
Abstract
Polyphenols are a group of active ingredients in olive oil, and have been reported to exhibit antioxidant activity. Salmonella enterica subsp. enterica serovar Typhimurium (Salmonella Typhimurium) and Staphylococcus aureus are common foodborne pathogens causing serious infections and food poisoning in humans. This study was conducted to analyze the antibacterial activity of olive oil polyphenol extract (OOPE) against Salmonella Typhimurium and S. aureus, and reveal the possible antibacterial mechanism. The antibacterial activity was estimated using minimum inhibitory concentration (MIC) values and bacterial survival rates when treated with OOPE. The antibacterial mechanism was revealed through determinations of changes in intracellular ATP concentration and cell membrane potential, sodium dodecyl sulfate–polyacrylamide gel electrophoresis, and transmission electron microscopy analysis. The results showed the MICs of OOPE against Salmonella Typhimurium and S. aureus were 0.625 and 0.625–1.25 mg/mL, respectively. The growth of Salmonella Typhimurium and S. aureus (∼8 log CFU/mL) was completely inhibited after treatments with 0.625 mg/mL of OOPE for 3 h and 0.625–1.25 mg/mL for 5 h, respectively. When Salmonella Typhimurium and S. aureus were exposed to OOPE, the physiological functions associated with cell activity were destroyed, as manifested by reduction of intracellular ATP concentrations, cell membrane depolarization, lower bacterial protein content, and leakage of cytoplasm. These findings suggested a strong antibacterial effect of OOPE against Salmonella Typhimurium and S. aureus, and provided a possible strategy of controlling contamination by these two pathogens in food products.
Introduction
Foodborne illnesses are a widespread and growing public health problem in both developed and developing countries. It is estimated that 44% of foodborne diseases are caused by foodborne pathogenic microorganisms, among which Salmonella and Staphylococcus aureus are responsible for a large number of severe foodborne illnesses (Nyachuba, 2010; Rezaei et al., 2015; Weerakkody et al., 2010). Salmonella and S. aureus could contaminate food utensils and a variety of food sources with a high rate of colonization, resulting in rising global mortality rates and a heavy public health burden (Tong et al., 2015; Andoh et al., 2017). Although many measures have been taken in the food industry to reduce the contamination of Salmonella and S. aureus strains, occurrence of contamination by these two pathogens remains significant (Erickson et al., 2018; Papadopoulos et al., 2018; Wang et al., 2018).
Addition of synthetic preservatives was considered to be an effective and common method to control microbial contamination of food products. However, considering the possible potential risk of chemical preservatives, consumers, food companies, government regulators, and official control authorities are more willing to accept the use of natural antimicrobial extracts to inhibit the growth of microorganisms in food (Li et al., 2015). In this context, several natural substances have been proposed to replace synthetic preservatives to inhibit the growth of pathogenic bacteria (Parthasarathy et al., 2009; Naili et al., 2010; Chen et al., 2017). Naili et al. (2010) reported that polyphenolic extracts of two Libyan folk medicinal plants have strong antibacterial activity against Salmonella typhi and S. aureus. Parthasarathy et al. (2009) found the Mitragyna speciosa leaf extracts containing phenolics and flavonoid could effectively inhibit the growth of S. typhi and Bacillus subtilis, with minimum inhibitory concentrations (MICs) from 3.12 to 6.25 mg/mL. In addition, sugar beet molasses polyphenols showed a good bacteriostatic effect against S. aureus, Listeria monocytogenes, Escherichia coli, and Salmonella Typhimurium. These suggested that natural products can be used as substitutes for synthetic preservatives to inhibit foodborne pathogen growth, resulting in extending shelf life of food and reducing the risk of foodborne disease outbreaks.
Olive oil polyphenol extract (OOPE) is composed of polyphenols, hydroxytyrosol, tyrosol, organic acids, and sugars, and it has been demonstrated to exhibit strong antioxidant activity (Gordon et al., 2001; Hernáez et al., 2017). The antioxidant effect in tea polyphenols, blueberry proanthocyanidins, cranberry extracts, and impatiens polyphenols is reported to be the most important contributor to microbial inhibition (Mishra et al., 2011; Joshi et al., 2014; Shi et al., 2016c; Kim et al., 2018). Therefore, it is reasonable to hypothesize that OOPE has a similar antibacterial effect against foodborne pathogens. In this study, the antibacterial activity of OOPE against both Salmonella enterica subsp. enterica serovar Typhimurium (Salmonella Typhimurium) and S. aureus was evaluated on the basis of MIC values and survival of bacteria after treatment with OOPE in sterile normal saline (NS) and Luria-Bertani (LB) broth. Further analyses of changes in intracellular ATP concentration, membrane potential, bacterial protein, and cell morphology were used to explore the mechanism of action of the observed antibacterial activity.
