
Abstract
Three severe acute gastroenteritis patients were identified within a 5-h period in a sentinel hospital enrolled in the foodborne pathogen surveillance project in Beijing. All patients had high fever (over 38.5°C), diarrhea, abdominal pain, vomiting, and headache. Ten grams of fresh patient stool sample and 25 g of six suspected foods were collected for real-time PCR screening for 10 major pathogens. Bacterial isolation was performed. Pulsed-field gel electrophoresis (PFGE), multilocus sequence typing (MLST), and antibiotic susceptibility tests were conducted for all the isolates. Whole-genome sequences of the three Campylobacter coli isolates were compared using whole-genome MLST. All stool samples were positive for C. coli, as revealed by PCR. Eleven of the C. coli isolates had the same PFGE and ST type. All isolates were resistant to nalidixic acid, ciprofloxacin, streptomycin, and tetracycline, consistent with the findings of the in silico antibiotic resistance gene profiling. Most coding sequences (99%, 1736/1739) were identical among the three sequenced isolates, except for three frameshift-mutated genes caused by the simple sequence repeats (poly-Gs). This was likely a single-source outbreak caused by a group of highly clonal C. coli. This was the first outbreak of severe gastroenteritis caused by C. coli in China.
Introduction
Campylobacter jejuni and Campylobacter coli are the main pathogens that cause sporadic gastroenteritis worldwide (Sheppard and Maiden, 2015; Costa and Iraola, 2019). However, the outbreak caused by Campylobacter in China is rare. Recently, the isolation of Campylobacter in sporadic diarrheal cases significantly increased following the use of a laboratory modified isolation kit based on the filtration method (Wang et al., 2015; Li et al., 2018). Using this method, a point source outbreak caused by C. coli was first identified in Beijing. This was also the first identified outbreak of gastroenteritis caused by C. coli in China.
Materials and Methods
Ethics statement
Written consent for stool sample collection was obtained from the patients in this study and the data were analyzed anonymously. Ethics approval for this study was also obtained from the Ethics Committee of the Shunyi District CDC, and the academic committee of the National Institute for Communicable Disease Control and Prevention in China CDC.
Patients
In the afternoon of June 6, 2018, three patients with acute gastroenteritis visited Beijing Shunyi district hospital, which is just one sentinel hospital enrolled in the foodborne pathogen surveillance project. There was a 5-h interval between the onset times of the first and last patients. These three patients showed similar clinical symptoms, including high fever (over 38.5°C), diarrhea, abdominal pain, vomiting, and headache. All of them had watery stool and diarrhea 2–10 times per day. The white blood cell count, neutrophil percentage (NEUT%), and C-reactive protein values in the routine blood tests of all patients were significantly increased. According to the epidemic investigation, these three patients worked as taxi drivers and they ate the same food at the same restaurant for lunch on 3 consecutive days before the onset time. According to their statements, the suspected foods were Chinese hamburger (steamed bun with roasted pork) and sheep giblets soup.
Sample collection
A total of 10 g of fresh stool sample was collected from each of the patients (identified as s004, s005, and s006) and 25 g of the suspected food and food components in the restaurant, including steamed buns, cooked pork, cooked rectum meat, cooked sheep entrails, cooked sheep lungs, and other cooked meat (six food samples in total) were collected from the freezer in the restaurant. All of the samples were collected on the second day after the onset of the patients' illness. The samples were transported to the central laboratory of the Shunyi District CDC for examination.
Enteric pathogen screening and bacteria isolation
TaqMan real-Time PCR was performed for the 10 major enteric pathogens, Vibrio cholerae, Vibrio parahaemolyticus, C. jejuni, C. coli, Salmonella, Shigella, diarrheagenic Escherichia coli, norovirus, rotavirus, and enteric adenovirus, using a commercial kit (Qingdao Sinova Biotechnology Co., Ltd., Qingdao, China for Campylobacter; Beijing Applied Biological Technologies Co., Ltd., Beijing, China for the other pathogens). DNA templates from the stool samples and the food enrichment samples were extracted using commercial kits (QIAamp Fast DNA Stool Mini Kit for bacteria, Qiagen GmbH, Hilden, Germany; High Pure Viral Nucleic Acid Kit for viruses, Roche Diagnostics, Mannheim, Germany) using 200 μg of samples from each patient. The positive and negative results refer to the cycle threshold (Ct) value of the real-time PCR that, according to the manual, is Ct<35 with the typical S curve determined as positive; Ct>40 or no typical S curve are determined as negative; and 35 ≤ Ct ≤40 should be repeated 3 times and determined as weak positive after the third repeat.
