
Abstract
Shiga toxin–producing Escherichia coli (STEC) are a leading cause of foodborne illnesses worldwide, with beef and beef products as a common food reservoir. STEC strains may be present in beef-processing environments in the form of biofilms. The exudate of raw beef, also referred to as beef juice, has been identified as an important source of bacterial contamination on food-processing surfaces. This study applied beef juice as a food-based model to study its effects on biofilm formation of six STEC isolates on stainless steel. Crystal violet staining and cell enumeration demonstrated that beef juice inhibited the biofilm formation of strains O113, O145, and O91 up to 24 h at 22°C, but that biofilm increased (p < 0.05) thereafter over 72 h. Biofilms formed by O157, O111, and O45 were not affected by the addition of beef juice over the whole incubation period. Electron microscopy showed that the morphology of biofilm cells was altered and more extracellular matrix was produced with beef juice than with M9 medium. The present study demonstrated that beef juice residues on stainless steel can enhance biofilm formation of some STEC strains. Thorough and frequent cleaning of meat residues and exudate during meat production and handling is critical to reduce STEC biofilm formation even at 13°C.
Introduction
Shiga toxin–producing Escherichia coli (STEC) are important foodborne pathogens associated with large outbreaks, hemolytic uremic syndrome, kidney failure, or even death (Peco-Antić, 2016). STEC outbreaks are frequently associated with beef and beef products. Contamination of beef with STEC may occur during slaughter, dressing, chilling, or cutting (Koutsoumanis and Sofos, 2004). Consequently, STEC populations may be distributed on surfaces of equipment used to produce meat during slaughter and fabrication, contaminating carcasses and fresh meat products (Stromberg et al., 2018; Toro et al., 2018). The presence and survival of STEC on beef fabrication food-contact surfaces and equipment are often mediated through formation of biofilms (Aslam et al., 2004; Wang et al., 2012).
Bacterial biofilms are microbial sessile communities attached to a surface, an interface, or to each other. In biofilms, cells are embedded in a self-produced matrix and protected against environmental stressors such as disinfectants (Nesse et al., 2014). Compared with planktonic cells, members of a biofilm community exhibit altered phenotypes for growth rate and gene transcription (Flemming et al., 2016).
Attachment of bacterial cells to abiotic and biotic surfaces is affected by factors, including the presence of organics, nutrient availability, temperature, production of extracellular polysaccharides, physicochemical properties of both food-contact and cell surfaces, and cell-to-cell communication (Hood and Zottola, 1997; Passos da Silva et al., 2017). Biofilms enhance bacterial survival rates in suboptimal conditions and increase resistance to disinfectants and antimicrobials (Teirlinck et al., 2017). Biofilms formed on food-contact surfaces are a major risk factor for contamination during food processing.
To date, most studies of STEC biofilms have used polystyrene microwell assays, which do not mimic conditions encountered in food-processing environments (Chen et al., 2013; Wang et al., 2016). It is important to ensure that studies allow interpretation and extrapolation of laboratory-based results to the food industry. Various experimental systems have been used to mimic conditions encountered by STEC during meat processing, including cooked or raw meat (Abdul-Raouf et al., 1993; Dourou et al., 2011), packaging conditions (Miya et al., 2014), and adherence to stainless steel surfaces (Rivas et al., 2007).
“Meat juice” has been used as a food-based model, mimicking the nutrient profile of meat processing (Hood and Zottola, 1997; Ferreira and Domingues, 2016). Previous studies demonstrated that beef juice influences growth and gene expression in planktonic E. coli (Hood and Zottola, 1997; Noyce et al., 2006), but there are few reports on impacts of this exudate on biofilm formation by STEC. Brown et al. (2014) identified that chicken juice enhanced cell survival and biofilm formation by Campylobacter. Wang et al. (2013) found that growth of Salmonella within a biofilm was slower with chicken juice compared with tryptic soy broth (TSB). Therefore, meat juice represents a complex nutrient matrix that may promote or inhibit STEC biofilm formation.
In this study, we evaluated effects of beef juice and temperature on biofilm formation by STEC on stainless steel coupons, a common contact surface in food-processing operations.
