
Abstract
We studied the efficacy of cold atmospheric-pressure plasma (CAP), generated by a two-dimensional array of integrated, coaxial, microhollow, dielectric barrier discharge plasma, against Salmonella enterica serovar Heidelberg (SH) on stainless steel, romaine lettuce, and chicken breast. Exposure of SH to CAP on a dry stainless steel surface had low bactericidal efficacy; only 2.5 log10 colony-forming units (CFUs) were inactivated after 10 min of exposure. On the other hand, the presence of moisture led to decontamination of ∼6.5 log10 CFUs after only 3 min. Although complete decontamination was not achieved on lettuce and chicken breast samples after 10 min of exposure, SH counts were reduced by ∼4.5 and 3.7 log10 CFUs, respectively. A partial suppression of bactericidal effects was observed on steel surfaces when it was coated with bovine serum albumin before spiking with bacteria and exposure to plasma, indicating that the proteinaceous nature of chicken meat may be partially responsible for lower efficacy of CAP on chicken muscles. The initial bacterial load was also found to affect the anti-SH efficacy; at high (∼6.5 log CFUs) and low (∼3.5 CFUs) initial counts, the time required for complete decontamination on stainless steel and lettuce decreased from 3 to 0.5 min and >10 to 1 min, respectively. However, the analysis of inactivation kinetics showed that effects of initial loads of contamination on the rate of bacterial inactivation were not statistically significant. This is consistent with other findings for conditions where both bacterial loads were under the multilayering threshold that might have affected the rate of killing.
Introduction
A large variety of foods is implicated in foodborne salmonellosis such as poultry meat (Zhao et al., 2008), eggs, and fresh produce (Gomez et al., 1997). Among the 2500 serotypes of Salmonella, Salmonella enterica serovar Heidelberg (SH) is one of the most frequently isolated serovars from clinical cases in humans, from retail meats and chicken, and chickens at slaughter (Zhao et al., 2008; CDC, 2019). Antimicrobial drug resistance in this serovar contributes to severe extraintestinal infections such as septicemia and myocarditis (Crump et al., 2011).
Strategies for reducing bacterial pathogen load in poultry farms, slaughterhouses, and food-processing plants depend upon maintaining sanitary conditions using regular schedules of cleaning and disinfection (Vandeplas et al., 2010). Chlorine rinses are often used during poultry processing (Zhao and Doyle, 2006), but their use is coming under increased scrutiny because of toxicity associated with byproducts of disinfection (Rowan et al., 2007). Therefore, there is an urgent need to develop alternative approaches and technologies for the destruction of microbial pathogens on contaminated surfaces and foods.
Cold atmospheric pressure gaseous plasma (CAP) is a promising, alternative nonthermal technology for decontamination of food and food-processing surfaces without affecting the quality of treated foods (Misra et al., 2014; Ulbin-Figlewicz et al., 2015; Wang et al., 2016). CAP refers to gas, which is partially ionized by an electric field and comprises photons, ions, free electrons, and reactive species that have germicidal activities. These include atoms (e.g., O and N), molecules (e.g., O3, H2O2, and HNO2), and radicals {•OH, NO•, and singlet oxygen [O2(a1Δg)]} (Graves, 2012). Several studies have described the bactericidal effect of CAP against a wide spectrum of foodborne bacteria, including different serovars of S. enterica such as Typhi, Typhimurium, Stanley, and Enteritidis (Niemira and Sites, 2008; Dirks et al., 2012; Matan et al., 2014; Choi et al., 2016; Min et al., 2016; Oh et al., 2017). However, no study is available on the effect of CAP on SH, and the work on multidrug-resistant (MDR) strains of Salmonella is scarce (Dirks et al., 2012). Since the bactericidal efficacy of CAP often depends on the type of bacterial strain (Min et al., 2016), we conducted this study to determine the effect of CAP on an MDR strain of SH on chicken breast, romaine lettuce, and stainless steel coupons as models for retail poultry, leafy vegetables, and food-processing surfaces, respectively.
Materials and Methods
Plasma generation system
A detailed description of the microhollow, dielectric barrier discharge plasma (two-dimensional array of integrated, coaxial, microhollow, dielectric barrier discharge [2D-AICM-DBD]) setup (Fig. 1) has been reported (Nayak et al., 2017, 2018; Aboubakr et al., 2020).

