
Abstract
Escherichia coli recovered from poultry, and extraintestinal pathogenic E. coli (ExPEC), responsible for most cases of urinary tract infection (UTI) and bloodstream infection (BSI) in humans, may share genetic characteristics, suggesting that poultry are a potential source of ExPEC. Here, we compared E. coli isolated from commercial broiler and backyard chickens (n = 111) with ExPEC isolated from patients with community- or hospital-acquired UTI or BSI (n = 149) from Southeast Brazil. Isolates were genotyped by multilocus sequence typing, tested for susceptibility to antimicrobial agents, and screened for β-lactamase genes. We found that 10 genotypes were shared among poultry and human isolates: sequence type (ST) 10, ST48, ST58, ST88, ST90, ST93, ST131, ST602, ST617, and ST1018. Thirty-five (23%) ExPEC and 35 (31%) poultry E. coli isolates belonged to the shared STs. ST58 and ST88 isolates from human and poultry sources shared identical antimicrobial resistance profiles. bla TEM-1 was the most prevalent β-lactamase gene, identified in 65 (92%) of 71 ExPEC and 29 (67%) of 43 poultry E. coli that tested positive for β-lactamase genes. Commercial broiler chicken isolates shared the extended-spectrum β-lactamase (ESBL) genes bla CTX-M-2, bla CTX-M-8, and bla SHV-2 with human isolates; backyard chicken isolates lacked ESBL genes. In conclusion, several genotypic and phenotypic characteristics were shared between human and poultry E. coli; this suggests that there is potential for transmission of E. coli and antimicrobial resistance genes from poultry to humans, perhaps through environmental contamination, direct contact, or consumption. Additional research is needed to understand the potential direction and pathways of transmission.
Introduction
Extraintestinal pathogenic Escherichia coli (ExPEC) are a subset of E. coli—distinct from nonpathogenic commensal E. coli and intestinal pathogenic E. coli—that cause extraintestinal infections and represent a serious public health concern (Russo and Johnson, 2000, 2003). In humans, ExPEC are responsible for most cases of urinary tract infection (UTI), bloodstream infection (BSI), and other community-associated and health care-associated infections (Russo and Johnson, 2000; Weiner et al., 2016). Similarly, E. coli that cause infections in poultry are termed avian pathogenic E. coli (APEC); APEC can cause colibacillosis, a disease of significant economic impact in farmed poultry (Dziva and Stevens, 2008).
While ExPEC are genetically diverse, a small number of important clonal groups—sequence type 131 (ST131), ST69, ST95, ST73, ST127, and ST10—are responsible for a large proportion of human extraintestinal E. coli infections worldwide (Izdebski et al., 2013; Maluta et al., 2014; Riley, 2014). Some of these strains produce extended-spectrum β-lactamases (ESBLs), which mediate resistance to extended-spectrum cephalosporins, and are often also resistant to other antimicrobial agents, including aminoglycosides, fluoroquinolones, and trimethoprim-sulfamethoxazole (Coque et al., 2008; Brisse et al., 2012; Adams-Sapper et al., 2013; Kim et al., 2018). These ESBL-producing and multidrug-resistant isolates are often associated with treatment failure.
Some ExPEC isolated from humans and E. coli isolated from poultry meat demonstrate similar phenotypic and genotypic characteristics, such as antimicrobial resistance patterns, antimicrobial resistance genes, virulence genes, serotype, and sequence types. Therefore, it has been hypothesized that food animals, especially poultry, are a potential source of human ExPEC (Bergeron et al., 2012; Mora et al., 2013; Maluta et al., 2014; Yamaji et al., 2018). This zoonotic potential could represent a serious public health problem (Giufrè et al., 2012; Egervärn et al., 2014; Casella et al., 2015). Brazil is a large producer and exporter of poultry, particularly chicken meat (
Materials and Methods
E. coli isolates
E. coli isolated from humans and poultry were obtained for this study. Human E. coli strains were isolated from patients with suspected community-acquired UTI (CA-UTI), hospital-acquired UTI (HA-UTI), or hospital-acquired BSI (HA-BSI) from September 2014 to July 2015, at the University Hospital Faculty of Medicine of Ribeirão Preto, University of São Paulo, Ribeirão Preto, SP, Brazil. All patients resided within 150 km of the hospital. Isolates were identified by the Vitek2® automated microbial identification system (bioMérieux, Marcy-l'Étoile, France). Poultry E. coli strains were obtained from the Bacteriology Laboratory in the Veterinary Pathology Department, São Paulo State University (UNESP), Jaboticabal, SP, Brazil. These strains were isolated from January to April 2014, in the Ribeirão Preto region, SP, Brazil from the cloaca of (1) commercial broiler chickens from two slaughterhouses, and (2) backyard chickens from seven small backyard farms (mostly family farms in urban and periurban centers). The slaughterhouses and backyard farms were within 100 km from the hospital. Poultry isolates were recovered according to a previous work (Borges et al., 2017).