Materials and Methods
Bacterial strains and cultivation
Based on our previous study (Ma et al., 2019), three strong biofilm-forming STEC strains (EC20020170 O113:H21, EC19990166 O145:H25, and EC20010076 O91:H21) and three weak biofilm-forming STEC strains (EC2011007 O157:H7, EC19940040 O45:H2, and EC20030053 O111:NM) were selected. All strains were streaked onto Luria–Bertani (LB) agar (BD) and incubated at 37°C for 18 h. An isolated colony was then inoculated into 10 mL of minimal salt (M9) broth (Thermo), supplemented with 0.4% glucose, 0.02% MgSO4, and 0.001% CaCl2 (wt/vol), and grown at 37°C on a rocker platform at 180 rpm for 18 h.
Food-contact surface materials and preparation of beef juice
Type-304 stainless steel coupons (2.54 × 7.62 × 0.081 cm; Biosurface) were soaked for 5 min in 10% bleach, thoroughly rinsed with deionized water, air-dried, and autoclaved. Beef juice was prepared as described previously (Wang et al., 2013), with minor modifications. Briefly, a block of fresh beef (i.e., rib eye) was purchased from a local beef processor and frozen at −20°C. The beef was sectioned into 60 g portions and homogenized (WARING, Torrington, CT) with sterile water at a maximum speed for 3 min to generate a 20% (wt/wt) suspension. Large particles were removed by centrifugation (8000 × g, 10 min) and supernatant was sterilized by passage through a 0.22-μm filter (Millipore, MA). Concentration of protein in beef juice was measured using a biuret reagent kit (Sigma-Aldrich, St Louis, MO) and adjusted to 5 mg protein/mL through addition of sterile water as required (Wang et al., 2013).
Biofilm formation in beef juice
A conventional static biofilm formation assay (Thiran et al., 2018) assessed impacts of beef juice with minor modifications. Briefly, overnight bacterial cultures were diluted with M9 medium, M9 medium supplemented with 25% (vol/vol) beef juice, and M9 medium supplemented with 50% (vol/vol) beef juice or 100% beef juice to achieve a final concentration of 107 CFU/mL bacterial cells. Subsequently, 20 mL of the mixture was added into 50 mL Falcon tubes containing a sterile stainless steel coupon. Tubes were incubated at 22°C or 13°C without shaking for 24, 48, or 72 h. Two technical replicates for each strain were conducted in each independent assay. Results were expressed as an average of three independent assays. Coupons incubated with medium only served as negative controls.
Crystal violet staining
Biofilms formed on stainless steel surfaces were assessed using a modification of procedures by Wang et al. (2016). After each incubation period, stainless steel coupons were removed from tubes using sterilized forceps and rinsed in three different tubes containing 25 mL sterilized deionized water, with immersion 40 times in each to remove loosely attached bacteria. Subsequently, coupons with adherent biofilms were fixed with 25 mL absolute methanol (Sigma-Aldrich) for 15 min. Fixed coupons were air-dried for 2 min and stained with 0.5% (w/v) crystal violet solution (Sigma-Aldrich) on a rocker platform at 40 rpm for 15 min.
After staining, coupons were immersed three times in 25 mL sterile water, removing excess crystal violet solution, and air-dried at room temperature for 5 min. Coupons were then immersed in 25 mL of 33% glacial acetic acid (Sigma-Aldrich) and left at room temperature for 15 min. Dissolved dye was measured at 590 nm using a spectrophotometer (Genesys 20; Thermo).
Enumeration of planktonic and attached cells
To count planktonic cells after each period, 1 mL of culture was serially diluted with 10 mM sterile phosphate-buffered saline (PBS, pH 7.4), plated on LB agar, and incubated at 37°C for 18 h. To enumerate biofilm cells, coupons were rinsed three times with sterile deionized water as described above, immersed in 25 mL of sterile PBS, and sonicated at 20 kHz for 10 min (M3800, Branson; Thermo). After sonication, tubes containing coupons were vortexed, and 1 mL of bacterial suspension serially diluted and plated on LB agar. Colonies were counted after incubation for 18 h at 37°C.
