
Abstract
The mcr-1 gene has been identified in bacterial isolates obtained from humans, animals, environment, and food, including Salmonella spp., which is one of the major foodborne pathogens worldwide. The aim of this study was to evaluate the presence of mcr-1 gene in Salmonella spp. from food produced in Brazil and to characterize the isolates harboring this gene. A total of 490 Salmonella spp. isolates from the Brazilian National Program for the Control of Foodborne Pathogens were screened for the presence of mcr-1 gene by polymerase chain reaction (PCR). Whole genome sequencing (WGS) was performed in positive isolates to characterize the sequence type (ST), plasmid families and resistance genes. Antimicrobial susceptibility tests were performed by broth microdilution. Selected isolates were submitted to conjugation experiments using the Escherichia coli J53 as a receptor. We detected eight isolates harboring the mcr-1 gene; seven belonged to Salmonella enterica serovar Typhimurium and its monophasic variant 4,[5],12:i:-, and one belonged to serovar Saintpaul. Seven of the mcr-1 positive isolates displayed a high rate of resistance to other antibiotics in addition to colistin. Analysis of the WGS indicated that the ST 19 was the most common ST among the mcr-1 positive isolates. The mcr-1 gene was located in an IncX4 plasmid of ∼33 kb, with no additional resistance genes and with high identity with a plasmid obtained from a clinical isolate of E. coli mcr-1 positive in Brazil. All plasmids harboring the mcr-1 gene were able to conjugate. Our results suggest the spread of a single plasmid type in Brazil harboring the mcr-1 among Salmonella spp. The horizontal transfer of this mobile element has been contributing to the spread of the colistin resistance in the country.
Introduction
The use of polymyxins in human medicine has recently drawn more attention due to the increasing number of infections caused by carbapenemase producing Enterobacteriaceae in the previous years (Poirel et al., 2017). In fact, polymyxins have been used for decades in veterinary medicine and agricultural production (Poirel et al., 2017). Colistin (polymyxin E) is commonly prescribed for treatment and prevention of enteric diseases, mainly in poultry and pigs submitted to intensive husbandry systems. Furthermore, it has been extensively used as growth promoter worldwide (Poirel et al., 2017). In Brazil, the use of colistin as zootechnical additive in animal feed was prohibited only in November 2016 (Brazil, 2016).
Polymyxin resistance is mainly associated with the addition of cationic groups to lipopolysaccharide (LPS) in the outer membrane of Gram-negative bacteria, preventing antibiotic binding (Rhouma et al., 2016). It was believed that this mechanism was restricted to chromosomal mutations, which allowed only vertical transmission of this resistance determinant. However, in 2015, a plasmid transferable polymyxin resistance, mediated by the mcr-1 gene was described for the first time by Liu et al. (2016). The mcr-1 gene encodes a protein that is responsible for the addition of a phosphoethanolamine group to the lipid A, resulting in a more cationic LPS and conferring resistance to colistin (Liu et al., 2016).
The mcr-1 gene has been identified worldwide in bacterial isolates obtained from humans, animals, environment, and food (meat and vegetables), mostly in Escherichia coli but also in other Enterobacteriaceae, such as Enterobacter cloacae, Enterobacter aerogenes, Klebsiella pneumoniae, Shigella sonnei, and Salmonella enterica (Schwarz and Johnson, 2016; Poirel et al., 2017). Although there are already several variants of the mcr gene (mcr-1 to mcr-9) described in the literature (Carrol et al., 2019), the vast majority of the reports in Brazil are related to the mcr-1. In fact, in Brazil, mcr-1 was described in E. coli isolated from animals, inpatients, seawater, and food (Fernandes et al., 2016, 2017; Lentz et al., 2016; Monte et al., 2017; Pillonetto et al., 2019), in K. pneumoniae from hospitalized patients (Aires et al., 2017; Dalmolin et al., 2018), and in S. enterica from food (Rau et al., 2018; Moreno et al., 2019).
