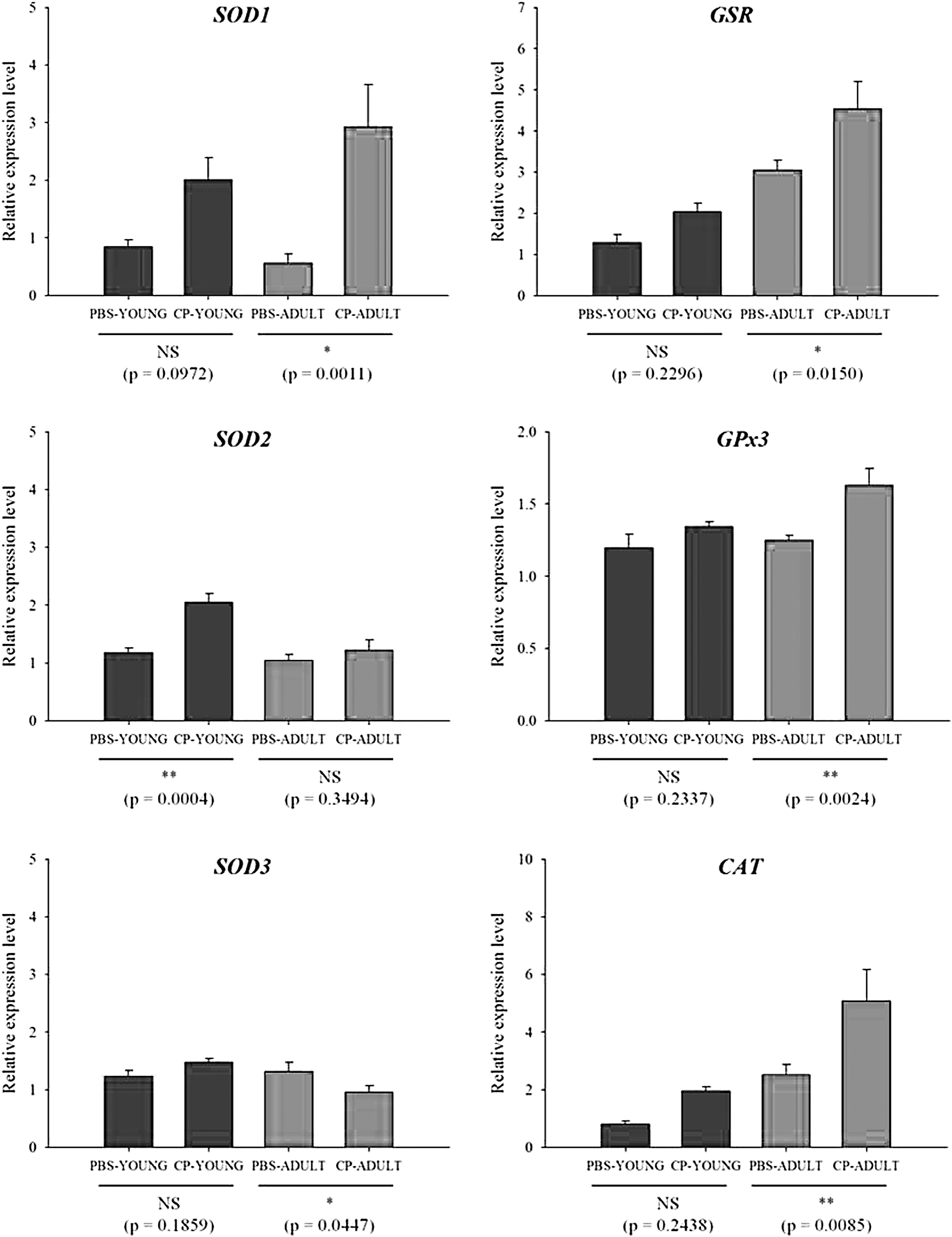
Relative mRNA expression levels related to oxidative stress in brain tissues of YOUNG and ADULT mice. Significance levels are presented as *p < 0.05, **p < 0.01, and NS. CAT, catalase; CP-ADULT, Clostridium perfringens-injected adult mice; CP-YOUNG, C. perfringens-injected young mice; GPx, glutathione peroxidase; GSR, glutathione reductase; NS, no significance; PBS-ADULT, phosphate-buffered saline solution-injected adult mice; PBS-YOUNG, phosphate-buffered saline-injected young mice; SOD, superoxide dismutase.