Materials and Methods
Olive oil polyphenol extract
OOPE was purchased from Shanghai Kai Da Biotechnology Co. Ltd. (Shanghai, China). The polyphenols were extracted from olive oil using supercritical fluid extraction method, and ethanol was used as an extraction agent. The extracts were concentrated in a rotary evaporator. The detailed chemical compositions are as follows: moisture (<7%), total polyphenols (≥30%), hydroxytyrosol (≥6%), tyrosol (≥0.8%), phenolic acids (≥1.5%), sugars (<40%), and residual solvent content ethanol (<0.1%).
Bacterial strains and culture conditions
Salmonella Typhimurium ATCC 14028 and S. aureus ATCC 13565 were obtained from American Type Culture Collection (ATCC, Manassas, VA). Eight other Salmonella Typhimurium isolates and six other S. aureus isolates were provided by Key Laboratory of Dairy Science at Northeast Agricultural University (Harbin, China), which were isolated from a variety of food sources, including fresh chicken, pork, beef, and milk. All isolates and ATCC strains were used to assess the MIC, whereas Salmonella Typhimurium ATCC 14028 and S. aureus ATCC 13565 were used to analyze the antibacterial mechanism. All cultures of isolates were stored in 40% glycerol at −80°C. Working cultures were prepared by transferring 300 μL of frozen stocks of each strain into 10 mL of LB broth and incubating at 37°C for 20 h.
MIC determination
The MICs of OOPE against Salmonella Typhimurium and S. aureus ATCC strains and isolates were determined according to the agar dilution method as described by the European Committee for Antimicrobial Susceptibility Testing (EUCAST) of the European Society of Clinical Microbiology and Infectious Diseases (2000). Ampicillin was chosen as the reference antibiotic. All tested strains and strains were treated with different concentrations of OOPE (0, 0.3125, 0.625, 1.25, 2.5, 5, and 10 mg/mL) and 0.1 mg/mL ampicillin. The bacterial cultures with 2 μL (104 CFU) were individually dripped on solid medium tryptic soy agar, and then were incubated at 37°C for 24 h. The lowest concentration of OOPE that completely inhibited the test strain growth was identified as the MIC for that particular isolate or strain.
Reduction of Salmonella Typhimurium ATCC 14028 and S. aureus ATCC 13565 by OOPE
Cell density of the tested cultures was adjusted to 108 CFU (colony-forming unit)/mL using sterile NS. OOPE was dissolved in the aforementioned working cultures to obtain the final concentrations of 0.625 and 1.25 mg/mL for Salmonella Typhimurium, and 1.25 and 2.50 mg/mL for S. aureus. Untreated Salmonella Typhimurium and S. aureus cells were used as negative controls. The strains treated with OOPE in NS were incubated at 37°C for 0.5, 1, 3, and 5 h. Cell counts and the survival rate of each strain were obtained and calculated.
Measurement of intracellular ATP concentrations
Changes in intracellular ATP concentrations of the two strains treated with 1 MIC and 2 MIC of OOPE at 37°C for 30 min were analyzed as described by Sánchez et al. (2010) and Shi et al. (2016a). The untreated Salmonella Typhimurium ATCC 14028 and S. aureus ATCC 13565 cells were used as negative controls. The intracellular ATP concentrations of samples were detected using an ATP assay kit (Beyotime Bioengineering Institute, Shanghai, China) according to the manufacturer's instructions. In addition, to reduce the loss of ATP, the supernatant used to determine intracellular ATP concentrations was prepared at 0°C using a high-speed freezing centrifuge (Shanghai Centrifuge Institute Co., Ltd., Shanghai, China), and then stored in ice box for the next extraction of ATP.