In view of the extensive use of colistin in the food production chain and the importance of Salmonella spp. as one of the major foodborne pathogens in the world, the aim of this study was to evaluate the presence of the mcr-1 gene in Salmonella spp. obtained from food produced in Brazil, as well as to characterize phenotypically and genotipically the isolates harboring this gene.
Materials and Methods
Bacterial isolates and identification procedures
A total of 490 Salmonella spp. isolates were randomly selected from the Brazilian National Program for the Control of Foodborne Pathogens of Ministry of Agriculture, Livestock and Food Supply. All the isolates were obtained according to ISO 6579:2002 procedures, from retail poultry, turkey and pork meat, produced in Brazil between 2014 and 2017, and from pork carcasses, in 2014 and 2015, comprising 154 different slaughterhouses under federal inspection in 140 cities and 13 states. Additional identification was performed by ribotyping, using RiboPrinter® System (DuPont Qualicon) and PvuII restriction enzyme, to determine the serotype of the isolates.
Identification of mcr-1 gene
Salmonella spp. isolates were evaluated for the presence of the mcr-1 gene by pooling seven isolates together and submitting them to DNA extraction and conventional polymerase chain reaction (PCR), with specific primers CLR5-F (5′-CGGTCAGTCCGTTTGTTC-3′) and CLR5-R (5′-CTTGGTCGGTCTGTAGGG-3′) as previously described (Liu et al., 2016). All isolates from a pool with mcr-1 positive result were retested individually by the same conventional PCR to identify the isolate(s) which presented the gene. For DNA extraction, pure colonies were transferred to TE buffer (Tris HCl 10 mM, EDTA 1 mM, pH 8.0) and submitted to heat shock at 80°C for 20 min, followed by freezing at −20°C. The amplicon of isolates with PCR positive for the mcr-1 gene was sequenced by Sanger method to confirm the mcr-1 gene sequence.
Antimicrobial susceptibility testing
Minimal inhibitory concentration (MIC) of 13 antimicrobials was determined by broth microdilution, according to ISO 20776-1:2006 procedures. The tested antibiotics included ampicillin, azithromycin, cefotaxime, ceftazidime, chloramphenicol, ciprofloxacin, gentamicin, meropenem, nalidixic acid, sulfamethoxazole/trimethoprim, tetracycline, tigecycline, and colistin. Results were interpreted according to European Committee on Antimicrobial Susceptibility Testing (EUCAST) clinical breakpoints (2019—version 9.0). For those antibiotics which do not have defined breakpoints for Salmonella spp., the results were interpreted with the EUCAST epidemiological cut-off values. E. coli ATCC 25922, Staphylococcus aureus ATCC 29213, and Pseudomonas aeruginosa ATCC 27853 were used as quality controls.
Whole-genome sequencing
Total DNA was extracted with the Wizard® Genomic DNA Purification Kit (Promega Corporation). Libraries were prepared with Nextera XT DNA Sample Preparation Kit and sequenced on an Illumina MiSeq analyzer using a 250 bp paired-end library, according to the manufacturer's instructions. Sequences were assembled on SPAdes Assembly version 3.9 (Nurk et al., 2013). Genome annotation was carried out using the RAST Server (Rapid Annotations using Subsystems Technology) version 2.0 (Aziz et al., 2008). Multilocus sequence typing (MLST) and plasmid incompatibility family were determined using the Center for Genomic Epidemiology tools (MLST 2.0 and PlasmidFinder 2.0), as well as acquired antimicrobial resistance (AMR) genes and chromosomal point mutations (ResFinder 3.1) (Larsen et al., 2012; Zankari et al., 2012; Carattoli et al., 2014). Detailed analyses were performed in Geneious v. 11.0.4 software.
Conjugation experiments
Plasmid transferability was evaluated by conjugation experiments using E. coli J53. Each of mcr-1 positive isolates was incubated with the E. coli J53 on a cellulose membrane, in a Luria-Bertani agar plate. After incubation at 37°C for 24 h, colistin-resistant transconjugants were selected on culture media supplemented with azide (100 mg/mL) and colistin (0.5 μg/mL).
