
Abstract
Shiga-toxigenic Escherichia coli (STEC) and enterotoxigenic E. coli (ETEC) can cause diarrhea in piglets. This is the first report and complete genome sequence of an extended-spectrum β-lactamase-producing hybrid STEC/ETEC strain isolated from a piglet with diarrhea on a swine farm in China. We investigated the virulence genes and phylogenetic diversity with publicly available E. coli genomes. Both E. coli strains S17-13 and S17-20 harbored multiple virulence genes, mainly including stx2e and eastA genes. Other important virulence genes (estIa, estIb, fedABCDEF, and hlyABCD) were located in the plasmid p1713-1 of S17-13, which could be transferred from E. coli S17-13 to S17-20 by conjugation. The presence of virulence genes associated with different pathogroups (STEC or ETEC) confirmed the hybrid status of E. coli strain S17-13. Phylogenetic analysis showed that STEC/ETEC S17-13, STEC S17-20, avian pathogenic E. coli (APEC) O78, and APEC ACN001 are located in the same evolutionary branch, indicating that they may originate from a common ancestor. It is crucial to understand the phylogeny of pathogenic bacteria to evaluate how they have evolved and to monitor the emergence of potential new pathogens. The emergence of novel hybrid E. coli strains presents a new public health risk. More attention must be paid to these hybrid pathogens during typing and epidemiological surveillance of E. coli infections, which challenges the traditional diagnostics of E. coli infections.
Introduction
E
Pathogenic E. coli are common agents responsible for a variety of intestinal disorders, such as diarrhea, edema disease syndrome, and even death in pigs (Kim et al., 2010). Diarrheal E. coli (DEC) cause constant problems at large-scale farms, particularly among piglets weaned at 3–4 weeks of age. Traditionally, DEC were classified into six pathogroups according to virulence factors and pathogenic characteristics, including enterotoxigenic E. coli (ETEC), enterohemorrhagic E. coli (EHEC) or Shiga-toxigenic E. coli (STEC), enteroaggregative E. coli (EAEC), enteropathogenic E. coli (EPEC), enteroinvasive E. coli (EIEC), and diffusely adherent E. coli. STEC and ETEC are two important pathogroups of DEC.
STEC can cause bloody or nonbloody diarrhea. The common features of STEC strains are the production of Shiga toxin 1 (Stx1), Shiga toxin 2 (Stx2), a variant of Stx2 (Stx2c, Stx2d, Stx2e, Stx2f, Stx2g), or a combination of these toxins (Kim et al., 2010). Stx2e is typically produced by STEC strains that cause edema disease in swine. ETEC causes watery diarrhea due to the production of heat-labile (LT) and/or heat-stable (ST) enterotoxin. F18 fimbriae is one of the main adhesive factors of ETEC.
E. coli are able to acquire virulence genes through horizontal gene transfer from other pathogroups leading to the development of intermediate or hybrid pathogroups (Mellmann et al., 2011). A hybrid of EAEC/STEC O104:H4 caused a large outbreak of severe disease with mortalities in Germany in 2011 (Mellmann et al., 2011). Hybrids of STEC/ETEC have recently been reported in Korea (Kim et al., 2010), Germany, the United States, and Finland (Nyholm et al., 2015a, 2015b), but have not, to date, been reported in China.
We isolated a new extended-spectrum β-lactamase (ESBL) producing STEC/ETEC hybrid strain S17-13, and a STEC strain S17-20 from a piglet with diarrheal disease in Jilin province of China. In this study, we investigated these two porcine strains to determine their phylogenetic position among E. coli, and to identify the similarities and differences of gene contents and virulence properties between them and other pathogroups. It is very important to understand the phylogeny of pathogenic bacteria for evaluating how they have evolved and to monitor the emergence of potential new pathogens.
Materials and Methods
Bacterial strains
The E. coli strains S17-13 and S17-20 were isolated from rectum fecal samples collected from piglets with diarrhea on a swine farm in Jilin province of China in July 2017. Typical clinical symptoms of the disease included severe diarrhea and weight loss. To identify the pathogen, 12 rectum fecal samples were collected from diarrhea of piglets. S17-13 and S17-20 were isolated from these samples, but no other pathogen was isolated or detected. These two strains were inoculated onto MacConkey agar and identified using the Phoenix-100 system (BD, MD).
According to a previously reported method (Kim et al., 2010), polymerase chain reaction was used to identify the virulence genes of these E. coli isolates. These results showed that E. coli strain S17-13 possessed the pathogenic features of both STEC and ETEC, and could be classified as a hybrid STEC/ETEC strain. S17-13 was positive for virulence genes stx2e, eastA, estIb, estIa, and the F18 fimbriae fed gene. E. coli strain S17-20 was positive for virulence genes stx2e and eastA. These two strains S17-13 and S17-20 were selected for whole-genome sequencing.
Antibiotic susceptibility testing
Antibiotic susceptibility of strains S17-13 and S17-20 was tested and analyzed by using BD Phoenix-100 Automated Microbiology System (Panel NMIC/ID-4; BD Diagnosis Systems). The phenotypes of extended-spectrum β-lactamases (ESBLs) were also recognized by the system. Minimum inhibitory concentrations (MICs) were based on broth microdilution susceptibility method following the standards of the Clinical Laboratory Standards Institute (Document No. M100-28th Edition, 2018).