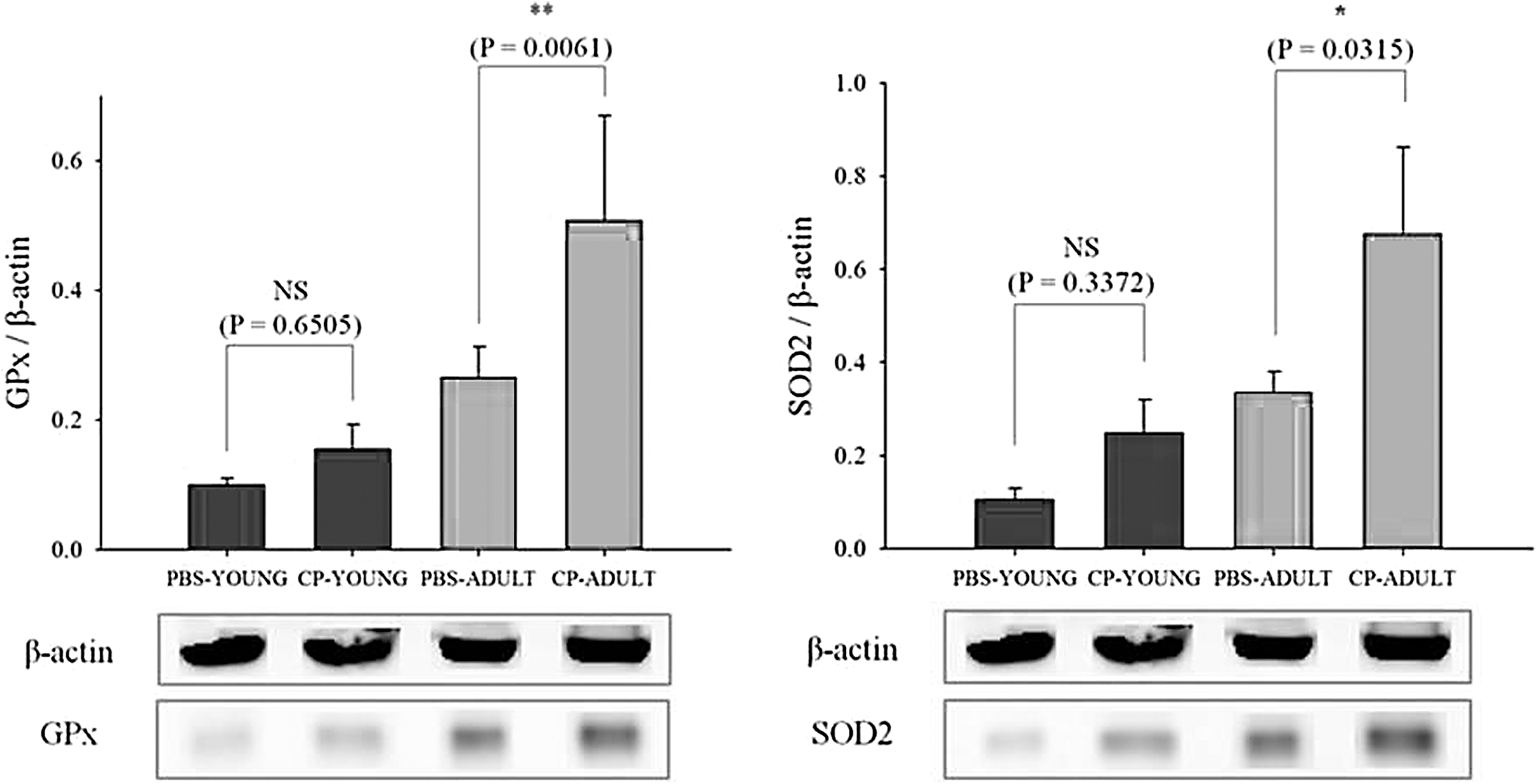
Protein expression levels related to oxidative stress in brain tissues of YOUNG and ADULT mice brain tissue. Significance levels are presented as *p < 0.05, **p < 0.01, and NS. CP-ADULT, Clostridium perfringens-injected adult mice; CP-YOUNG, C. perfringens-injected young mice; GPx, glutathione peroxidase; NS, no significance; PBS-ADULT, phosphate-buffered saline solution-injected adult mice; PBS-YOUNG, phosphate-buffered saline-injected young mice; SOD, superoxide dismutase.