2D-AICM-DBD setup used for treatments reported in this work. A key part of this setup is a DBD electrode (Kyocera, Inc.) that has 105 holes (600 μm diameter) arranged in a grid-like array distributed in a 25 × 25 mm2 area. The plasma is produced within these holes. Filtered dry air (feed gas) is blown through the holes at a flow rate of 16.4 slm. In this study, samples spiked with SH were placed on a Teflon® block platform (30 × 30 mm2), followed by treatment with the plasma source at a distance of 10 cm. The enclosure was 12 cm in length. The samples were not in direct contact with the plasma and were exposed to the plasma effluent, which contains ROS and RNS (Nayak et al., 2018). This study was conducted at a plasma power of 14.5 W for all results reported in this work, a power previously shown to have a significant germicidal effect (Nayak et al., 2018). 2D-AICM-DBD, two-dimensional array of integrated, coaxial, microhollow, dielectric barrier discharge; RNS, reactive nitrogen species; ROS, reactive oxygen species; SH, Salmonella enterica serovar Heidelberg; slm, standard liters per minute. Color images are available online.
Bacterial strain
An MDR strain of SH (strain NS-029) was used (Nisar et al., 2017). A stationary-phase culture [∼109 CFUs/mL] was prepared by subculturing the revived culture in trypticase soy broth (TSB; Becton Dickinson) overnight. The working culture of SH was prepared by adjusting the count of bacterial cells to 3.5 × 108 CFUs/mL using sterile distilled water. The stationary-phase culture was used because log-phase cultures are more susceptible to inactivation by CAP and other treatments (Scher et al., 2005; Deng et al., 2007).
Qualitative bactericidal effect of CAP against SH on agar medium
One-milliliter aliquots of stationary-phase cultures of SH (∼109 CFUs) were spread over the surface (78.5 cm2) of tryptic soy agar (TSA) plates (Becton Dickinson), and the cell density on agar surface was ∼1.3 × 107 CFUs/cm2. The plates were kept open inside a biosafety cabinet for 2 h for drying the agar surface. Duplicate plates were exposed to CAP for 0 (control), 1, 5, and 10 min at 16.4-standard liters per minute (slm) airflow rate, 14.5-W plasma generation power, and 12.5-cm exposure distance (length of enclosure 12 cm plus 0.5-cm distance between the edge of the enclosure and the surface of TSA. Because the enclosure's diameter (45 mm) in the 2D-AICM-DBD setup is smaller than that of the Petri dish (∼100 mm), we adjusted the location of the enclosure on the center of the TSA plate.
Preparation of food samples and food-contact surface coupons
Chicken breast obtained from a local market was cut aseptically into squares (∼1 cm2 area and ∼0.5 thickness). Squares of romaine lettuce (∼1 cm2) were cut from interior noncurly leaves. To remove native microbial loads, samples of chicken and lettuce were immersed in a sodium hypochlorite solution (300 ppm) for 5 min (Min et al., 2016), followed by rinsing three times with sterile distilled water. These pieces were dried for 30–45 min inside a biosafety cabinet. To ensure samples were salmonella free, we took prespiking swabs from each sample and inoculated them in tetrathionate broth (TTB) for enrichment. After incubation at 37°C overnight, a loopful from each TTB tube was streaked on brilliant green agar (BGA) plates, followed by incubation at 37°C for 24 h. As a model for food-processing surfaces, disc-shaped coupons of stainless steel (area ∼1 cm2) were used. Before use, the coupons were washed with soap and water, followed by rinsing in distilled water and autoclaving. For spiking of all three types of samples, 20-μL aliquots of working culture (∼7 × 106 CFUs) were spread aseptically on several pieces of each sample, followed by storage inside a biosafety cabinet for 30–45 min to dry.
Recovery of surviving bacterial cells
Immediately after CAP exposure, samples were transferred into 10-mL sterile plastic tubes containing 0.5, 1.0, and 2.0 mL of elution buffer (3% beef extract in 0.05 M glycine, pH 7.2; Goyal and Aboubakr, 2016). for stainless steel, lettuce, and chicken samples, respectively. All tubes were vortexed for at least 2 min and then the eluates were used to determine the number of surviving bacteria using 10-fold serial dilutions.