According to the results of the PCR screening, Campylobacter isolation from the stool samples was carried out using the isolation kit (ZC-CAMPY-001; Qingdao Sinova Biotechnology Co., Ltd., Qingdao, China) described previously(Li et al., 2018). Isolation from the suspected contaminated food was carried out according to the manufacturer's instructions in the kit (ZC-CAMPY-002; Qingdao Sinova Biotechnology Co., Ltd.). Briefly, 25 g of the remaining suspected contaminated food was added into 225 mL of buffered peptone water (BPW) and alkaline peptone water (APW). Then, 2 mL BPW mixed with suspected contaminated food was added into 4 mL of enrichment buffer provided in the kit. The enrichment buffer was incubated at 42°C for 24 h in a microaerophilic atmosphere consisting of 5% O2, 10% CO2, and 85% N2. BPW and APW were incubated at 37°C for 24 h. The suspected monoclonal colonies on the culture medium were picked for colony identification with PCR (Wang et al., 2002). The real-time PCR assay for Campylobacter was also performed on the enrichment buffer of the suspected contaminated food.
Pulsed-field gel electrophoresis and multilocus sequence typing for the Campylobacter isolates
Pulsed-field gel electrophoresis (PFGE) was performed as described previously (Zhou et al., 2011). PFGE images were obtained using a Universal Hood II (Bio-RAD) and analyzed using BioNumerics software version 7.1 (Applied Maths, Sint-Martens-Latem, Belgium). Multilocus sequence typing (MLST) was performed by amplifying and sequencing seven housekeeping genes loci, aspA, glnA, gltA, glyA, pgm, tkt, and uncA, and by using previously described primers for C. jejuni and C. coli. The nucleotide sequences of the amplicons were determined using the published oligonucleotide primers and reaction conditions. Allele numbers and STs were assigned using the Campylobacter MLST database website (
Antimicrobial susceptibility testing for Campylobacter isolates
The minimum inhibitory concentrations (MICs) for 11 antimicrobials (erythromycin, azithromycin, nalidixic acid, ciprofloxacin, gentamicin, streptomycin, chloramphenicol, florfenicol, tetracycline, telithromycin, and clindamycin) were determined for all Campylobacter isolates using an agar dilution method as recommended by CLSI (M100-S25, 2015). Briefly, Mueller–Hinton agar (CM0337, Oxoid, United Kingdom) plates supplemented with 5% de-fibrinated sheep blood and double-diluted antimicrobial agents at concentrations ranging from 0.02 to 256 μg mL−1 were used. The turbidity of the bacterial suspension was 0.5 McFarland (∼108 CFU mL−1). Plates were inoculated with a multipoint inoculation instrument with 1 mm diameter inoculating pins and incubated at 37°C for 48 h under microaerobic conditions. Inoculated plates started with the lowest concentration. The growth control plates were inoculated before and after each inoculation for different agents. The MIC was read as the lowest concentration without visible growth.
The breakpoints for resistance used in this study were those with a MIC value greater than or equal to the standard values used in the National Antimicrobial Resistance Monitoring System (NARMS-2014) for Campylobacter in the USA: erythromycin (≥32 μg mL−1), azithromycin (≥8 μg mL−1), nalidixic acid (≥64 μg mL−1), ciprofloxacin (≥4 μg mL−1), gentamicin (≥8 μg mL−1), streptomycin (≥16 μg mL−1), chloramphenicol (≥32 μg mL−1), florfenicol (≥8 μg mL−1), tetracycline (≥16 μg/mL−1), telithromycin (≥16 μg mL−1), and clindamycin (≥8 μg mL−1). C. jejuni ATCC 33560, Staphylococcus aureus ATCC29213, and E. coli ATCC25922 were used as controls.