Brain cell apoptosis
Apoptosis is an ordered cascade of enzymatic events that culminates in cell death, and BAX1 and HMGB1 genes are related to the apoptosis of brain cells. BAX1 gene expression is a signal of apoptosis initiation, and the HMGB1 gene controls the cellular apoptosis checkpoint during inflammation (Pawlowski and Kraft, 2000; Zhu et al., 2015). In our study, BAX1 and HMGB1 gene expression levels in the brain were higher (p < 0.05) in CP-challenged mice, compared with PBS-challenged mice (Fig. 5A). In addition, HMGB1 protein expression level in the brain were also higher (p < 0.01) in CP-ADULT than in PBS-ADULT mice, and the expression levels were higher (p < 0.01) in ADULT than in YOUNG groups (Fig. 5B, Supplementary Fig. S4). These results suggest that CP in the intestine may induce brain cell apoptosis. Although mRNA expressions of HMGB1 does not regulate directly its protein, HMGB1 mRNA expression interacts with various molecule-associated inflammation and apoptosis. Thus, the gene and protein expressions could be evidence for the cell apoptosis of the brain. Pawlowski and Kraft (2000) suggested that the inappropriate activation of apoptosis may be a contributing factor in the brain. Thus, the effect of intestinal CP on brain disorder was further analyzed.
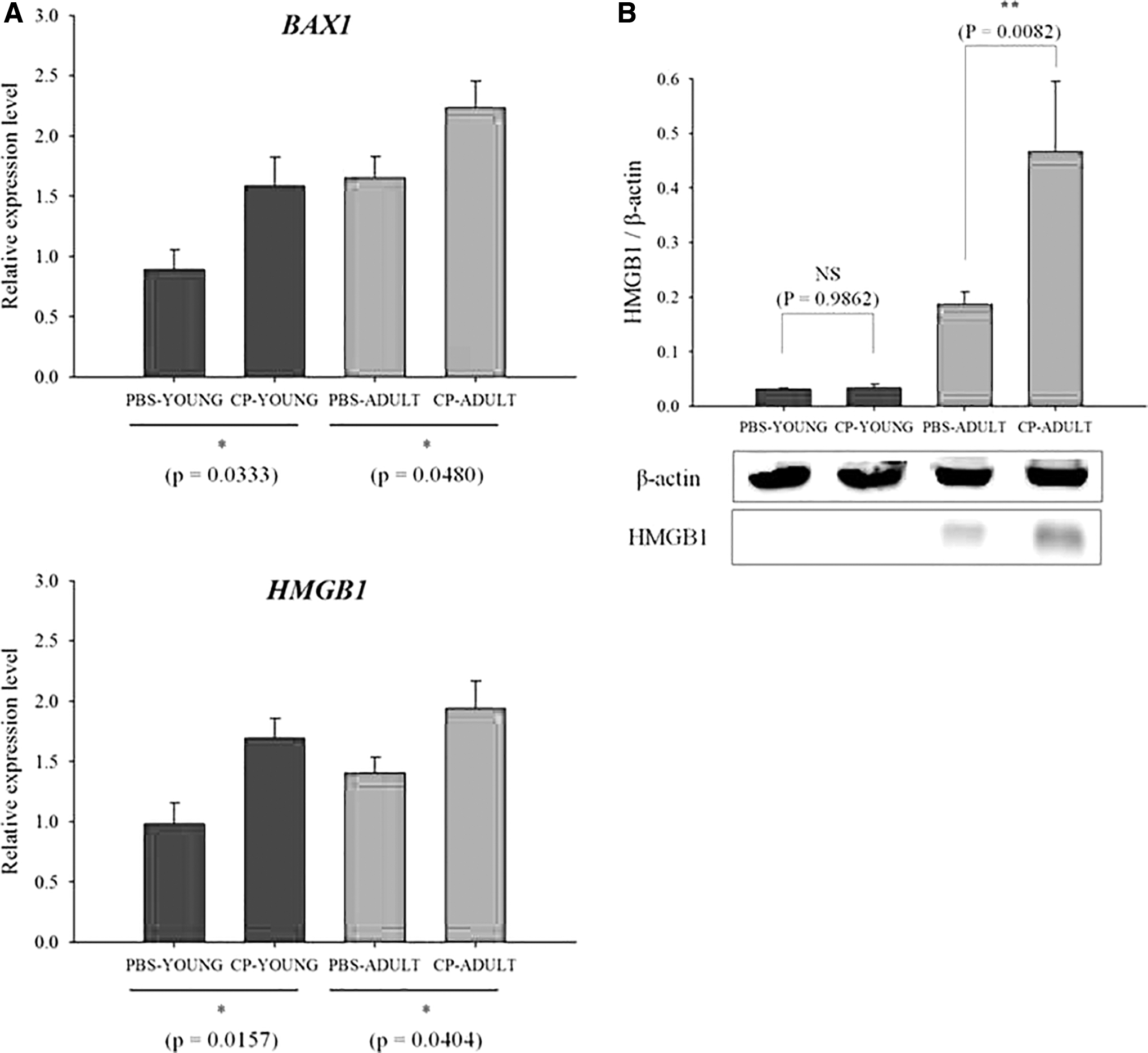
Relative mRNA
Brain disorder
Increased expression of the C/EBP family could accelerate aging and be a potential risk factor for brain disorder because C/EBP regulates the expression of many proinflammatory genes, which show increased expression following the injury of the aged brain (Sandhir and Berman, 2010). C/EBPβ is an important member of the C/EBP family that plays a role in cell differentiation and is a major mediator of inflammatory responses and brain injury (Cortes-Canteli et al., 2008). C/EBPδ was elevated in the brains of patients with Alzheimer's disease (Li et al., 2004), although the link between elevated C/EBPδ and