Wet versus dry exposure
This experiment was conducted on stainless steel coupons only. Bacteria applied to stainless steel discs were exposed to CAP under dry and wet conditions. In wet exposure, surfaces of the SH-spiked steel coupons were sprayed with sterile distilled water (∼30 ± 5 μL) intermittently every 2 min for maintaining a thin layer of moisture on the sample during exposure. In dry exposure, no water was sprayed on the sample either before or during the exposure. Subsequently, all samples spiked with SH were exposed to CAP using only the wet exposure approach for 1, 2, 3, 4, 5, 7, and 10 min of exposure time at the same operational conditions as mentioned above.
Effect of initial count of SH on CAP efficacy
To determine if there is any difference in CAP efficiency between low and high initial bacterial counts, all three sample types were spiked with 20 μL of high-count (∼3.5 × 108 CFUs/mL) or low-count (∼3.5 × 105 CFUs/mL) SH cultures (i.e., ∼7 × 106 CFUs and ∼7 × 103 CFUs per sample for high and low initial loads, respectively). Both pairs of samples (spiked with high and low counts of SH) were exposed to CAP at the same operational conditions.
Influence of protein on CAP's bactericidal effect
Three groups of stainless steel coupons were coated separately with 20-μL aliquots of 1%, 5%, and 10% bovine serum albumin (BSA; Sigma-Aldrich) solutions. After drying, the surfaces of steel coupons were spiked with SH, as described above, followed by exposure to CAP. The bactericidal efficacy of CAP against SH on BSA-coated coupons versus uncoated coupons was compared.
Microbial analysis
Enumeration of surviving bacterial cells
To determine the SH cell count, serial 10-fold dilutions of each eluate prepared in phosphate-buffered saline were plated on duplicate plates of TSA. After incubation at 37°C for 24–48 h, the number of SH cells (expressed as log10 CFUs per sample) was calculated by averaging the number of colonies on two countable plates multiplied by the dilution factor. The reduction in SH cells due to CAP exposure was measured by the difference of CFUs between CAP-treated and control samples.
Detection of viable, but nonculturable, SH
This test was performed on samples that showed zero growth on direct plating to ensure that plasma had eliminated all SH cells, even those that might be in a viable, but nonculturable (VBNC), state. For this purpose, 1 mL of the sample eluate was inoculated in a tube of TTB for enrichment, followed by incubation at 37°C overnight. Then, a loopful from TTB was streaked on BGA plates, followed by incubation at 37°C for 24–48 h.
Statistical analysis and bacterial inactivation kinetics
The results are expressed as means and standard deviations of triplicate experiments. Statistically significant differences between CAP-exposed and control samples were evaluated by one-way analysis of variance performed with Fisher's least significant difference (LSD) test using STATISTICA software, v. 10 (StatSoft, Inc., Tulsa, OK).
The log-linear regression model [Eq. (1)] was used to analyze the bacterial inactivation kinetics after CAP treatment:
where Nt is the count of surviving bacterial cells after CAP treatment (expressed as log10 CFUs/sample), N 0 is the initial bacterial count, D is the decimal reduction time, and t is the plasma exposure time (min).
The D-value, as determined from Equation (1), is defined as the time (min) needed to achieve a 1-log reduction in the bacterial count in a given plasma treatment.
Results and Discussion
Qualitative bactericidal effect of CAP against SH on agar medium
Remote exposure to plasma effluent generated by the 2D-AICM-DBD setup showed inactivation of ∼109 SH CFUs on the surface of TSA plates (Fig. 2). This lethal effect increased with an increase in exposure time. At 10 min of exposure, a complete decontamination of the TSA surface (as indicated by clear agar without the presence of any bacterial colony) was observed on the area located directly underneath the enclosure. A few SH cells survived and formed colonies in the area surrounding the direct treatment zone after incubation, but their number was extremely low (1.2 × 101 CFUs) compared with the thick and continuous lawn of bacterial growth seen in the same area in the control TSA plate. The decontamination effect beyond the area immediately underneath the enclosure is probably due to radial convective transport of reactive oxygen species (ROS) and reactive nitrogen species (RNS) by the continuous airflow along the agar plate.