Determination of membrane potential
The membrane potential of the two strains after treatment with 1 MIC and 2 MIC of OOPE at 37°C for 30 min was measured according to previous reports with minor modifications (Shan et al., 2007; Fei et al., 2018). Untreated cells were used as negative controls. Bis-(1,3-dibutylbarbituric acid) trimethine oxonol [DiBAC4 (3); Beijing Solarbio Sciences and Technology Co. Ltd., Beijing, China] was used as a membrane potential-sensitive fluorescent probe. Cells were grown in NS at 37°C to an OD600 of 0.5 and centrifuged at 5000 × g for 5 min. Bacterial suspension after being washed twice with phosphate-buffered saline (pH 7.0) and DiBAC4 (3) fluorescent probe was added to a black, opaque 96-well plate (Corning Institute) to balance at 37°C for 30 min. OOPE was added to achieve final concentrations of 0 MIC, 1 MIC, and 2 MIC. Fluorescence was recorded at the excitation and emission wavelengths of 492 and 515 nm, respectively, using a fluorescence microplate reader (Infinite 200 PRO; Tecan, Grodlg, Austria).
Sodium dodecyl sulfate–polyacrylamide gel electrophoresis observation
To investigate the effect of OOPE on bacterial proteins, the sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) analysis was performed. Approximately 107 CFU/mL of the tested strains was treated by 1 MIC of OOPE at 37°C for 3, 6, 9, and 12 h, respectively, whereas untreated tested strains were used as negative controls.
The SDS-PAGE process was performed in accordance with the method provided by Chen et al. (2017).
Gels were dyed with 0.1% Coomassie Brilliant Blue R250 and destained with glacial acetic acid and methanol in distilled water. After decoloration, photographs of protein gels were taken using the Gel DocTM EZ imaging system (Bio-Rad, CA).
Transmission electron microscopy observation
To reveal the effects of OOPE on the cell morphology of tested strains, transmission electron microscopy (TEM) observation was performed according to previous reports (Li et al., 2015; Fei et al., 2019). Salmonella Typhimurium and S. aureus were treated with 1 MIC and 2 MIC of OOPE for 4 h. The working cells were prepared after centrifugation at 5000 × g at 4°C for 5 min, and fixed with 0.1 M sodium phosphate buffer (2.5% glutaraldehyde) at 4°C for 12 h followed by washing thrice with NS. The samples were kept in 1% osmium tetroxide at 4°C for 2 h. After washing thrice with NS, the cells were dehydrated with a graded series of ethanol (50%, 70%, 90%, and 100%) for 10 min, and embedded in Epon Lx-112 (Ladd Reseaerch, Williston, VT) followed by double staining with uranyl acetate and lead citrate. Finally, all samples were observed using a transmission electron microscope (Hitachi, Tokyo, Japan) to obtain their TEM pictures.
Statistical analysis
All experiments were carried out in triplicate. Data were expressed as mean values ± standard deviations. The date of the survival counts of bacteria, ATP concentrations, and membrane potentials were tested with both analysis of variance (ANOVA) and Duncan's test. These two data analysis methods are generally applied to significance analysis and were used in our study to get a more rigorous data analysis. ANOVA was conducted using SPSS 22.0 software (SPSS, Inc., Chicago, IL). Duncan's multiple range test was used to identify significant differences among the means (p < 0.05).
Results
MICs of OOPE on Salmonella Typhimurium and S. aureus strains and isolates
The MICs of OOPE on Salmonella Typhimurium and S. aureus strains and isolates are shown in Table 1. The MICs of OOPE on the strain and all the eight isolates of Salmonella Typhimurium strains were 0.625 mg/mL. Meanwhile, the MICs of OOPE against S. aureus KLDS-G-4 and the remaining five S. aureus strains and one strain were 0.625 and 1.25 mg/mL, respectively.
Minimum Inhibitory Concentrations of Olive Oil Polyphenol Extract Against Nine Salmonella Typhimurium and Seven Staphylococcus aureus Strains and Isolates
MIC, minimum inhibitory concentration.
Reduction of Salmonella Typhimurium ATCC 14028 and S. aureus ATCC 13565 by OOPE
The survival counts of Salmonella Typhimurium and S. aureus after treatments with OOPE were evaluated (Fig. 1). The growth of Salmonella Typhimurium (∼8 log CFU/mL) was completely inhibited after treatments with 1 MIC of OOPE for 3 h and 2 MIC of OOPE for 1 h (Fig. 1A). Meanwhile, the survival counts of S. aureus treated with OOPE at 1 MIC for 5 h and 2 MIC for 3 h could not be detected (Fig. 1B).