Alzheimer's disease is unclear. Hence, C/EBPβ and C/EBPδ expression levels in the brain were measured in our study. C/EBPβ expression levels were not different between the PBS- and CP-challenged mice, regardless of age (Fig. 6A). C/EBPδ gene expression levels were higher in CP-challenged mice than in PBS-challenged mice, and the gene expression levels were also higher (p < 0.05) in ADULT mice than in YOUNG mice (Fig. 6A). CHOP protein also mediates CHOP through the induction of apoptosis by activation of caspase-12 (Szegezdi et al., 2003), and increased expression of CHOP gene expression has been reported to be related to transient ischemia (Paschen et al., 1998). In our study, CHOP gene expression levels were higher (p < 0.05) in CP-challenged mice than in PBS-challenged mice, regardless of age (Fig. 6A). Alzheimer's disease, the leading cause of dementia, is characterized by the accumulation of the β-amyloid peptide (Aβ) in the brain. Aβ peptide generation from sequential APP proteolysis is a crucial step in the development of Alzheimer's disease (O'Brien and Wong, 2011). Thus, APP gene expression levels were measured, and were found to be higher in CP-challenged mice than in PBS-challenged mice, and there were significantly higher (p < 0.05) expression levels of APP gene in CP-YOUNG than in CP-ADULT mice (Fig. 6A). In addition, Aβ protein levels were higher (p < 0.05) in CP-YOUNG than in PBS-YOUNG mice (Fig. 6B, Supplementary Fig. S5). Intestinal CP is not observed in the young, but the levels rapidly increased in the aged (Mitsuoka, 1992). Hence, this result may not be observed in a real situation, but it indicates that the young may be more sensitive to express Aβ protein if they are exposed to intestinal CP. In addition, it can be hypothesized that the continuous exposure may increase the possibility of Alzheimer's disease in the aged. These results indicate that CP in the intestine may influence the development of brain disorders.