Multilocus sequence typing
MLST was performed following the Achtman scheme (
Antimicrobial susceptibility testing
Antimicrobial susceptibility testing (AST) for human E. coli isolates was performed with the Vitek2 system. AST for poultry E. coli isolates was performed by standard disk diffusion, according to the testing and interpretive criteria of the Clinical and Laboratory Standards Institute—CLSI (CLSI, 2015), which included the following classes and antimicrobial agents: aminoglycosides (amikacin, gentamicin), β-lactams (ampicillin, amoxicillin-clavulanic acid, cefuroxime, ceftriaxone, cefepime, ertapenem, and imipenem), fluoroquinolones (ciprofloxacin), folate pathway inhibitors (trimethoprim-sulfamethoxazole), and nitrofurans (nitrofurantoin). E. coli ATCC 25922 from the American Type Culture Collection was used as a reference strain. Isolates with intermediate susceptibility were classified as nonsusceptible. Multidrug resistance (MDR) was defined as resistance to one or more antimicrobial agents in three or more classes of tested drugs (Magiorakos et al., 2012).
β-Lactamase gene detection
Ampicillin-resistant E. coli were tested for the presence of bla TEM, bla SHV, bla OXA, bla CMY (Dallenne et al., 2010), and bla CTX-M (Batchelor et al., 2005; Tarlton et al., 2018) β-lactamase genes by PCR. The amplification products were sequenced at the University of California, Berkeley DNA Sequencing Facility. Sequences were analyzed with Geneious, then submitted to BLASTn for β-lactamase variant identification.
Statistics
Categorical variables were compared by Fisher's exact test (two tailed) with GraphPad Prism v.6.01 (GraphPad Software, San Diego, CA). A p value of <0.05 was considered statistically significant. Genotype diversity was analyzed by Simpson's diversity index (Simpson, 1949).
Results
Distribution of MLST genotypes among human and poultry E. coli isolates
We analyzed 149 human ExPEC isolates and 111 poultry E. coli isolates. The human isolates were classified into 55 distinct STs, whereas the poultry isolates were classified into 30 STs (Fig. 1 and Supplementary Table S1). Of these isolates, 10 ExPEC and 28 poultry E. coli could not be assigned to a known ST. The most common STs identified in human ExPEC were ST69, ST131, ST789, and ST73, accounting for 42 (28%) of the human E. coli isolates. The most common STs identified in poultry E. coli were CC155 (ST58, ST155) and ST117, accounting for 33 (30%) of the E. coli isolates from chickens. The Simpson's diversity index for poultry ST genotypes was 0.91, whereas the diversity index for human ST genotypes was 0.97.