Genome analysis for the three C. coli isolates
The draft genomes of three C. coli isolates from the three patients (identified as cc04, cc05, and cc06, respectively) were sequenced by an Illumina HiSeq 2500 platform at the Beijing Genomics Institute (BGI). The DNA for each isolate was purified by the QIAamp DNA Mini Kit (#51304; Qiagen GmbH, Hilden, Germany). To sequence the genomes, a 270 bp paired-end library was constructed and then 150 bp reads were generated for each isolate. The sequencing reads were trimmed with Trimmomatic (version 0.38) to remove sequencing adapters(Bolger et al., 2014). The processed reads were assembled de novo by SKESA with the parameter “vector_percent” set to zero (Souvorov et al., 2018). The assembly sequences were annotated with Prokka (version 1.13.3) (Seemann, 2014).
Whole-genome MLST (wgMLST), in silico MLST and wgSNP
The ad hoc wgMLST analysis of these three genome sequences was performed with fast-GeP using the annotated cc004 genome draft as reference (Zhang et al., 2018a). The 7-locus MLST sequence type (ST) of the three draft genome sequences was determined by using the online tool Genome Comparator on PubMLST website (
Antibiotic resistance genes
The antimicrobial resistance genes on the whole-genome sequences were screened by searching the latest NCBI Bacterial Antimicrobial Resistance Reference Gene Database (BioProject Accession: PRJNA313047) (accessed: 21.02.2019) with the pipeline ARRicate (version 0.8.10) (
Results
Pathogen screening with real-time PCR
Except for C. jejuni and C. coli, the three stool samples were negative for the other five bacterial pathogens, V. cholerae, V. parahaemolyticus, Salmonella, Shigella, diarrheagenic E. coli, and three major enteric viruses, norovirus, rotavirus, and enteric adenovirus. All of the stool samples from the three patients (s004, s005, and s006) were positive for C. coli (with Ct values of 21.19, 38, and 20.88, respectively) and two patients (s004 and s005) were also positive for C. jejuni (with Ct values of 25.86 and 27.21, respectively). Both the stool sample and the culture enrichment of s006 had negative PCR results for C. jejuni (no Ct value was obtained).
Although the DNA samples from the tested food samples, which the patients had eaten in the restaurant, were negative for all 10 tested pathogens the DNA samples from the culture enrichment of the cooked sheep entrails and cooked sheep lungs were weak positive for C. coli (with Ct values of 40).
Molecular subtyping for the Campylobacter isolates
In total, 11 C. coli isolates were obtained from the three patient stool samples (five C. coli isolates were picked from s004, one C. coli isolate was picked from s005, and five C. coli isolates were picked from s006). In total, 10 C. jejuni isolates were obtained from only two patients' stool samples (six isolates from s004 and four isolates from s005). No isolates were identified from any tested food samples.
All 11 C. coli isolates from the three patients showed an identical PFGE profile (Fig. 1). Ten C. jejuni isolates showed two types of PFGE pattern. Six C. jejuni isolates from sample s004 had an indistinguishable PFGE pattern and four C. jejuni isolates from sample s005 had an indistinguishable PFGE pattern (Fig. 2). Meanwhile, all 11 C. coli isolates had the same ST, ST9227, a novel ST type. Six C. jejuni isolates from sample s004 had the identical ST type (ST9254) and four C. jejuni isolates from sample s005 had the identical ST type (ST883).

PFGE of 11 Campylobacter coli isolates from three patients in this outbreak. Lanes 1, 5, 10, and 15: reference standard H9812; lanes 2 through 4 and 6 through 7: 5 C. coli isolates from s004; lanes 8: 1 C. coli isolate from s005; lanes 9 and 11 through 14: 5 C. coli isolates from s006; All of 11 C. coli isolates had the same pattern. Pfge, pulsed-field gel electrophoresis.