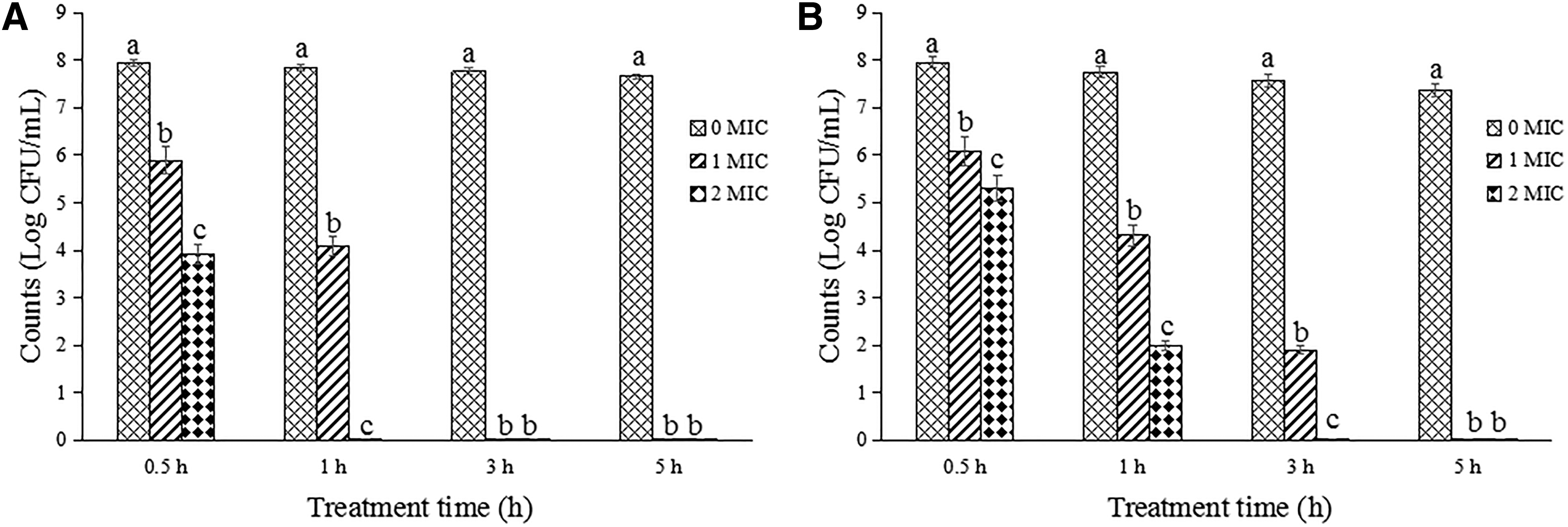
Reduction of Salmonella Typhimurium ATCC 14028 and Staphylococcus aureus ATCC 13565 after treatments with OOPE at different concentrations.
Changes in intracellular ATP concentrations
The intracellular ATP concentrations of Salmonella Typhimurium ATCC 14028 and S. aureus ATCC 13565 strains were measured after being treated with OOPE (0 MIC, 1 MIC, and 2 MIC) for 0.5 h (Fig. 2). Compared with control groups, the cells treated with OOPE appeared a significant reduction on intracellular ATP concentrations (p < 0.05). No significant difference in intracellular ATP concentrations was found between the samples treated with OOPE at 1 MIC and 2 MIC (p > 0.05).