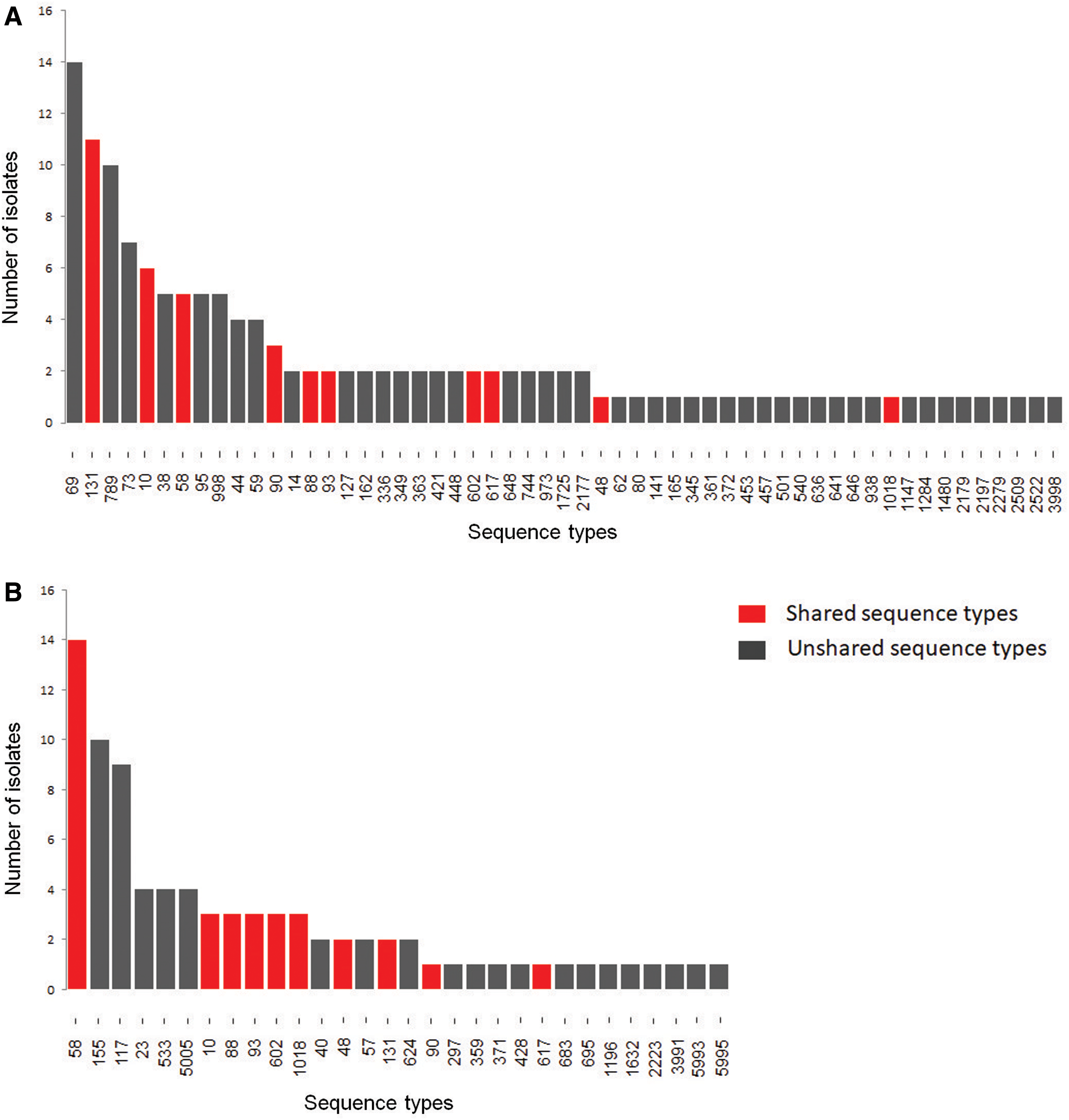
Distribution of multilocus STs of Escherichia coli isolates obtained in Brazil from
Overall, 75 different STs were detected among 222 E. coli isolates. Twenty STs were found exclusively among poultry isolates, while 45 STs were found only among human isolates. Ten STs were shared between human and poultry isolates (ST10, ST48, ST58, ST88, ST90, ST93, ST131, ST602, ST617, and ST1018). Thirty-five (32%) poultry E. coli and 35 (23%) human ExPEC isolates belonged to the 10 shared genotypes (Table 1).
Distribution of Escherichia coli Isolates Among the 10 Shared Genotypes Detected in Human Clinical Samples and Poultry Samples
BSI, bloodstream infection; CC, clonal complex; ST, sequence type; UTI, urinary tract infection.
Phylogenetic relationship between human and poultry E. coli isolates
The E. coli STs were compared phylogenetically based on a minimal spanning tree using eBURST (goeBURST). Figure 2 shows the distribution of STs found in human clinical isolates (community-acquired or hospital-acquired) and in poultry (commercial broiler chickens or backyard chickens). STs from human clinical isolates are distributed across the entire spectrum of the goeBURST diagram. Putative founder STs in both human ExPEC and poultry E. coli belonged to CC10, CC23, CC131, CC155, CC168, CC446, and CC648. Putative founder STs that contained human isolates only belonged to CC86, CC95, and CC141, whereas founder STs that contained avian isolates only belonged to CC350.