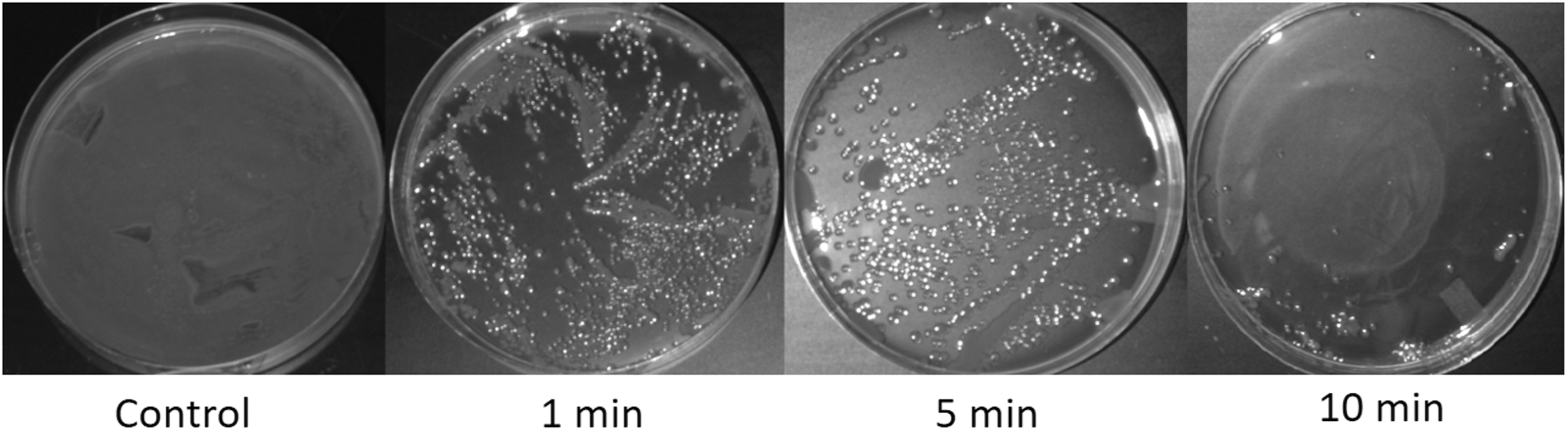
Time-dependent decontamination of the agar surface by exposure to the effluent of 2D-AICM-DBD at a 12.5-cm distance, 16.4-slm airflow rate, and 14.5-W discharge power. Diameter of the enclosure tube = 45 mm. Diameter of the agar plate = 100 mm.
Wet versus dry exposure
We previously found that a moist surface was necessary for virucidal efficacy of this CAP setup (Nayak et al., 2018) and hence we studied if moistening the surfaces before CAP treatment is essential for the bactericidal effect too. The efficacy of CAP on MDR-SH on dry and wet stainless steel surfaces is shown in Figure 3. Under dry conditions, only 2.5 log (D-value = 2.35 min) reduction was observed with no significant differences (p > 0.05) among 3, 5, 7.5, and 10 min of exposure. On the other hand, SH was completely inactivated (∼6.5 log10 CFUs/sample) after 3 min (D-value = 0.49 min) of plasma exposure on moist stainless steel surfaces.