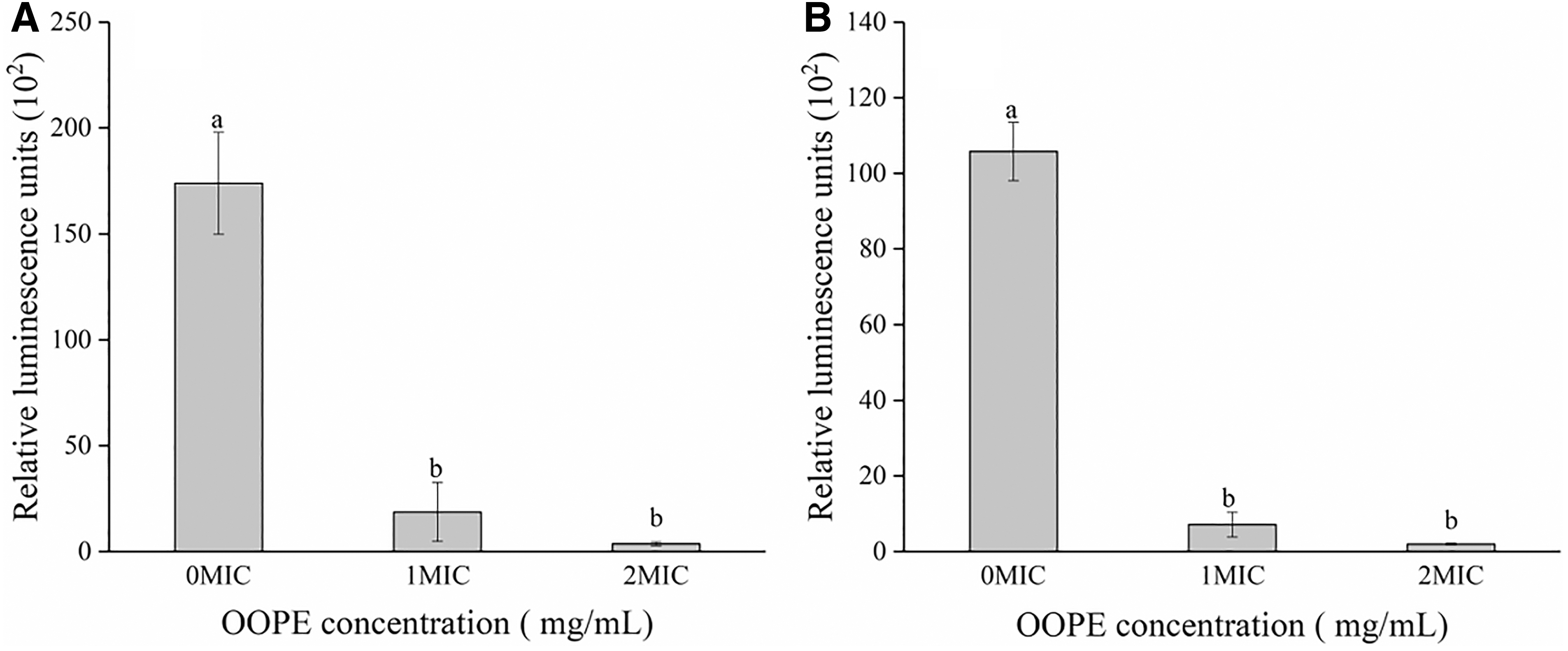
Change in intracellular ATP concentrations of Salmonella Typhimurium ATCC 14028
Changes in membrane potential
The effects of OOPE on membrane potential of Salmonella Typhimurium ATCC 14028 and S. aureus ATCC 13565 were assessed. Fluorescence produced by DiBAC4 (3) for treated cells was significantly increased compared to that of control cells (p < 0.05) (Fig. 3). Because relative fluorescence increase means a depolarization of the membrane, it suggests that both pathogens displayed cell membrane depolarization induced by OOPE.

Change in membrane potential of Salmonella Typhimurium ATCC 14028
SDS-PAGE analysis
The changes in bacterial protein of Salmonella Typhimurium and S. aureus after a treatment with 1 MIC of OOPE for 12 h were elucidated using SDS-PAGE analysis (Fig. 4). After exposure to OOPE, the protein bands of Salmonella Typhimurium and S. aureus became much fainter compared with those of untreated samples. It is evident that all protein bands of Salmonella Typhimurium treated with different MIC of OOPE disappeared from 8 h onward (Fig. 4A).