Scanning electron microscopy
Scanning electron microscopy (SEM) analyses of biofilm formation of STEC strains on stainless steel surfaces were performed as described previously (Huang et al., 2019). Briefly, coupons with biofilms were rinsed three times in deionized water as described above, fixed with 2.5% glutaraldehyde at 4°C overnight, and twice washed with PBS. Biofilms were dehydrated for 10 min in a graded ethanol series (50%, 70%, 90%, and 100%) and for 15 min in 100% ethanol, followed by a mixture (1:1) of 100% ethanol and tertiary butanol, and finally, absolute tertiary butanol. Specimens were coated with gold and examined using S-4800 SEM (Hitachi, Tokyo).
Statistical analysis
All experiments were conducted three times independently. Data were analyzed using MIXED and least-squares differentiated means with SAS (Statistical Analysis Systems Institute, Cary, NC). Significant differences are presented at a 95% confidence level (p ≤ 0.05).
Results
Growth of the STEC planktonic cells in beef juice
At 22°C, isolates did not differ in growth in M9 media or M9 media supplemented with 25% or 50% beef juice over 72 h (Fig. 1A). Growth of all isolates decreased (p < 0.05) in 100% beef juice at 22°C at 24 h, but populations of all isolates, except for O111, increased (p < 0.05) after 72 h of incubation with beef juice. At 13°C, growth of all strains over 24 h was reduced compared with 22°C, particularly in