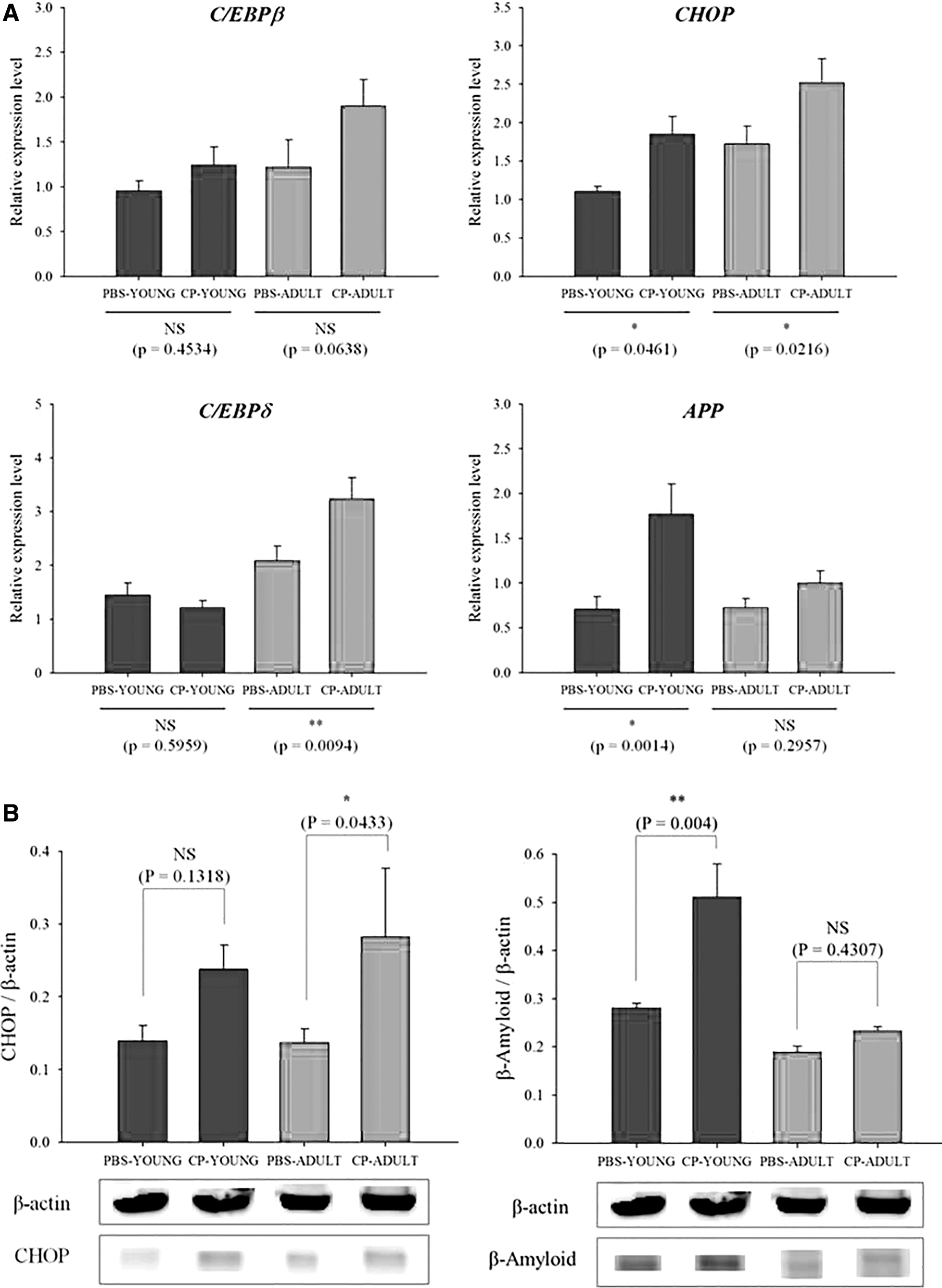
Relative
Effects on the intestine and liver
Inflammation
TNF-α, IL-1β, and IL-6 (proinflammatory cytokines) gene expression levels in the intestine were not different between the PBS- and CP-challenged mice, regardless of age (Fig. 7A). In the liver, TNF-α was not detected, and IL-1β and IL-6 expression levels were also not different between PBS- and CP-challenged mice, regardless of age (Fig. 7B). This indicates that CP in the intestine does not cause inflammation in the intestine and liver.