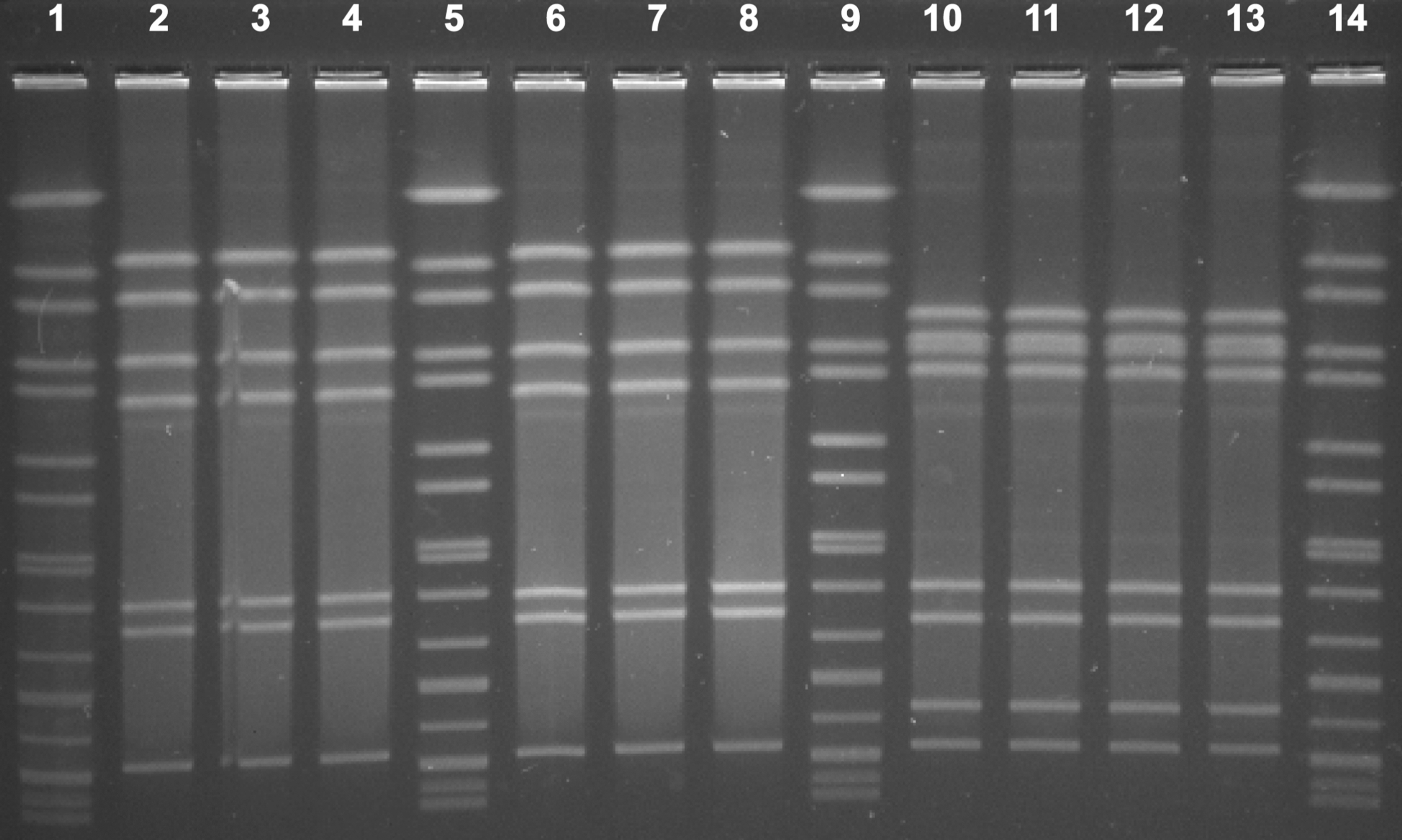
PFGE of 10 Campylobacter jejuni isolates from s004 and s005 in this outbreak. Lanes 1, 5, 9, and 14: reference standard H9812; lanes 2 through 4 and 6 through 8: 6 C. jejuni isolates from s004; lanes 10 through 13: 4 C. jejuni isolates from s005; 6 C. jejuni isolates from s004 had the same pattern; 4 C. jejuni isolates from s005 had the same pattern; C. jejuni isolates from different stool samples had different patterns.
Antibiotic susceptibility
Antibiotic susceptibility tests were performed for a total of 21 isolates (11 C. coli and 10 C. jejuni). All 11 C. coli isolates had the same susceptibility pattern: they were all resistant to ciprofloxacin, nalidixic acid, streptomycin, and tetracycline. Six C. jejuni isolates from s004 had the same profile and all were resistant to ciprofloxacin, nalidixic acid, and tetracycline. However, the other four C. jejuni isolates from s005 were resistant to tetracycline only. The MIC and ST of each isolate are presented in Table 1.