M9 medium. At 13°C, compared with M9 medium, growth of O113, O91, O157, O111, and O45 was improved by 25% and 50% beef juice at all incubation times, whereas O145 was only enhanced at 24 h. Pure beef juice improved growth of O113 at 48 and 72 h, O91 at 48 h, O111 at 48 and 72 h, and O45 at 24 h when compared with M9 at 13°C.
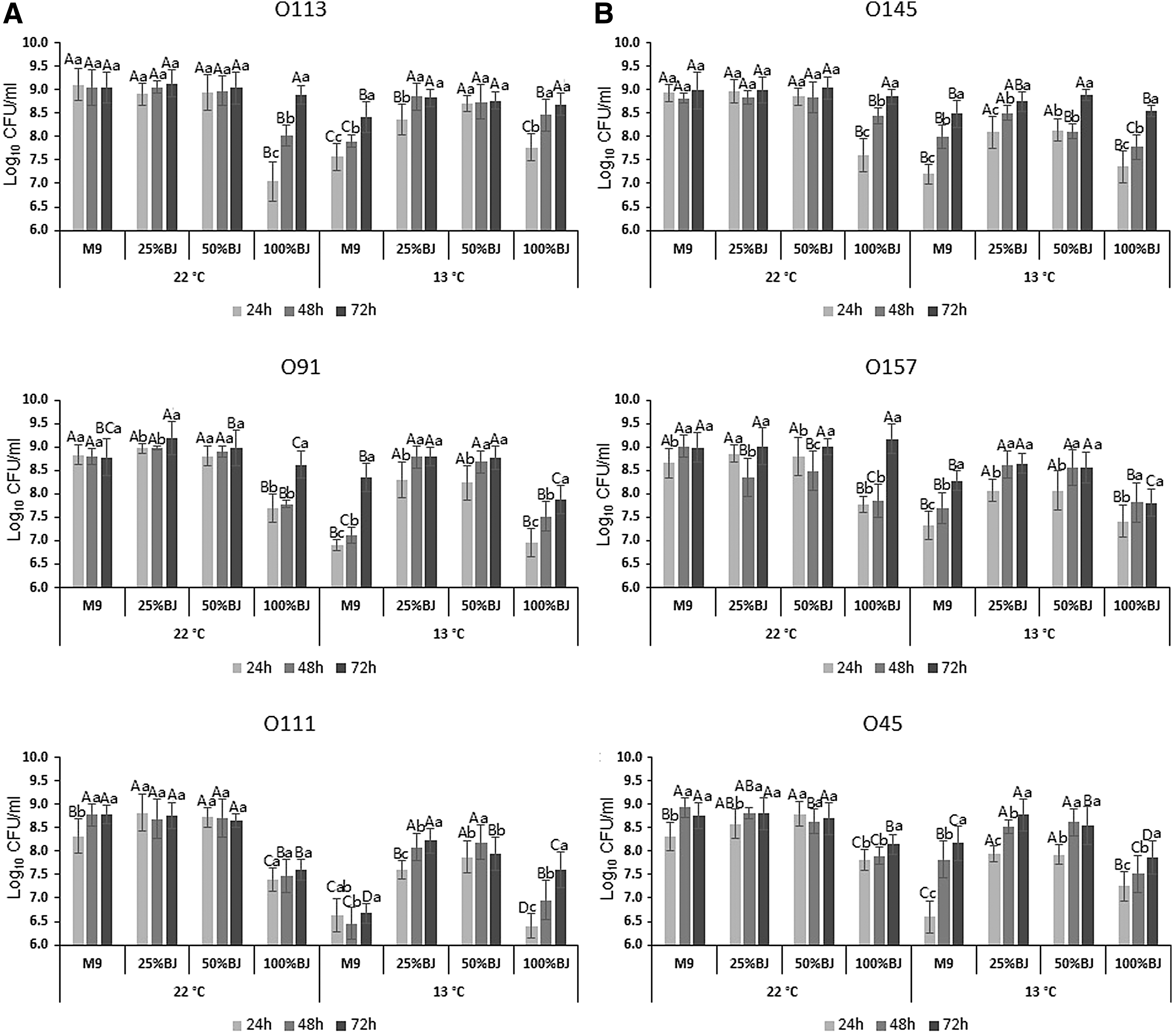
Growth of planktonic cells of the six Shiga toxin–producing Escherichia coli isolates after incubation at 22°C
Beef juice increased biofilm production of some STEC on stainless steel
Based on the optical density (OD)590nm, the cutoff optical density value (ODc) of 0.044 was three standard deviations above the mean OD of negative controls. Strains were classified as non (0.044 > A590), weak (0.088 > A590 > 0.044), intermediate (0.176 > A590 > 0.088), or strong biofilm formers (A590 > 0.176; (Stepanović et al., 2000) (Fig. 2). As shown in Figure 2A, after 24 h, only O113, O145, and O91 exhibited strong biofilm-forming abilities in M9 medium at 22°C, and this activity decreased (p < 0.05) thereafter. The biofilm-forming ability of O113, O145, O91, and O157 was reduced (p < 0.05) in M9 with 25%, 50%, and pure beef juice at 24 h of incubation. However, biofilm formation was enhanced (p < 0.05) with increasing incubation time, except for O157.
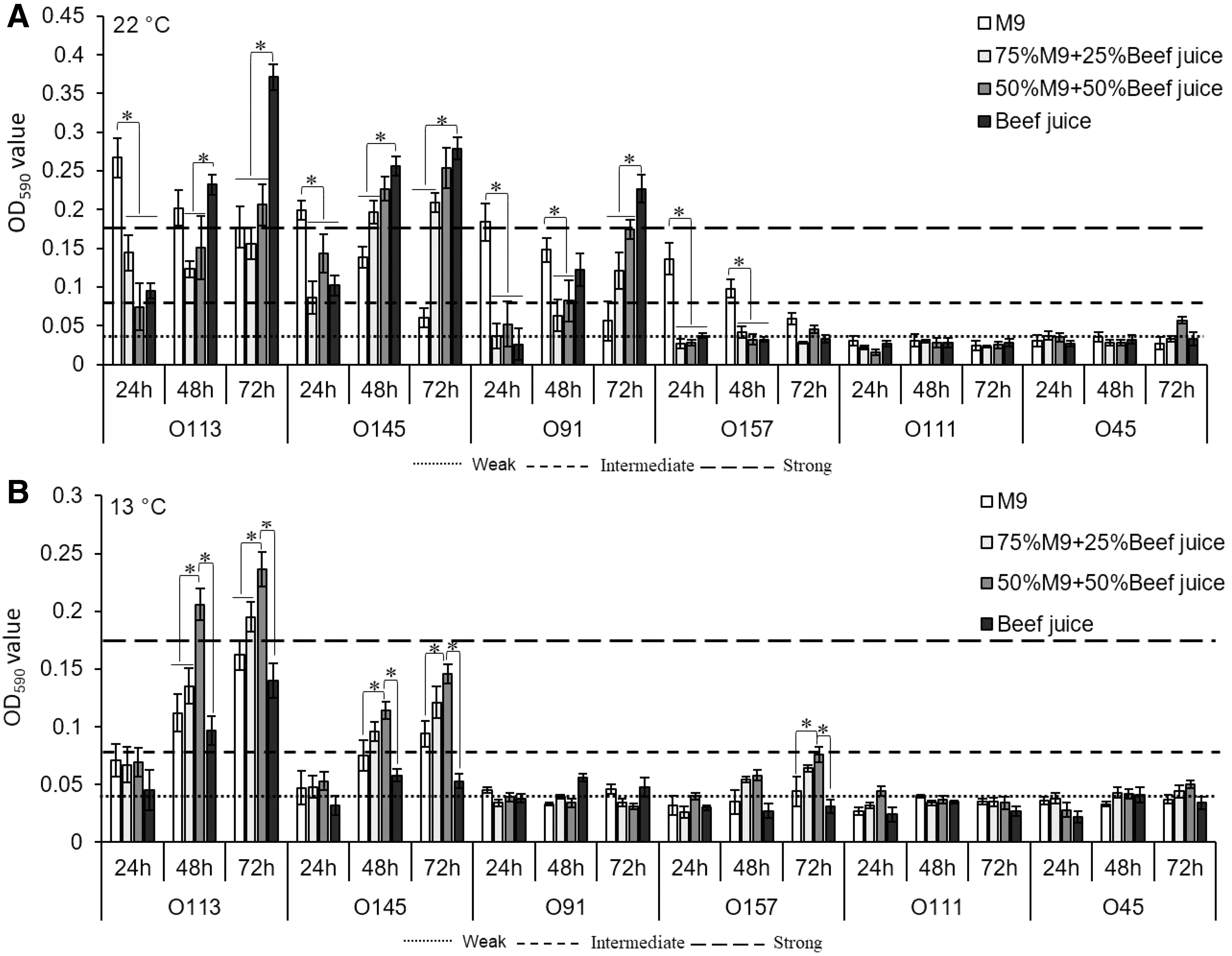
Biofilm formation of six Shiga toxin–producing E. coli strains on stainless steel surface at 22°C