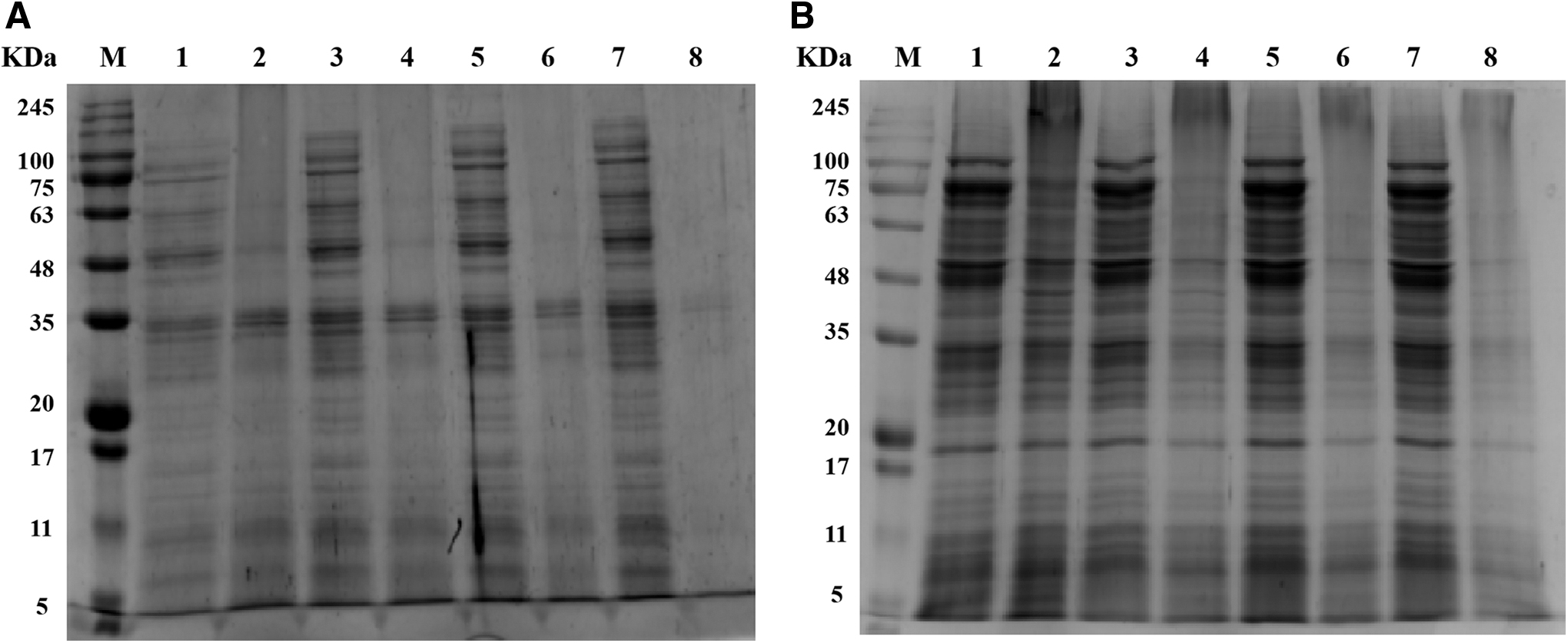
SDS-PAGE analysis of Salmonella Typhimurium ATCC 14028
TEM observation
TEM observation is a powerful tool for investigators to reveal the effects of stressors on bacterial cells (Xing et al., 2009). The changes in cell morphology of Salmonella Typhimurium ATCC 14028 and S. aureus ATCC 13565 after treatments with 1 MIC and 2 MIC of OOPE for 4 h were observed using TEM analysis (Fig. 5). Compared to untreated cells (Fig. 5A1, B1), cells treated with OOPE displayed a serious deformation and cytoplasmic leakage (Fig. 5A2, A3, B2, B3). These results suggested that treatment of OOPE could increase cell membrane penetrability and cause significant alteration in cell morphology.