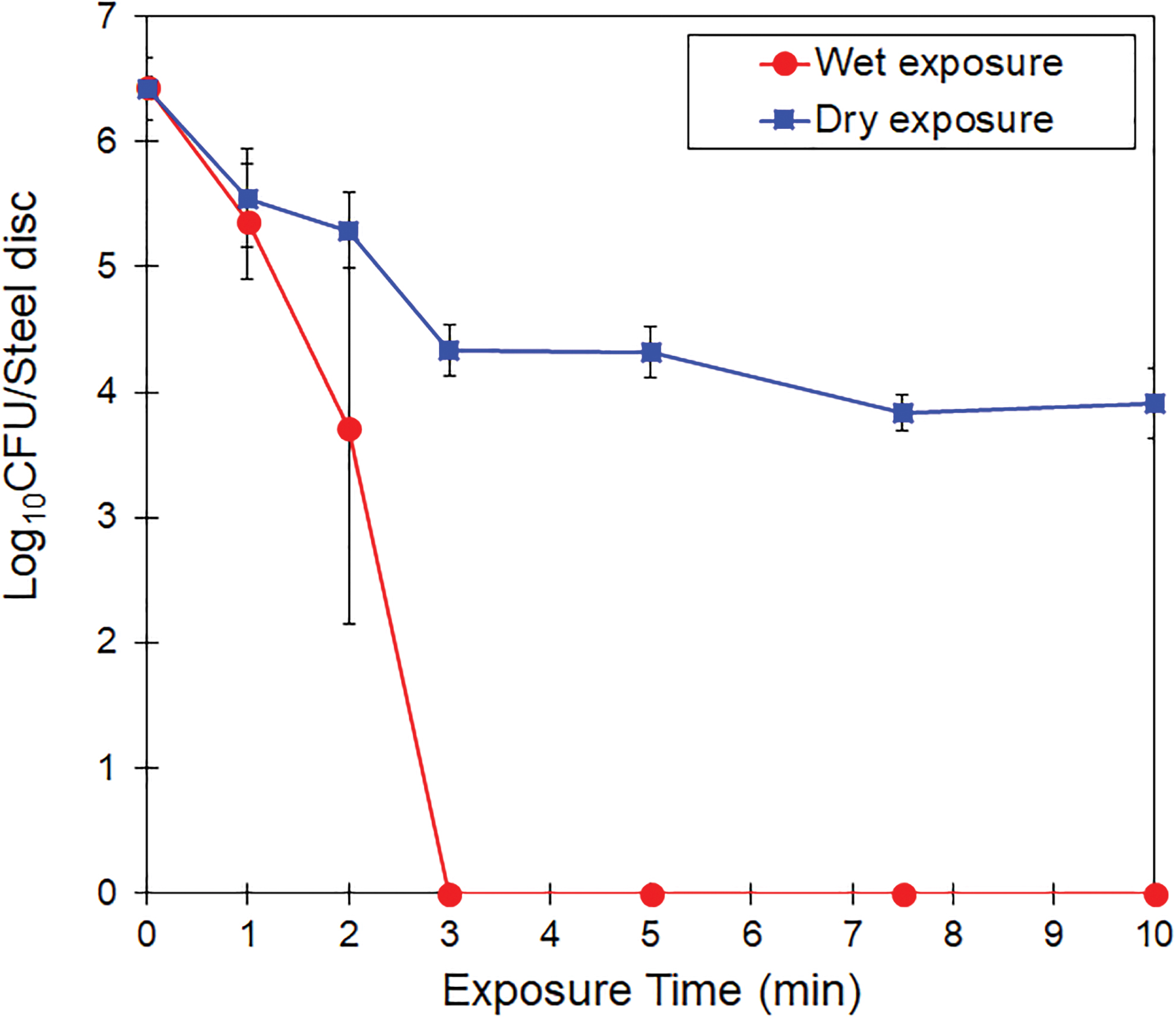
Survival curves of SH after treatment with plasma on dry and moist stainless steel surfaces. The presented data are averages of triplicate experiments. The error bars represent standard deviations. Color images are available online.
These results are consistent with those of Menetrez et al. (2009), who observed that an increase in humidity increased the bactericidal and fungicidal activity of ozone. Ozone is one of the most abundant reactive species produced by CAP source used in this study (Nayak et al., 2018). However, it was shown that RNS played a dominant effect on virus inactivation (Nayak et al., 2018). Similarly, exposure to helium plasma on wet laminates showed greater bactericidal activity than under dry conditions (Neuber, 2016). This suggests that moisture on the treated surface is important to increase the effectiveness of the CAP-produced reactive species. Based on these results, we decided to use humidified substrates in all subsequent experiments.
Bactericidal effect of CAP against SH on chicken, lettuce, and stainless steel surfaces
Complete killing (>6.5 log10 CFUs/sample) of SH with undetectable VBNC cells was achieved after 3 min of exposure on a moist stainless steel surface, but not on lettuce and chicken breast even after 10 min (Fig. 4). Approximately 4.0 and 2.5 log reductions were observed for lettuce and chicken after 5 min of exposure, respectively, while 4.5 and 3.7 log reductions were achieved after 10 min of exposure. These results indicate higher bactericidal efficacy of CAP against SH on abiotic surfaces than on biotic surfaces, which is consistent with previous studies (Noriega et al. 2011; Fernández and Thompson, 2012; Fernández et al., 2013).