This response was most apparent when O113, O145, and O91 were incubated in pure beef juice. At 13°C, biofilm formation of O113, O145, O91, and O157 was weak at 24 h (Fig. 2B). However, after incubation for 48 and 72 h, biofilm strength of O113 and O145 was enhanced (p < 0.05) in media supplemented with 50% beef juice, compared with M9 medium. Unexpectedly, there was no difference in biofilm formation from cultures in M9 medium and 100% beef juice for all tested strains at 13°C. Both O111 and O45 did not form strong biofilms at either 22°C or 13°C, regardless of media composition.
Enumeration of biofilm cells in beef juice
Biofilm cells associated with O113, O145, O91, and O157 achieved 7.9 to 8.3 log CFU/coupon in M9 medium at 22°C over 24 h (Fig. 3). However, after 72 h of incubation, cell numbers dropped by 0.52 to 1.1 log CFU/coupon in M9 medium. In contrast, the numbers of biofilm cells of O113, O145, and O91 in pure beef juice increased (p < 0.05) by 1.5 to 2.3 log CFU/coupon over 72 h. After 24 h at 13°C, numbers of biofilm cells of all six strains were less than 4 log CFU/mL in all media (Fig. 3). After 72 h of incubation, O113, O145, and O157 had the highest (p < 0.05) numbers of biofilm cells in M9 with 50% beef juice, with 7.5, 6.9, and 6.3 log CFU/coupon, respectively. Noticeably, no biofilm cells were detected for O111 in M9 and 100% beef juice at 13°C (Fig. 3).