Results
Prevalence and characteristics of mcr-1 positive S. enterica
We detected 8 (1.6%) positive isolates from the 490 Salmonella spp. screened by PCR for the presence of mcr-1 gene. Most (5) isolates mcr-1 positive were obtained from pork carcasses during 2015 (Table 1). All mcr-1 positive isolates presented 100% identity to mcr-1 gene described by Liu et al. (2016). According to ribotyping, the isolates belonged to serovar Typhimurium and its monophasic variant 4,[5],12:i:-, except for the isolate SLRe1401, which belonged to serovar Saintpaul. Phylogeny analysis of mcr-1 positive isolates indicated that there was no correlation between clusters and the epidemiological data (data not shown).
Epidemiological Features and Phenotypic and Genotypic Characterization of mcr-1 Positive Isolates of Salmonella enterica from Brazil
AMP, ampicillin; AMR, antimicrobial resistance; AZM, azithromycin; CHL, chloramphenicol; CIP, ciprofloxacin; CST, colistin; GEN, gentamicin; MIC, minimal inhibitory concentration; MLST, multilocus sequence typing; NAL, nalidixic acid; ST, sequence type; SXT, sulfamethoxazole/trimethoprim; TET, tetracycline; TGC, tigecycline.
Antimicrobial susceptibility
Antimicrobial susceptibility testing indicated that most of the mcr-1 positive isolates displayed resistance to other antibiotics in addition to colistin; most isolates were resistant to ampicillin (seven isolates), tetracycline (six isolates), nalidixic acid (five isolates), chloramphenicol (five isolates), ciprofloxacin (five isolates), and gentamicin (five isolates) (Table 1). In contrast, none of the isolates was resistant to cefotaxime, ceftazidime, or meropenem. MIC for colistin varied from 4 to 8 mg/L, which were just above the breakpoint to be classified as resistant by the EUCAST (Table 1).
Genetic characterization and plasmid analysis
Genome analysis using Center for Genomic Epidemiology tools revealed that six isolates belonged to the sequence type (ST)19 and two isolates (SLRe1401 and SLRe1503) belonged to ST50 and ST4556, respectively. Different types of plasmids (Table 1) were identified among the mcr-1 positive isolates by the in silico analysis of whole-genome sequencing (WGS) data. However, all eight isolates carried an IncX4 plasmid harboring the mcr-1 gene. No additional resistance genes were found in IncX4 plasmids. Nevertheless, several other acquired antibiotic resistance-encoding genes, as well as chromosomal point mutations, were identified according to ResFinder 3.1; these genetic determinants were associated to resistance to aminoglycoside, β-lactam, fluoroquinolone, sulfonamide, and tetracycline classes. None of the isolates had mutations in the pmrAB, phoPQ, or mgrB genes associated with colistin resistance.
Detailed WGS analysis, performed in Geneious software, indicated that the IncX4 plasmids presented ∼33 kb. The insertion sequence ISApl1 was not found neither upstream nor downstream the mcr-1 gene. However, the mcr-1-pap2 sequence and the transposase IS26 were present in all IncX4 plasmids analyzed. Alignment with pICBEC72Hmcr, a plasmid obtained from a clinical isolate of E. coli mcr-1 positive from Brazil (GenBank CP015977.1), indicated 99% of identity (Fig. 1).