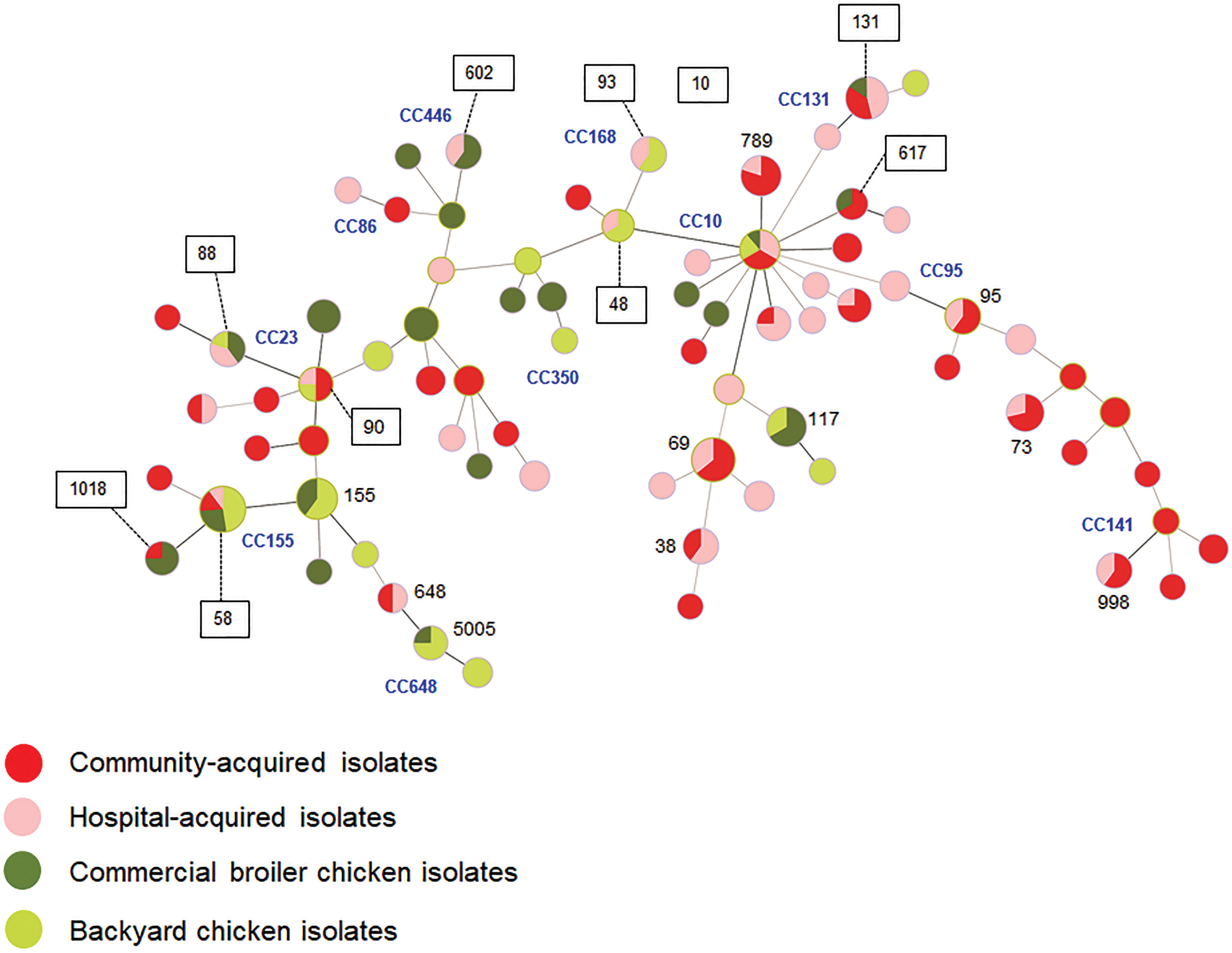
A population snapshot of 222 Escherichia coli isolates, diagrammed based on a minimal spanning tree using optimized eBURST (goeBURST), based on PHYLOViZ software (
Antimicrobial susceptibility of human and poultry E. coli isolates
E. coli isolates were tested for susceptibility to 12 antimicrobial agents (Supplementary Table S2). Fifty-two (35%) of 149 human clinical isolates, 23 (45%) of 51 backyard chicken isolates, and 3 (5%) of 60 commercial broiler chicken isolates were susceptible to all agents tested (pan-susceptible; p < 0.0001 for backyard vs. commercial broiler chicken and human vs. commercial broiler chicken isolates). The most prevalent antimicrobial resistance phenotypes found in both human ExPEC and poultry E. coli were to ampicillin and to trimethoprim-sulfamethoxazole, found in 81 (54%) and 69 (46%) human clinical isolates, and 54 (49%) and 56 (50%) poultry isolates, respectively. Forty-three (29%) of 149 human clinical isolates were MDR—21 (26%) of 81 CA-UTI, 13 (41%) of 32 HA-UTI, and 9 (25%) of 36 HA-BSI. Interestingly, 39 (65%) of 60 commercial broiler chicken isolates were MDR; however, only 4 (8%) of the 51 backyard chicken isolates showed this profile (p < 0.0001). Among human ExPEC isolates, ST131, ST69, and CC10 comprised 16 (37%) of 43 MDR strains; among commercial broiler chicken E. coli, ST117, CC155, and CC23 comprised 15 (39%) of 38 MDR strains. Antimicrobial resistance profiles of the 70 isolates belonging to the 10 shared genotypes are shown in Table 2. Eight (23%) human isolates and seven (20%) poultry isolates demonstrated pan-susceptibility. All poultry isolates belonging to ST48 and ST90 were pan-susceptible, whereas among human clinical isolates none of the shared genotypes were composed entirely of pan-susceptible isolates. Fourteen (40%) human isolates and 10 (29%) poultry isolates were MDR. Two identical antimicrobial resistance profiles were shared by human clinical isolates and poultry isolates within two genotypes—ST58 and ST88, both profiles including resistance to ampicillin and trimethoprim-sulfamethoxazole.
Antimicrobial Resistance Profiles and β-Lactamase (bla) Genes of the 70 Isolates Belonging to the 10 Shared Genotypes in Human Clinical and Poultry Samples
All isolates tested were susceptible to ertapenem and imipenem.
Pan-susceptible isolates were defined as those susceptible to all 12 antimicrobial agents tested.