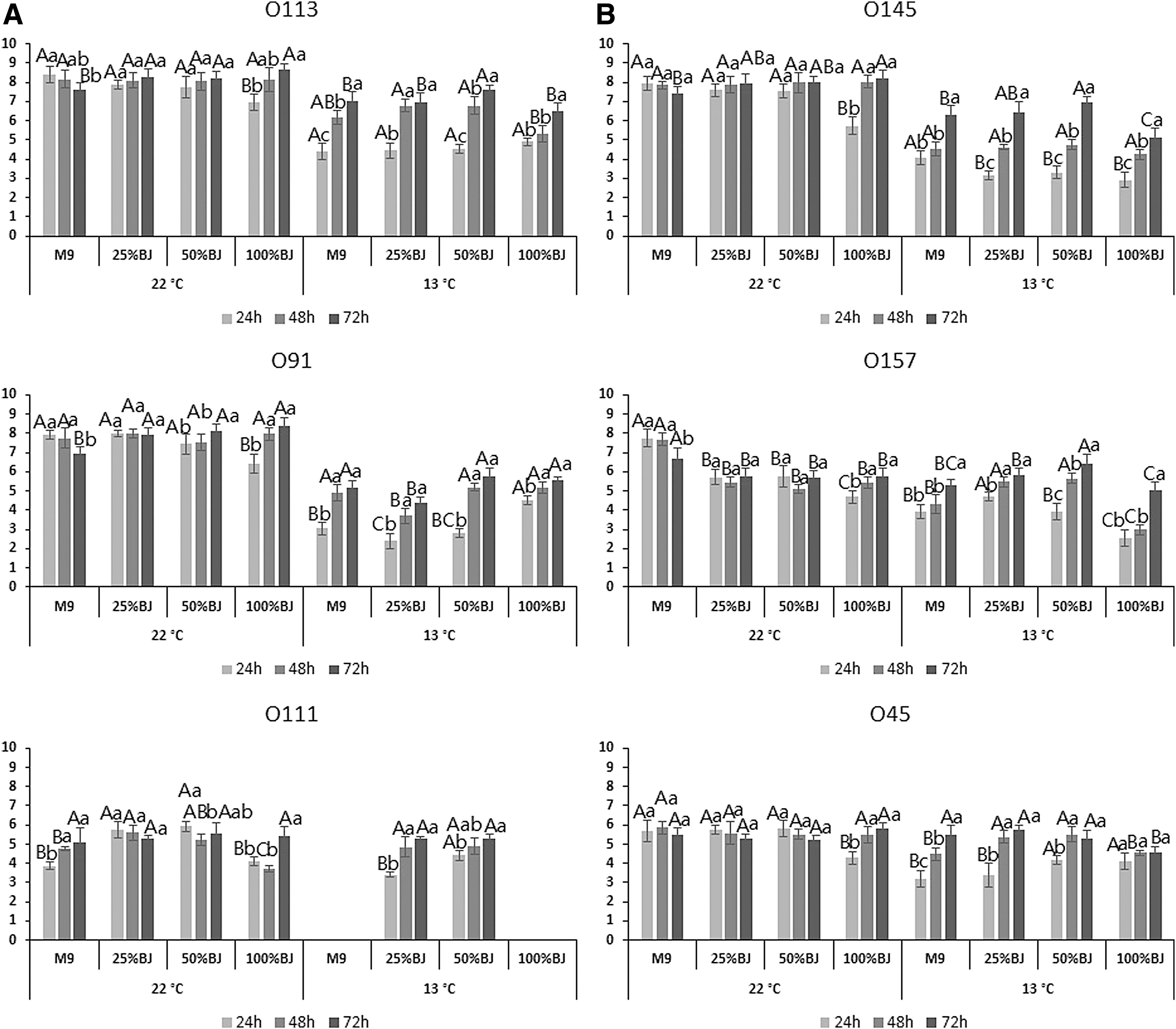
Quantification of biofilm cells of six Shiga toxin–producing E. coli isolates after incubation at 22°C
Scanning electron microscopy
Over 24 h, O113 formed strongest biofilms in M9 medium, with many layers of bacterial cells overlapping on surfaces of stainless steel coupons (Fig. 4). Many biofilm cells also attached to the surface in media containing beef juice (25%, 50%, and 100%), but they remained in monolayers and were sporadic, with few cell aggregates observed. Interestingly, with 100% beef juice, cells were linked by an obvious extracellular matrix, and most cells exhibited an elongated morphology compared with those grown in M9.