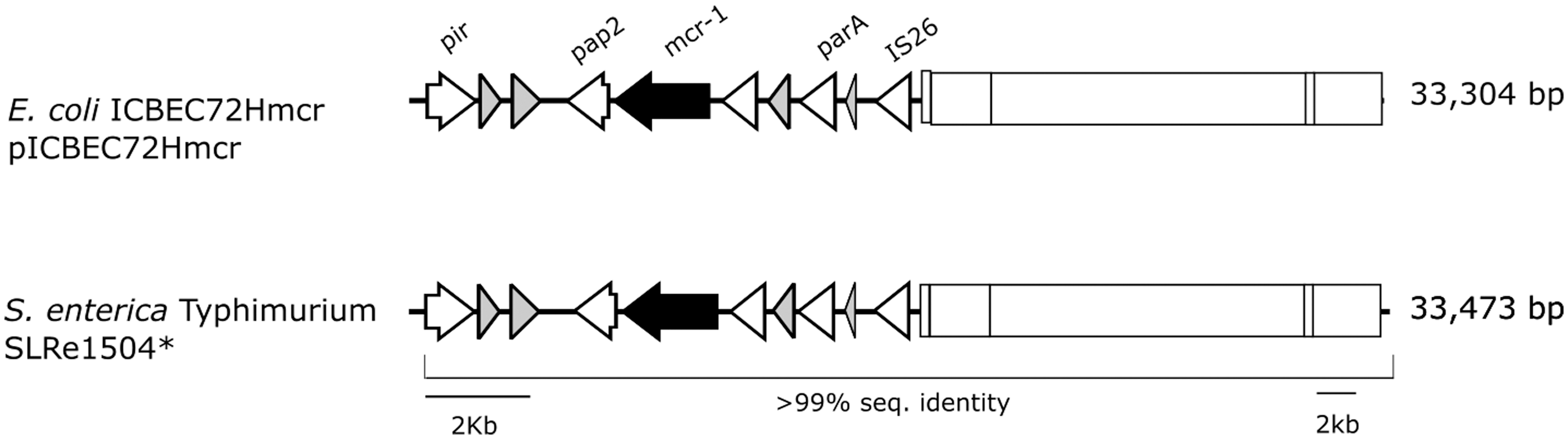
Genomic context of IncX4 mcr-1 carrying plasmid from isolate SLRe1504 compared to pICBEC72Hmcr (GenBank CP015977). Plasmid region represented by arrows shows sequence details close to mcr-1 gene insertion. Plasmid region represented by rectangle shows sequences related to plasmid functionality. Gray arrows represent hypothetical proteins. * The plasmid from isolate SLRe1504 was used as reference. The other mcr-1 carrying plasmids described in this study presented the same gene organization.
Transfer of mcr-1 gene
To investigate the transference potential of the plasmids harboring the mcr-1 gene, four of the S. enterica isolates were submitted to conjugation experiments using the E. coli J53 as a recipient strain. Results demonstrated that all plasmids were able to conjugate. The presence of the mcr-1 gene in the transconjugants was confirmed by PCR.
Discussion
In this study, we have detected and confirmed the presence of the mcr-1 gene in eight Salmonella isolates from meat (turkey, poultry, and pork origin), belonging to two different serotypes. Our data showed that most of the isolates (seven isolates) which harbored the mcr-1 gene were Salmonella Typhimurium or its monophasic variant 4,[5],12:i:-. Although mcr-1 bearing plasmids have been identified in different Salmonella serovars, Typhimurium seems to be the most often serotype associated with this resistance gene. This association still needs further investigation; however, it may indicate the requirement of a specific genetic background for acquisition and maintenance of mcr-1 harboring plasmids (Doumith et al., 2016; Li et al., 2016b; Cui et al., 2017a).
The antimicrobial susceptibility profile varied among different Salmonella spp. mcr-1 positive isolates, although the vast majority of isolates (except isolate SLRe1401) displayed a multidrug resistance profile, being resistant to three or more classes of antibiotics (Magiorakos et al., 2012). In fact, multiple resistance genes were detected by in silico analysis of WGS in most isolates. Although the frequency of multiresistance varies in different Salmonella serotypes, Salmonella Typhimurium is commonly associated to this pattern worldwide (Leekitcharoenphon et al., 2016; Mąka and Popowska, 2016; Wang et al., 2017). It is important to highlight that none of the isolates was resistant to third-generation cephalosporins or to carbapenems, which are considered critically important antimicrobials for human medicine, according to the World Health Organization (WHO, 2016).