MDR was defined as resistance to one or more antimicrobial agents in three or more classes of tested drugs.
Isolates with antimicrobial resistance were resistant to at least one of the 12 antimicrobial agents tested.
AMC, amoxicillin-clavulanic acid; AMK, amikacin; AMP, ampicillin; CIP, ciprofloxacin; CRO, ceftriaxone; CXM, cefuroxime; FEP, cefepime; GEN, gentamicin; MDR, multidrug resistance; NIT, nitrofurantoin; Pan-S, pan-susceptible; ST, sequence type; SXT, trimethoprim-sulfamethoxazole.
β-Lactamase gene detection in β-lactam-resistant human and poultry E. coli isolates
Of the β-lactam-resistant isolates, 34 (89%) of 38 HA-UTI/BSI isolates, 37 (86%) of 43 CA-UTI isolates, 34 (85%) of 40 commercial broiler chicken isolates, and 9 (64%) of 14 backyard chicken isolates contained at least one of the β-lactamase genes tested (Table 3). Of the β-lactamase genes examined, bla TEM-1 was the most prevalent, found in 29 (85%) of 34 HA-UTI/BSI isolates, 36 (97%) of 37 CA-UTI isolates, 21 (62%) of 34 commercial broiler chicken isolates, and 8 (89%) of 9 backyard chicken isolates. The distribution of ESBL genes was different in human clinical isolates compared with commercial broiler chicken isolates; 15 (21%) of 71 human clinical isolates compared with none of 34 commercial broiler chicken isolates carried bla CTX-M-15 (p = 0.002), while one (1.4%) of 71 human clinical isolates compared with 13 (38%) of 34 commercial broiler chicken isolates carried bla SHV-2 (p < 0.0001). However, both human clinical isolates and commercial broiler chicken isolates carried bla CTX-M-2 and bla CTX-M-8. None of the backyard chicken isolates contained ESBL genes. No ESBL genes were found among the pandemic lineages ST69, ST73, or ST95. The bla OXA-1 gene (non-ESBL) was detected only in human isolates, and found only in strains that also contained bla CTX-M. Among isolates from the 10 shared genotypes, 1 human ST58 and 7 poultry ST58 were found to contain the exact same β-lactamase gene—bla TEM-1 (Table 2). Across all STs, the bla TEM-1-only profile was found in 44 (62%) of 71 ExPEC and 23 (53%) of 43 poultry E. coli isolates. Two additional β-lactamase gene profiles were shared; one HA ExPEC and three commercial broiler chicken E. coli carried bla CTX-M-8, whereas six ExPEC and one commercial broiler chicken E. coli carried bla CTX-M-8+bla TEM-1.