Representative scanning electron microscopy images of O113 biofilms cultivated in M9 medium supplemented with 0%, 25%, 50%, or 100% beef juice at 22°C for 24, 48, and 72 h on stainless steel coupons. Arrows indicate extracellular matrix.
After 48 h, biofilm cell numbers were reduced in M9 medium, but cell aggregates were larger in media containing beef juice. A higher concentration of beef juice (i.e., 50% and 100%) contributed to a continuous increase in cell attachment to stainless steel surfaces. After 72 h, a strong biofilm was observed in 100% beef juice, with many cell aggregates linked by an extensive extracellular matrix (arrows in Fig. 4). A similar pattern of biofilm formation was observed for O145 over the same incubation period, although the biofilm-forming ability of O145 appeared weaker than that of O113 (Fig. 5).

Representative scanning electron microscopy images of O145 biofilms incubated in M9 medium supplemented with 0%, 25%, 50%, or 100% beef juice at 22°C for 24, 48, and 72 h on stainless steel coupons.
Discussion
In this study, beef juice was a food-based model mimicking meat-processing industry conditions, which was validated for investigating filter-sterilized beef juice effects on STEC growth and biofilm formation. All STEC strains in the present study were capable of growing in beef juice, indicating that beef juice may serve as a nutrient source and promote growth of these pathogens in beef-processing environments.
Planktonic growth of all six strains did not differ between cells grown in M9 medium alone or mixed with 25% or 50% beef juice at 22°C. In contrast, STEC strains grew slower in pure beef juice than in M9 medium for the first 24 h at 22°C. This was consistent with a previous report, in which E. coli O157:H7 and Salmonella Typhimurium had lower numbers of viable planktonic cells in beef juice than in TSB after 24 h of incubation at 23°C (Hood and Zottola, 1997).
There are several possible reasons for this. Standard laboratory media, such as TSB, LB, and M9, are considered balanced-nutrient media, while diluted beef juice is generally considered low-nutrient media. Accordingly, the log phase of bacterial growth will be prolonged in low-nutrient media (Hood and Zottola, 1997; Li et al., 2017). Also, slower growth could, in part, be due to the presence of natural inhibitors (serum proteins and antimicrobial peptides) or missing essential nutrients within beef juice (Bugla-Płoskońska et al., 2009; Lee et al., 2011).
Previous reports have demonstrated that STEC form stronger biofilms at 22°C than at 37°C (Wang et al., 2016). Low temperatures (5–15°C) are generally maintained in meat-processing environments (Nesse et al., 2014). Therefore, two temperatures (22°C and 13°C) relevant for food production and handling were used to investigate the biofilm-forming abilities of STEC isolates in beef juice. As shown in Figure 2A, the biofilm-forming capacity of O113, O145, and O91 in M9 medium was inversely related to that formed in pure beef juice at all time points after 24 h at 22°C. In M9 medium, strains, including O113, O145, and O91, formed strong biofilms after 24 h, but biofilm cell numbers declined over time.
It is not clear why the numbers of biofilm cells did not always increase over the entire incubation period. One possible explanation is that bacterial cells were dispersed from formed biofilms. Previous literature has reported that dispersal of cells from the biofilm is an essential stage of the biofilm life cycle (Barraud et al., 2015). Conversely, biofilm formation in 100% beef juice for O113, O145, and O91 increased over time. Differences in biofilms formed in M9 and pure beef juice may be attributed to the growth phase of the bacteria. Previous studies have demonstrated that, for cells in exponential or stationary phase, as numbers of cells in suspending medium increased, the numbers of biofilm cells initially increased, plateaued, and declined thereafter (Fletcher, 1977; Hood and Zottola, 1997).
Under conditions of the present study, isolates capable of forming biofilms reached stationary phase in M9 medium within 24 h. However, isolates growing in beef juice were still in log growth phase for at least 72 h (Fig. 1A), so biofilm formation continued to increase during this period. Also, as M9 is a minimal medium, a lack of nutrients and metabolic waste accumulation in M9 after 24 h of incubation may decrease biofilm surface populations. Furthermore, SEM images indicated that the morphology of biofilms formed in beef juice differed from those formed in M9. In beef juice, biofilm cells were covered by an extensive extracellular matrix, which may slow release of cells from biofilms.