Antimicrobial Susceptibility and Multilocus Sequence Type of the Tested Isolates
ERY, erythromycin; AZI, azithromycin; NAL, nalidixic acid; CIP, ciprofloxacin; GEN, gentamicin; STR, streptomycin; CHL, chloramphenicol; FLO, florfenicol; TET, tetracycline; TEL, telithromycin; CLI, clindamycin.
Genomic characteristics of three C. coli isolates
The genome coverage of the sequencing results for the three isolates were all 300 X. The full length of the genome of the three isolates (cc004, cc005, and cc006) are 1691710, 1692096, and 1691990, respectively. The mean length of the scaffold of the three isolates (cc004, cc005, and cc006) are 89037.36842, 94005.33333, and 84599.5, respectively. The MLST analysis with the whole-genome sequences confirmed that all three isolates belonged to ST-9227 of the ST-828 clonal complex. The ad hoc wgMLST analysis found that among the 1743 reference coding sequences (CDSs), 1739 CDSs were complete and shared by the three genome sequences. Most of the CDSs were identical (n = 1736) and three of them were polymorphic. Four CDSs were excluded from the analysis due to truncation caused by assembly gaps.
Mapping the polymorphic loci to the published C. jejuni strain YH502 complete genome (GenBank Acc. NZ_CP018900) revealed that the genetic differences were caused by simple sequence repeats (SSRs) in all the three cases (Fig. 3). The variance in the length of the homopolymeric tracts (Gs) caused frame shift mutations in the three loci. Consequently, frameshift mutations were found at the gene BU815_RS06565 (homology protein Fmt, coding methionyl-tRNA formyltransferase, Cj0098 in C. jejuni NCTC11168) for isolates cc005 and cc006; BU815_RS00280 (homology iron-binding protein, cj0045c in C. jejuni NCTC11168) for isolates cc004 and cc005; and the gene BU815_RS6490 (homology hypothetical protein, cj1295 in C. jejuni NCTC11168) for isolates cc004 and cc006. No SNP was found in the wgSNP analysis.

The assembled sequences of the C. coli isolate cc004 were ordered by length. The outer ring indicates the CDSs in the forward strand, and the inner ring indicates the CDS in the reverse strand. Three polymorphic loci were marked in the corresponding contigs and were marked.
Three antimicrobial resistance genes were found in the whole-genome sequences. These three genes were identical in all three isolates and highly similar (>99% identity and alignment coverage) to tetracycline resistance ribosomal protection protein gene (GenBank Acc. NG_048259; tetracycline resistance), OXA-61 family class D beta-lactamase gene (GenBank Acc. NG_057542; multi-resistance to β-lactam antibiotics), and aminoglycoside 6-adenylyltransferase gene (GenBank Acc. NG_054665; streptomycin resistance) respectively. All of the isolates had a Thr-86 to Ile (ACTCATT) gyrA mutation.
Discussion
Once the membrane filter method became widely used to isolate Campylobacter from diarrheic patients in China's pathogen surveillance project, Campylobacter infection was identified as the major cause of diarrhea in China (Li et al., 2018). Although most Campylobacter infections are caused by C. jejuni, C. coli infection has increased significantly (from about 5% to 15%) after 2000 (unpublished data). However, the outbreak had never been identified before. In addition, the most serious concern is that the antibiotic resistance in C. coli has significantly increased in China. According to our previous study, almost 100% of C. coli were resistant to quinolones, fluoroquinolones, and tetracycline, and more than 80% of C. coli isolates in China were resistant to erythromycin and showed multidrug-resistant characteristics (Maojun et al., 2014). Infection caused by multidrug-resistant C. coli is a concern for China and the rest of the world.
In this study, PFGE, MLST, and the WGS analysis indicated the C. coli isolated from the three clinical cases were the same clonally; ST9227 was a novel ST type identified in the MLST database that belonged to the ST828 clonal complex, the major C. coli clonal complex in China (Maojun et al., 2014; Zhang et al., 2018b). This was the first report of severe gastroenteritis outbreak caused by C. coli in China. The virulence characteristics of this strain need further investigation.