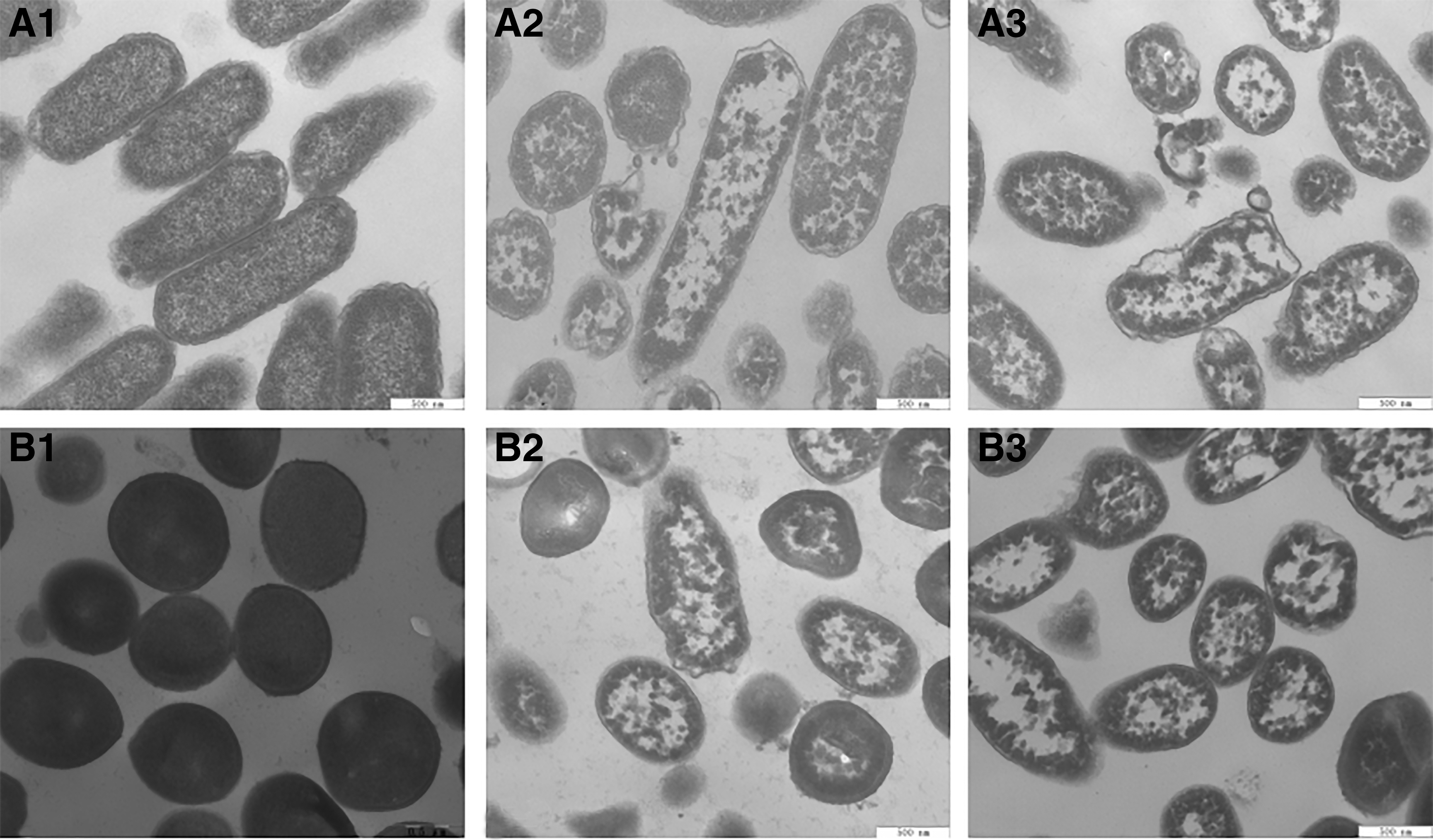
TEM images of Salmonella Typhimurium ATCC 14028 and Staphylococcus aureus ATCC 13565 cells (40,000 × ).
Discussion
Antibacterial effects of various natural plant extracts against Salmonella Typhimurium and S. aureus strains had been reported (Chen et al., 2017; Kittisakulnam et al., 2017; Kim et al., 2018). MICs of sugar beet molasses polyphenols on Salmonella Typhimurium and S. aureus were 1.25 and 2.5 mg/mL, respectively. Kittisakulnam et al. (2017) revealed 8.8 mg/mL of garlic extract and 9.3 mg/mL of pepper extract were the lowest concentration to prevent growth of S. aureus strains. Rice hull smoke extracts could effectively inhibit the growth of Salmonella Typhimurium with a 1.29% (v/v) MIC (Kim et al., 2018). Therefore, the antibacterial effects of OOPE on Salmonella Typhimurium (MIC = 0.625 mg/mL) and S. aureus (0.625–1.25 mg/mL) were superior comparable to previously identified antibacterial natural extracts. This is because, compared with other phenols, OOPE contains hydroxytyrosol and tyrosol, which have been shown to increase the antibacterial effect of OOPE (Fei et al., 2018).