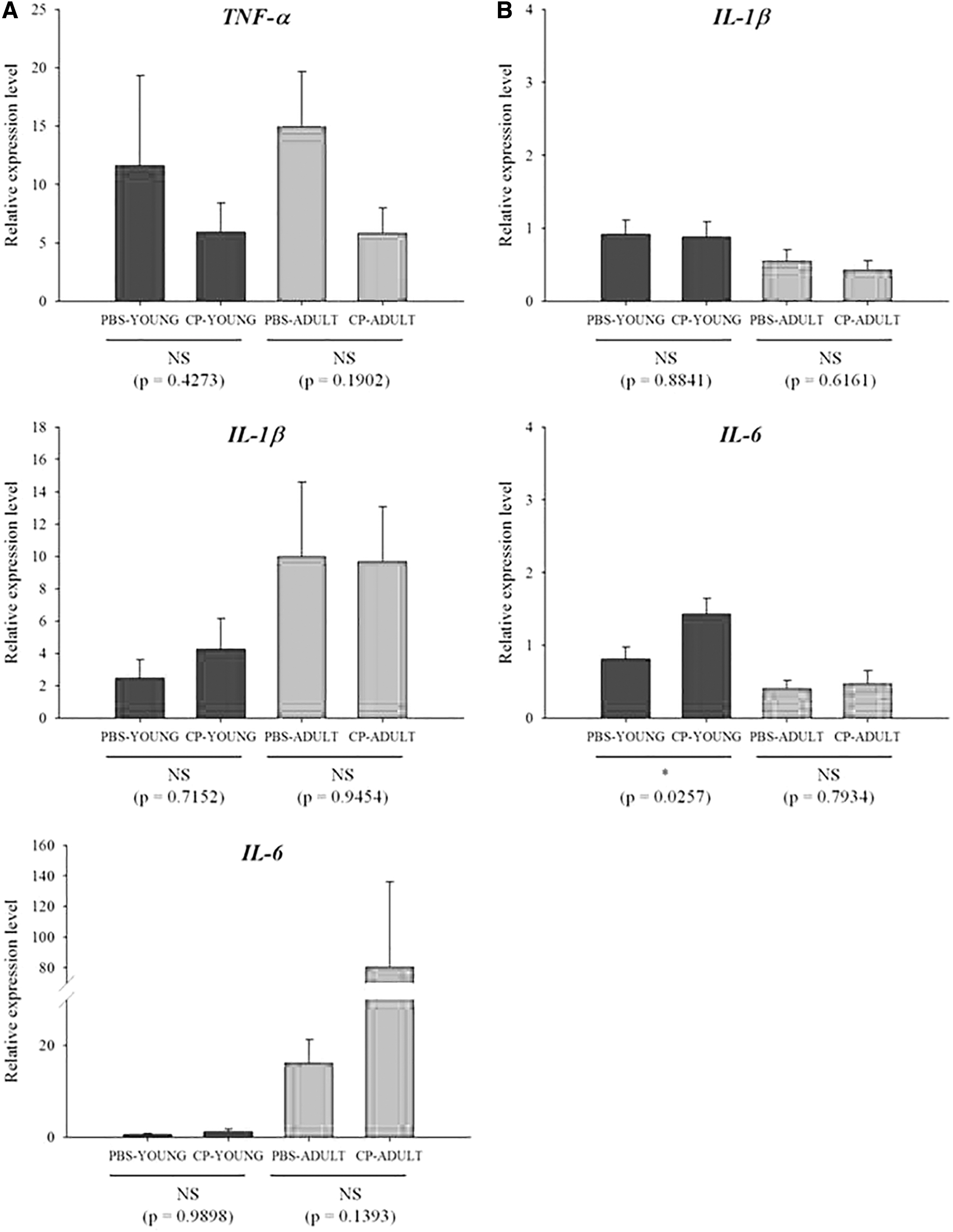
Relative mRNA expression levels related to inflammatory cytokines in intestine
Oxidative stress responses
Although few oxidative stress response gene expressions were higher in CP-ADULT than in PBS-ADULT, in general, there were no obvious differences in most oxidative stress response gene expression levels between PBS- and CP-challenged mice in the intestine and liver (Fig. 8).
Relative mRNA expression levels related to oxidative cytokines in intestine tissues of YOUNG and ADULT mice. Significance levels are presented as *p < 0.05, **p < 0.01, and NS. CAT, catalase; CP-ADULT, Clostridium perfringens-injected adult mice; CP-YOUNG, C. perfringens-injected young mice; GPx, glutathione peroxidase; GSR, glutathione reductase; PBS-ADULT, phosphate-buffered saline solution-injected adult mice; PBS-YOUNG, phosphate-buffered saline-injected young mice; SOD, superoxide dismutase.
Tight junctions
There were no significant differences in occludin and TJP expressions between the PBS- and CP-challenged mice in the intestine, regardless of age (Fig. 9). These results indicate that CP in the intestine may not induce an obvious oxidative stress response, inflammation, or tightening or loosening of tight junctions in the intestine, which could be caused by oxidative stress responses and inflammation.