Regarding the genetic context, MLST analysis revealed that most isolates belonged to ST19. Salmonella spp. mcr-1 positive of this ST has been described only in Denmark and in Spain (Torpdahl et al., 2017; Lozano-Leal et al., 2019). Of note, most reports of mcr-1 Salmonella Typhimurium are associated to ST34, including reports from Asia, Europe, and the only reported case in Latin America, in Colombia (Doumith et al., 2016; Li et al., 2016b; Saavedra et al., 2017). ST19 differs from ST34 by a single nucleotide change in the dnaN locus (Li et al., 2017).
To the best of our knowledge, this is the first report of ST50 in Salmonella Saintpaul and the ST4556 in Salmonella Typhimurium harboring the mcr-1 gene. ST4556 has a single nucleotide change in the sucA locus when compared to ST19.
Several plasmid incompatibility groups have already been associated with the mcr-1 gene. However, the global dissemination seems to be associated with three major groups: IncX4, IncI2, and IncHI2 (Matamoros et al., 2017). Among all of our isolates, the mcr-1 gene was located in IncX4 plasmids, with ∼33 kb. IncX4 plasmids have been reported in different Salmonella serotypes, mainly in Europe (Campos et al., 2016; Veldman et al., 2016; Torpdahl et al., 2017; Garcia-Graells et al., 2018) and Asia (Chiou et al., 2017; Cui et al., 2017b). There are no reports of IncX4 plasmids harboring mcr-1 gene in Salmonella isolates in Latin America, except in Brazil (Moreno et al., 2019). Indeed, all mcr-1 reports in Brazil are associated with IncX4 plasmids, including E. coli from humans, foods, and environment (Fernandes et al., 2016, 2017; Monte et al., 2017; Pillonetto et al., 2019), and K. pneumoniae from inpatients (Aires et al., 2017; Dalmolin et al., 2018).
The high identity between the IncX4 plasmids harboring mcr-1 gene from our study and pICBEC72Hmcr (GenBank CP015977.1) suggests the spread of a single plasmid type in Brazil, probably of the same origin, which can be found in different bacteria from distinct sources. This fact underlines the potential for horizontal transfer of the mcr-1 through the IncX4 plasmids. However, in silico analysis showed the absence of the mobile element ISApl1 in all of our plasmids (the same was found in pICBEC72Hmcr). This insertion sequence is associated with the mobilization and dissemination of mcr-1 gene among different bacterial strains and species, but is not present in IncX4 plasmids (Luo et al., 2017). The exact mechanism related to the dissemination of polymyxin resistance by IncX4 plasmids requires more investigation, but it seems to be associated with the mcr-1-pap2 sequence (Li et al., 2016a), which was present in all eight isolates that we have sequenced, near the transposase IS26.
Conjugation experiments demonstrated that all the four Salmonella isolates tested were able to transfer their mcr-1 bearing plasmids to E. coli J53, which is in concordance with earlier findings about the conjugative nature of IncX4 plasmids (Campos et al., 2016; Cui et al., 2017a, b).
Conclusions
We reported eight mcr-1 positive Salmonella isolates from food in Brazil. Our results suggest the spread of a single plasmid type in Brazil harboring the mcr-1 gene, indicating that horizontal transfer of this mobile genetic element has been contributing to the spread of the colistin resistance in the country. The presence of mcr-1 gene in a relevant foodborne pathogen emphasizes the importance of monitoring the AMR in the food-production chain and the need of promoting the responsible use of antibiotics in agriculture and livestock.
Sequence Data
Sequences were deposited to GenBank under accession numbers RXNI00000000, RXNJ00000000, RXNK00000000, RXNL00000000, RXNM00000000, RXNN00000000, RXNO00000000, and RXNP00000000.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by “Instituto Nacional de Pesquisa em Resistência a Antimicrobianos” (INPRA), by Ministry of Agriculture, Livestock and Food Supply from Brazil (MAPA) and by “Fundo de Incentivo à Pesquisa” (FIPE/HCPA 17-0541).