Distribution of β-Lactamase (bla) Genes in β-Lactam-Resistant Escherichia coli Isolates from Human Clinical Samples and Poultry Samples
The p value compares β-lactamase carriage in hospital-acquired and community-acquired human isolates.
The p value compares β-lactamase carriage in commercial broiler chicken and backyard chicken isolates.
The p value compares β-lactamase carriage in commercial broiler chicken and human isolates.
ESBL genes include SHV-2, CTX-M-2, CTX-M-8, CTX-M-15, and CTX-M-27.
BSI, bloodstream infection; EBSL, extended-spectrum β-lactamase; UTI, urinary tract infection.
Discussion
It has been suggested that poultry are a potential reservoir for E. coli that cause extraintestinal disease in humans, since shared genotypic and phenotypic characteristics have been found among human ExPEC and E. coli isolated from poultry (Vincent et al., 2010; Overdevest et al., 2011; Bergeron et al., 2012; Mora et al., 2013). Here, we analyzed the genotype and antimicrobial susceptibility profile of E. coli isolated from human infections and poultry sources that were collected in Southeast Brazil, to assess the potential and degree to which poultry may serve as a reservoir for ExPEC.
We found that 10 genotypes were shared among human ExPEC and poultry E. coli. Of the 10 shared genotypes, ST131 and ST10, recognized worldwide as pandemic lineages of ExPEC (Adams-Sapper et al., 2013; Riley, 2014), were the most common STs among human UTI and BSI E. coli isolates. This result is consistent with studies from Brazil and the United States, which compared genotypes of E. coli isolated from human UTI and/or BSI and avian sources (Maluta et al., 2014; Yamaji et al., 2018). Other shared genotypes—such as ST48, ST58, ST93, ST602, and CC23—while not considered pandemic lineages, are of interest because members of these STs have previously been detected in poultry and implicated in extraintestinal human diseases worldwide (Oteo et al., 2010; Bergeron et al., 2012; Maluta et al., 2014; Manges et al., 2015; Day et al., 2016; van Hoek et al., 2016; Liu et al., 2017). Some of these isolates shared more genetic characteristics than merely ST lineage membership, such as resistance genes.
ST69, ST73, and ST95—other important ExPEC pandemic lineages—were among the most common STs detected in human clinical E. coli in this study, accounting for 17% of all urine and blood isolates. However, we did not find these STs in poultry, which is in accordance with other reports (Maluta et al., 2014; Manges et al., 2015). The lack of detection of these STs in poultry isolates in this work may be due to the limited number of samples tested, as other studies from Brazil, the United States, and Europe have reported these STs in food animal sources, including poultry (Nandanwar et al., 2014; Day et al., 2016; Cunha et al., 2017; Yamaji et al., 2018). Of the 20 STs that were unique to poultry in this study, 15 (ST23, ST40, ST57, ST117, ST155, ST297, ST359, ST371, ST428, ST533, ST624, ST683, ST695, ST1196, and ST5005) have been reported previously from human ExPEC isolates deposited in the MLST database (
Identical antimicrobial resistance profiles were more commonly shared by different STs across human ExPEC and poultry E. coli. However, among the 10 shared genotypes, ST58 and ST88 isolates shared identical antimicrobial resistance profiles, and ST58 isolates also shared the bla TEM-1 β-lactamase gene. Interestingly, commercial broiler chicken isolates were significantly more likely to be MDR than human clinical or backyard chicken isolates. The fact that the poultry industry uses antimicrobial agents in animal feed to maintain health and productivity, which presumably selects for antimicrobial-resistant bacteria (Van Boeckel et al., 2015), may explain the higher level of antimicrobial resistance in commercial broiler chicken isolates compared with backyard chicken isolates. However, one study from the United States found that human UTI isolates were more likely to be antimicrobial resistant than retail meat isolates, which included poultry (Yamaji et al., 2018). The contrasting results from our study compared with the U.S. study may be due to differences in the source of poultry isolates (cloaca vs. meat) or sanitary management at the farms of origin.