Density of cells in biofilms formed in beef juice at 72 h was greater than that in M9 medium at 24 h. Researchers have speculated that low-nutrient systems may enhance biofilm formation (Brown et al., 1977; Allan et al., 2002). This increased biofilm formation during nutrient-deprived conditions may be related to changes in cell morphology and surface characteristics, which help cells become more closely associated with a surface, an adaptation enhancing nutrient trapping and scavenging (Kjelleberg and Hermansson, 1984; Wrangstadh et al., 1986). In our SEM images, changes in cell morphology and more extracellular matrix production were observed for both O113 and O145 in beef juice, which are consistent with this possible response.
Another explanation for enhanced biofilms is that beef juice promotes formation of a conditioning layer composed of carbohydrates, proteins, lipids, and minerals on abiotic surfaces. This conditioning layer may provide an adhesive foundation that stimulates biofilm establishment and growth (Brown et al., 2014). In comparison, we also supplemented M9 medium with 25%, 50%, and 100% sterilized water, and tested effects of diluted M9 on the biofilm formation of STEC isolates. Results demonstrated that the biofilm levels of O113, O145, and O91 did not increase over time as a result of the addition of water to M9 medium (Supplementary Fig. S1), suggesting that enhanced biofilm formation was not due to a dilution of nutrients in M9, but to the presence of beef juice.
At 13°C, 25% and 50% beef juice enhanced biofilm formation compared with M9 medium. However, pure beef juice did not improve biofilm formation. Similar results were reported by Li et al. (2017), who studied effects of 100% chicken juice on biofilm formation in Salmonella, which reduced numbers of attached cells compared with 50% chicken juice. One possible explanation for this is that levels of meat juice exceeding 50% promote formation of a precoated single layer on contact surfaces leading to detachment at lower temperatures (Brown et al., 2014; Li et al., 2017). Hence, biofilms may detach along with this loosely structured double layer during washing.
Addition of beef juice to M9 had no impact on biofilm formation of O111 and O45 at either 22°C or 13°C. Based on our previous studies (Wang et al., 2016; Ma et al., 2019), O111 and O45 formed strong biofilms on polystyrene in M9 and LB broth. However, they could not form biofilms on stainless steel in M9 or LB media. Therefore, it can be inferred that properties of the attachment surface can influence the affinity of these isolates regardless of the type of media used. Furthermore, previous literature reported that bacterial surface characteristics such as hydrophobicity, fimbriae, and flagella play an important role in biofilm formation (Houry et al., 2010; Nesse et al., 2014).
Although the morphology of O113 and O145 changed with the addition of beef juice, surface properties or metabolic mechanisms of O111 and O45 may not be altered by the presence of beef juice. Therefore, no difference in biofilms formed in beef juice and M9 was observed for low-biofilm producers.
In conclusion, results of this study confirm that beef juice promotes STEC biofilm formation on stainless steel surfaces, although impacts are strain-, temperature-, and concentration-dependent. Findings in the present study confirm that STEC isolates can survive in beef exudates as attached and planktonic cells. Formation of STEC biofilms was reduced at lower temperatures, so our results support the important role of cold-chain management in mitigation of STEC. Additional studies could attempt to define the specific beef juice components (e.g., types of proteins and quorum-sensing molecules) that alter bacterial biofilm formation. Such work could lead to identification of those specific components promoting biofilm growth and strategies to reduce biofilm formation.
Furthermore, we only investigated monospecies biofilms in the present study. Mixed-species biofilm formation by STEC and Salmonella in meat-processing environments has occurred, which would provide additional information as to how nutrient exudates influence natural biofilm formation within meat-processing facilities.
Footnotes
Acknowledgments
We acknowledge the technical assistance of R. Ha, C. Conrad, S. Trapp, and Y. Graham during the study as well as Dr. Roger Johnson (PHAC) for kind provision of STEC strains.
Disclosure Statement
No competing financial interests exist.
Funding Information
Financial support was provided by Agriculture and Agri-Food Canada, Alberta Agriculture and Forestry, China Scholarship Council, and the Beef Cattle Research Council Beef Cluster funding program.
Supplementary Material
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.