Survival curves of SH exposed to 2D-AICM-DBD on moist stainless steel discs, romaine lettuce, and chicken breast meat. The presented data are averages of triplicate experiments. The error bars represent standard deviations. Color images are available online.
Our results showed significant differences between the food samples themselves; inactivation rates of SH on lettuce at all CAP exposure times were significantly (p < 0.05) higher than those achieved on chicken breast muscles (Fig. 4). The type of treated food has previously been shown to affect the bactericidal efficacy of plasma. For instance, the efficacy of nitrogen CAP against Salmonella enterica serovar Typhimurium was higher on lettuce than on strawberry and potato (Fernández et al., 2013). Exposure to argon plasma generated in a corona-based setup killed Escherichia coli more easily on the surface of tomatoes than on lettuce and carrot (Bermúdez-Aguirre et al., 2013). The level of E. coli killed using remote, microwave-driven air plasma was higher on carrot than on apple slices (Baier et al., 2014).
Differences in CAP bactericidal efficacy between foods and abiotic surfaces and among foods themselves can be partially attributed to differences in the topographical features of the treated surface. Food surfaces can have irregularities and unsmooth features, while the stainless steel surface is flat and smooth. Different food structures (e.g., stomata and trichomes in lettuce and irregularities, cuttings, and punctures or cracked sites on the surface of chicken tissues and lettuce leaves) might induce a shielding effect for the CAP treatment, thereby protecting some bacterial cells from inactivation by CAP. Several studies have presented scanning electron micrographs (SEMs) of lettuce leaves' stomata entrapping cells of E. coli (Niemira and Cooke, 2010), Listeria monocytogenes (Ölmez and Temur, 2010), Salmonella Typhimurium (Fernández et al., 2013), and Aeromonas hydrophila (Jahid et al., 2014). Likewise, SEM pictures showed irregularities and fissures on the surface of chicken muscles entrapping Listeria innocua, which correlated with the lower efficacy of CAP compared with abiotic smooth surfaces such as membrane filters (Noriega et al., 2011).
We could not reach our objective of completely decontaminating lettuce and chicken muscle from SH at the initial bacterial counts used (∼7 × 106 CFUs/sample) within 10 min of treatment, which is needed in industrial sectors to meet the Salmonella zero-tolerance policy requirement (Hedberg, 2014). However, a reduction of ∼4.5 log (>99.99%) and 3.7 log (>99.95%) after 10 min of CAP exposure may still be of value since a decrease in the dose of the infectious agent may decrease the probability of illness (Teunis et al., 2010; McEntire et al., 2014).
Nevertheless, the level of Salmonella inactivation achieved on food samples using our plasma setup is much higher than that reported in earlier studies using different plasma sources. For instance, the maximum reduction of Salmonella Typhimurium on lettuce was only 2.72 log after 15 min of exposure to a nitrogen DBD plasma jet (Fernández et al., 2013) and 2.8 log after 10 min of exposure to the N2/O2 mixture-based microwave plasma system (Song et al., 2015). On lettuce, only 0.5 log of Salmonella enterica serovar Anatum, Salmonella enterica serovar Stanley, and Salmonella enterica serovar Enteritidis was inactivated by using an in-package DBD plasma system (Min et al., 2016). Similarly, only 2.5 log of Salmonella enterica serovar Typhi was inactivated on chicken breast using a DBD air plasma setup (Dirks et al., 2012).
Effect of proteins on CAP's bactericidal effect
Interaction of reactive species with functional proteins of target pathogens is one of the mechanisms by which CAP inactivates bacterial and viral pathogens (Aboubakr et al., 2016, 2018; Han et al., 2016). Tolouie et al. (2018) suggested that proteinaceous foods can interact with and consume a significant amount of CAP species. Lower vulnerability of SH on chicken than on lettuce and stainless steel seems to confirm these observations. To test this hypothesis, we measured inactivation of SH on stainless steel discs precoated with different concentrations of BSA aqueous solution. As seen in Figure 5, the presence of protein reduced the bactericidal effect of CAP with increasing protein concentration. This indicates that lower bactericidal efficacy of CAP on chicken is due to its proteinaceous nature, which might consume a certain amount of the reactive CAP species.