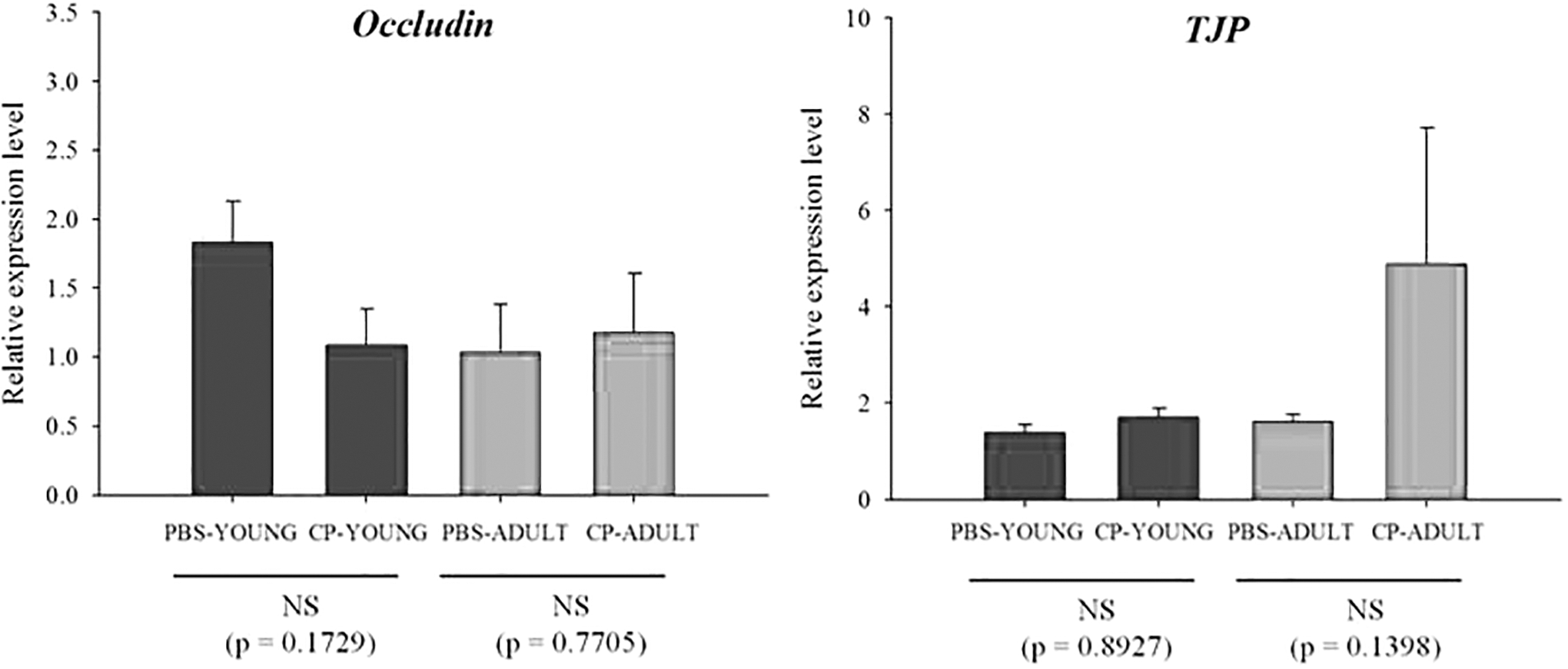
Relative mRNA expression levels related to tight junctions in intestine tissues of YOUNG and ADULT mice. CP-ADULT, Clostridium perfringens-injected adult mice; CP-YOUNG, C. perfringens-injected young mice; NS, no significance; PBS-ADULT, phosphate-buffered saline solution-injected adult mice; PBS-YOUNG, phosphate-buffered saline-injected young mice; TJP, tight junction protein.
In conclusion, CP in intestine can induce inflammation, oxidative stress, apoptosis, and damage in the brain, but it may not cause obvious pathogenic phenomena in other organs such as the intestine and the liver. ROS in the brain could be produced by certain substances related to CP in the intestine, and could induce inflammation and apoptosis in the brain. These results were more obvious in CP-ADULT than CP-YOUNG. Therefore, it can be suggested that although CP does not cause foodborne illness symptoms, CP inhabitation in intestine may induce brain disorder, especially for the aged group. However, interaction mechanisms between CP in the intestine and brain disorder should be elucidated in a further study.
Footnotes
Disclosure Statement
The authors declare no competing interests.
Funding Information
This study was supported by Wonkwang University in 2019.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.