ATP is associated with cellular signaling and metabolic regulation. The synthesis of ATP also played a key role to supply the energy generating in a microorganism. So the level of cellular ATP is an important target for evaluating the available energy and then response to the antibacterial effects of natural substance (Shi et al., 2016c). Intracellular ATP concentration of S. aureus treated with Helichrysum italicum oil was significantly reduced, which was suggested to be one of the main reasons for the antibacterial effect of H. italicum oil (Cui et al., 2015). Diminished ATP concentration could be caused by release of cytoplasmic ATP and unabated hydrolysis by the proton-pumping ATPase, which leads to rapid depletion of the intracellular ATP (Sánchez et al., 2010).
In this study, OOPE induced depolarization of Salmonella Typhimurium and S. aureus cells, while inhibiting cell growth. Similarly, cell membrane depolarization occurred in Vibrio cholerae cell treated by nopal cactus methanolic extracts and C. sakazakii cell treated by lipoic acid (Shi et al., 2016a, b). On the contrary, citral, syringic acid, and sweet acacia extracts were reported to lead to cell membrane hyperpolarization (Shan et al., 2007; Shi et al., 2016a, c). The depolarization and hyperpolarization of the cell membrane are a stress–response mechanism of the cells upon stimulation. Cell membrane depolarization was associated with Na+ flow into the cell after opening Na+ channel on the membrane, causing less negative charge inside the cell, whereas cell membrane hyperpolarization was attributed to the leakage of K+ from cells, resulting in an increase of negative charge inside the cell (Gries et al., 2013).
Bacterial proteins as essential cell components are closely related to cell vitality and the important physiological functions (Xu et al., 2017). In this study, after treatment with OOPE, the bands in SDS-PAGE profiles of Salmonella Typhimurium and S. aureus cells were significantly darkened or even disappeared. Similar findings have been reported in the antibacterial action of sugarcane bagasse extract against S. aureus, and sugar beet molasses polyphenols against S. aureus, L. monocytogenes, E. coli, and Salmonella Typhimurium (Gries et al., 2013; Chen et al., 2017). Thus, it suggests that in the process of inhibiting bacteria with natural extracts, the protein of tested strain was lost, or protein synthesis and related gene expression were suppressed (Fei et al., 2018).
The destruction of cell morphology is the most important reason for natural products to exert antibacterial effect, and the damaged cell morphology and increased cell membrane permeability are also reasons for the decrease of bacterial intracellular ATP and protein concentration (Fei et al., 2019; Guo et al., 2019). In this study, OOPE significantly damaged cell morphology, including collapse of cells, leakage of cell fluid, and cell fragmentation. It is similar to the conclusions from previous studies, where obvious cell deformation was used to explain the antibacterial mechanism of tea polyphenol, sugar beet molasses polyphenols, and punicalagin against foodborne pathogenic bacteria (Li et al., 2015; Chen et al., 2017). In addition, in report of Joshi et al. (2014), blueberry proanthocyanidins did not lead to the changes in cell morphology and leakage of cell fluid, but produced blebs and visible pores on the surface of cells, which is probably why OOPE has a bactericidal effect on the tested cells, while blueberry proanthocyanidins has only a bacteriostatic effect.
Conclusion
In conclusion, OOPE has an effective antibacterial activity against Salmonella Typhimurium and S. aureus with MICs value of 0.625 and 0.625–1.25 mg/mL. The antimicrobial effects of OOPE against these two pathogens were associated with reduction of intracellular ATP concentrations, cell membrane depolarization, a decrease in bacterial protein content, and leakage of the cytoplasm. These findings indicate that OOPE can be used as a natural preservative and antibiotic to reduce the risk of Salmonella Typhimurium and S. aureus in food processing. However, further toxicological evaluations in vitro and in vivo, determination of dosage, and effects on the sensory characteristics of foods are necessary before OOPE could be applied as a preservative in food.
Footnotes
Acknowledgment
We thank Professor Xinhuai Zhao for his help in experiment design.
Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported by the Open Fund of the Key Laboratory of Dairy Science of the Ministry of Education at Northeast Agricultural University (2017KLDS06), Key Scientific and Technological Project of Henan Province (161100110900), and National Key Research and Development Program of China (2017YFC1601904-02).