Survival curves of SH exposed to 2D-AICM-DBD on stainless steel discs coated with 1%, 5%, and 10% BSA. The presented data are averages of triplicate experiments. The error bars represent standard deviations. BSA, bovine serum albumin. Color images are available online.
Effect of initial SH counts on CAP efficacy
In practice, microbial contaminants on foods are usually present in relatively low numbers (Dufrenne et al., 2001; Oscar, 2013; Denis et al., 2016). In our study, we spiked samples with a high initial load of SH cells (approximately 6.00–6.5 log10 CFUs/sample). Several studies have indicated that the initial microbial cell count is a determining parameter for bactericidal efficacy of CAP treatment. This is because the high initial microbial load might lead to the formation of multilayers of the bacterial pathogen. In such situations, the top layers of bacterial cells, even if inactivated, could act as a physical barrier to protect those beneath them from plasma penetration (shadowing effect) and hence contribute to a decreased rate of inactivation (Deng et al., 2005; Kamgang-Youbi et al., 2008; Fernández et al., 2012). Therefore, we compared the efficacy of CAP treatment against SH at low (∼3.5 log10 CFUs/sample) and high initial counts (∼6.5 log10 CFUs/sample). Results show that the exposure time required for complete inactivation is determined by the quantity of bacterial cells contaminating the treated food or surface (Fig. 6). Although complete decontamination of stainless steel required 3 min of exposure at high bacterial counts, it was achieved in 0.5 min at low bacterial counts (Fig. 6A). Likewise, complete decontamination was achieved in 1 min of CAP exposure on lettuce with low bacterial cell counts, while it was unachievable with high bacterial counts during 10 min of CAP exposure (Fig. 6B). Complete decontamination of chicken breast was not achieved even at low initial bacterial counts (Fig. 6C). These findings indicate the utility of determining the bacterial contamination load on the food or surface before plasma treatment.
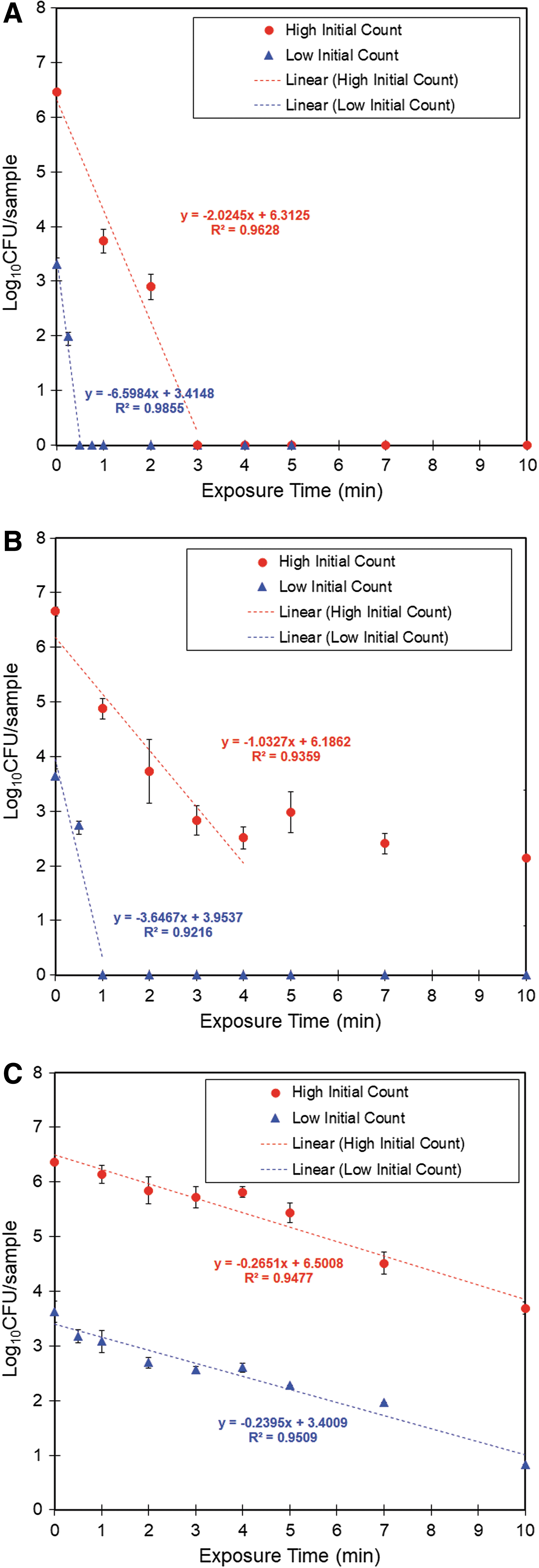
Survival curves of SH exposed to 2D-AICM-DBD on stainless steel discs
We also analyzed the kinetics of SH inactivation at low and high initial titers; D-values at low and high initial loads of SH were not significantly different in all sample types (Table 1). These results show that the initial counts of SH cells used in our study did not significantly affect the rate of bacterial killing (D-value). This finding is in contrast to that of Kamgang-Youbi et al. (2008) who found that initial bacterial loads affect the rate of bacterial inactivation. To explain this apparent contradiction, we calculated the cell density per spiked area of samples (1 cm2) as follows. With a cell diameter of Salmonella between about 0.7 and 1.5 μm and length from 2 to 5 μm (Fàbrega and Vila, 2013), the surface area of one cell on average is 3.85 ± 2.5 μm2. Assuming that the bacterial inoculum was homogeneously spread over the whole sample surface, forming a single layer of bacterial cells covering the sample requires 7.4 log10 CFUs/sample, which is much higher than the highest initial count of SH used in our study. This can explain the nonsignificant differences in rates of inactivation at low and high initial counts used in the present study. Similar findings were reported in a study on inactivation of spores of Bacillus subtilis by CAP (Deng et al., 2005) when the loads were under the stacking threshold. However, over that threshold, the rate of inactivation was reported to decrease significantly.