Antibiotic tests proved this C. coli strain was also multidrug-resistant. It was resistant to ciprofloxacin, nalidixic acid, streptomycin, and tetracycline; this phenotype is consistent with the results of the genomic antimicrobial resistance gene analysis and also showed why the first intravenous levofloxacin treatment did not significantly decrease the symptoms of gastroenteritis.
In this study, the pre-qPCR screening results were consistent with those of the bacterial culture of the patient stool samples. Unfortunately, no isolates were obtained from a food sample and no additional people could be recalled who ate the same food in the restaurant in the same time. Although no isolates were identified from any of the tested food samples, the cooked sheep entrails and lungs, which exhibited weak positive PCR results, were ingredients of the sheep giblets soup, which the patients had eaten for lunch before the onset of diarrheic symptoms. Thus, these ingredients were the likely source of contamination that caused the infection. According to the statement from the restaurant, the sheep giblets soup usually was made by adding the boiling soup on top of the cooked sheep entrails and lungs and the customer can eat it directly after several minutes. The boiling temperature or the boiling time for the soup might be not enough to inactivate the pathogen. Poultry and sheep were the main source species for C. coli, and sheep had been identified as a predominant source of human infection with C. coli in other countries (Sheppard et al., 2009; Roux et al., 2013; Nohra et al., 2016). So far, there have been few reports of Campylobacter from sheep in China, the results of this study will lead us to conduct further investigation.
The results of PFGE, MLST, and the antibiotic susceptibility analysis for all isolates indicated that two different C. jejuni strains (ST9254, ST883) were coinfected with C. coli (ST9227) in two patients (case 004 and case 005). C. jejuni ST9254 was the first ST to be identified in this region. These two C. jejuni strains might not be the main reason for the severe outcome of this event because patient s006 had only C. coli isolates and PCR positive results, but patient s006's clinical outcome was as serious as the other two cases. It is possible that C. jejuni caused a sporadic infection in these two patients. Multiple clonal coinfections have also been identified in diarrheal patients in China and in other countries (Lucarelli et al., 2016).
The whole-genome sequence analysis found very few genetic differences between the three isolates. This indicates that the outbreak was likely a single-source outbreak that was caused by a group of highly clonal C. coli. Although the allele differences were detected in three loci by ad hoc wgMLST analysis, the differences were not caused by DNA substitution mutation, but by variation of SSRs. SSR is an important bet-hedging mechanism in bacteria that increases the chance of survival in the fluctuating environments (Davidson and Surette, 2008; Veening et al., 2008). Variations in SSR loci are generated by a slipped strand mis-pairing mechanism and have a much higher mutation rate than DNA substitution mutation (Moxon et al., 2006); thus, they are a poor indicator of bacterial relationships (Revez et al., 2014; Llarena et al., 2017).
In this study, two of three loci (BU815_RS00280 and BU815_RS6490, homology cj0045c and cj1295, respectively) were previously recognized as core phase variable genes that had SSRs both in C. coli and in C. jejuni (Hitchen et al., 2010; Aidley et al., 2018). These genes might be the specific adaptation in a different host during this same clonal infection.
Conclusions
Although no isolated pathogen was obtained from the food, the results from the laboratory study and the epidemical investigation indicated this infection was likely a single-source outbreak caused by a group of highly clonal C. coli. An apparent shift of the dominant species from C. jejuni to the multidrug-resistant C. coli has been found in chicken previously (Wang et al., 2016). The campylobacteriosis caused by C. coli was also found increased in the foodborne pathogen surveillance project here (unpublished). This was the first identified outbreak of gastroenteritis caused by C. coli in China. The emergence of antibiotic resistance in C. coli should be a grave concern both in China and globally. Our findings also indicate that there is a need for a new wgMLST algorithm for more accurate genotyping that could identify allele differences that result from SSRs.
Footnotes
Disclosure Statement
No competing financial interests exist.
Acknowledgment
We thank the native English speaking scientists of Cactus Communications (Shanghai China) for editing our article.
Funding Information
This work was supported by the National Key Program of China (2018ZX10305409 and 2018ZX10712-001) and the Major State Basic Research Development Program (2013CB127204). NF and JZ were funded by the New Zealand Food Safety Science & Research Centre.