The bla TEM-1 gene was prevalent in β-lactam-resistant isolates from both humans and chickens. In contrast, many of the ESBL/pAmpC gene variants identified in human and poultry isolates were different; the bla CTX-M-15, bla CTX-M-27, and bla CMY-2 genes were found only among human ExPEC. Interestingly, bla SHV-2 was the most common ESBL gene detected in poultry isolates. To our knowledge, this is the first description of the bla SHV-2 gene in E. coli from poultry in Brazil (Egervärn et al., 2014; Ferreira et al., 2014; Casella et al., 2015). While groups from Canada, Europe, and Japan have detected bla SHV-2 in E. coli isolated from avian and human samples, it is not a commonly reported β-lactamase gene in E. coli (Blanc et al., 2006; Asai et al., 2011; Leverstein-van Hall et al., 2011; Peirano et al., 2017).
bla CTX-M-2, bla CTX-M-8, and bla SHV-2 were the ESBL genes in common among human clinical and poultry isolates, and most of the strains containing these genes were from distinct STs and CCs. Taken together, these results suggest that (1) some ESBL genes may be transferred from poultry E. coli to ExPEC, since all the β-lactamase genes detected in poultry isolates were observed among human isolates but not the contrary; while it is also likely that (2) ESBL genes in human ExPEC and poultry E. coli are acquired from different bacterial sources, perhaps due to existence in different intestinal bacterial communities within each host (Sommer et al., 2009), or (3) humans and poultry may acquire ESBL-producing E. coli from different sources due to other exposures, such as vegetables, water sources, or wildlife (Alcalá et al., 2016; Dropa et al., 2016; Müller et al., 2016; Gomi et al., 2017; Batalha de Jesus et al., 2019).
Most of the human ESBL-producing strains belonged to the ST10 CC (excluding ST10 itself). Since ST131 is one of the most common CTX-M-producing lineages (Brisse et al., 2012; Day et al., 2016; Merino et al., 2016), it was surprising that only three (27%) of 11 ST131 isolates in this study carried ESBL genes. Notably, only two ST131 E. coli were isolated from poultry; one was pan-susceptible and the other was resistant to gentamicin. This is in contrast to the 11 ST131 E. coli isolated from humans, which were each resistant to two or more antimicrobial agents. This again suggests that either poultry and human ST131 E. coli acquire resistance genes in the gastrointestinal tract from different types of bacteria (Sommer et al., 2009), or that human and poultry ST131 E. coli have different reservoirs, and represent phylogenetically distinct populations. Our second hypothesis is supported by a study which reported that ST131 strains isolated from human clinical samples and food animals did not show similarity by pulsed-field gel electrophoresis (Johnson et al., 2012).
In conclusion, while human and poultry E. coli isolates showed genetic diversity, some genotypic and phenotypic characteristics overlapped, reinforcing the zoonotic potential of avian E. coli. Since the ExPEC isolates were not necessarily obtained from people residing close to the chicken production sites, we cannot conclude that the ExPECs we found in this study originated from poultry. Additional research is needed to understand the potential direction and pathways of transmission of ExPEC. A better understanding of ExPEC reservoirs could lead to the identification of effective control interventions to disrupt transmission of these bacteria and their resistance genes.
Footnotes
Acknowledgments
We thank Fernando Avila, Roberto Martinez, and Lucia Vitali for providing the isolates used in this study. C.A.B. was supported by a postdoctoral fellowship from Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) during the development of this work.
Disclosure Statement
The authors have no competing interests to declare.
Supplementary Material
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.