Kinetics of Salmonella enterica Serovar Heidelberg Inactivation by Exposure to Cold Atmospheric Plasma at Low and High Initial Counts
D-Values within each substrate that have similar superscript letters have no significant difference (p > 0.05). D-value was determined from the log-linear model [Eq. (1)] as D-value =
where Nt is the count of surviving bacterial cells after CAP treatment (expressed as log10 CFUs/sample), N 0 is the initial bacterial count, t is the plasma exposure time (min), and D-value (decimal reduction time or inactivation rate) is the CAP exposure time (min) required for reducing 1 log10 CFU of the initial count of SH.
In other words, D-value is the negative reciprocal of the slope of the log-linear regression line (Fig. 6).
CAP, cold atmospheric-pressure plasma; CFU, colony-forming unit; SH, Salmonella enterica serovar Heidelberg.
In these two cases (i.e., SH on romaine lettuce with a high count and SH on dry stainless steel), D-value was calculated from the negative reciprocal of the slope of the log-linear regression line at the linear part of the survival curves.
Conclusions
We report strong bactericidal efficacy of CAP generated by a 2D-AICM-DBD against an MDR strain of SH under moist surface conditions. The observed lower efficacy of CAP against bacteria applied on chicken breast muscles was probably due to consumption of CAP's reactive species by proteins present in chicken muscles. Although the time required for complete decontamination of SH on stainless steel decreased from 3 min to 30 s at high and low initial counts, and low counts of bacteria on lettuce could be inactivated within 1 min, the kinetics of bacterial killing showed no statistically significant effect of initial titer on the rate of inactivation under the investigated conditions. In general, the levels of bactericidal effects seen by the 2D-AICM-DBD setup are promising and higher than some of the previously reported CAP setups for similar treatment times. Finally, treatment with 2D-AICM-DBD did not result in any visual changes in the appearance of chicken breast and romaine lettuce. However, effects of CAP treatments on sensory and nutritional properties of the treated foods and the feasibility of CAP as a new intervention technology for food safety require further detailed studies.
Footnotes
Acknowledgments
P.J.B. acknowledges Kyocera, Inc., for providing the electrode array used in this research. The technical help of Dr. Hamed Taghvaei, Desiree Reding, and Taylor Wesfall-McCoy is gratefully acknowledged.
Disclosure Statement
No competing financial interests exist.
Funding Information
This project was partially funded by the Agriculture and Food Research Initiative of the USDA National Institute of Food and Agriculture, grant number 2017-67017-26172.