DNA extraction and whole-genome sequencing
Genomic DNA was extracted using a Qiagen Genomic Tip 100/G (Qiagen, Gaithersburg, MD) according to the manufacturer's instructions. After the extraction, the intactness of the genomic DNA was verified by agarose gel electrophoresis, and the quantity of the genomic DNA was measured using a Qubit 2.0 Fluorometer (Invitrogen, Carlsbad, CA). Sequencing libraries were constructed using SMRTbell Template Prep Kit 1.0-SPv3 (Pacific Biosciences, CA) according to the manufacturer's protocol. Sequencing was performed on a PacBio Sequencer (Pacific Biosciences) by Realbio Technology (Shanghai, China).
De novo genome assembly and genome annotation
The data collected from the PacBio sequencer were processed and filtered using the single molecule real-time (SMRT) analysis software suite (Pacific Biosciences). Data were filtered according to read quality (>0.75) and read length (>500 bp). When processing continuous long read (CLR) data, raw reads from the SMRT cells were split on adapter sequence resulting in ≥1 subread or CLR per zero-mode wave guide. A total of 349,788 and 311,478 reads were achieved for E. coli S17-13 and S17-20, respectively. Assembled genomes were submitted to the GenBank database for genome annotation.
Extraction of virulence genes and ETEC colonization factors
The genomes were screened for a broad spectrum of known and characterized bacterial virulence and colonization factor genes using the virulence factor database (VFDB) (Chen et al., 2005). The target loci were imported from publicly available annotated sources (e.g., genomes, plasmids, coding sequences). Required thresholds for gene identification were ≥80% identity to a reference sequence and ≥99% alignment with a reference sequence.
Identification of prophage regions
The prophage regions in the STEC/ETEC genomes were identified using the PHAST tool (
Phylogenetic analysis and whole-genome alignment
To generate a phylogenetic tree depicting the positions of the STEC/ETEC strains, the genomes were compared with 38 published E. coli genomes. The core genome phylogeny was inferred from the shared genes among a diverse set of E. coli genome sequences using cgMLST (Nyholm et al., 2015a) and sequence alignment. Whole-genome alignment was performed using Mauve (version. 2.4.0) (Darling et al., 2010) to examine the alignment and distribution in the genomes.
Nucleotide sequence accession numbers
The genome nucleotide sequences of strains S17-13 and S17-20 were deposited into the GenBank nucleotide sequence database under accession numbers CP024997 and CP025036, respectively. The nucleotide sequences of the plasmids were deposited into the GenBank database under accession numbers CP031766 and CP031767 for strain S17-13, and CP031764 and CP031765 for strain S17-20.
Results
Antimicrobial susceptibility of E. coli S17-13 and S17-20
The S17-13 strain was resistant to six antimicrobials, including ampicillin, piperacillin, gentamicin, ciprofloxacin, levofloxacin, and tetracycline. As shown in Table 1, S17-13 showed the phenotype of ESBL, and high resistance to ampicillin (MIC >1024 mg/L) and piperacillin (MIC >256 mg/L). Both of the S17-13 and S17-20 strains showed higher tolerance to gentamicin (MIC >1024 mg/L), ciprofloxacin (MIC >32 mg/L), levofloxacin (MIC >32 mg/L), and tetracycline (MIC >256 mg/L).
Antimicrobial Susceptibility of Escherichia coli S17-13 and S17-20
Data obtained by broth microdilution susceptibility method.
ESBL, extended-spectrum β-lactamase; I, intermediate; MIC, minimum inhibitory concentration; R, resistant; S, susceptible.
General genome features of E. coli S17-13 and S17-20
The genome sequences of two E. coli strains isolated from fecal samples of piglets with diarrhea in the same swine farm were determined. The genome size of both isolates was ∼5.2 Mbp. A total of 5012 (S17-13) and 5045 (S17-20) protein-encoding genes were predicted. The general genomic features of E. coli S17-13 and S17-20 are presented in Table 2.
Assembly Data of the Escherichia coli S17-13 and S17-20 Genome Sequences
CDS, coding sequence; CRISPR, clustered regularly interspaced short palindromic repeats; GC, guanine-cytosine content.
Virulence genes and colonization factor genes analysis
The two genomes were screened using the VFDB. Both S17-13 and S17-20 harbored multiple virulence genes (Supplementary Table S1), including the major virulence genes stx2e encoding STEC variant Shiga toxin 2e and eastA encoding EAEC heat-stable enterotoxin I. Other virulence genes were also found, including espL1, L4, R1, X1, X4, and X5 encoding type III secretion system effectors homologous with EHEC O157:H7, the ecpABCDEF genes encoding E. coli common pilus homologous with EHEC O157:H7, the entABCDEFS genes encoding ferri enterobactin-related genes homologous with uropathogenic E. coli (UPEC) CFT073, the fim genes encoding type 1 fimbriae homologous with UPEC CFT073, the gsp genes encoding general secretion pathway proteins homologous with Shigella dysenteriae Sd197 (serotype 1), and the fli and flg genes encoding flagellar homologous with Yersinia enterocolitica subsp. enterocolitica 8081 (biotype 1B).
In addition, some virulence genes were located on the plasmid of strain S17-13 and were absent in strain S17-20, including estIa and estIb encoding ETEC heat-stable enterotoxins a and b (STa and STb), major colonization factor genes fedABCDEF encoding ETEC colonization factor F18 fimbriae, and the hlyABCD genes encoding hemolysin homologous with ETEC UMNK88.
Identified prophage regions and stx-phage
In both strains S17-13 and S17-20, seven prophage regions were identified, of which five regions were intact. For strain S17-13, one region was incomplete, and one region was questionable. For strain S17-20, two regions were questionable. The characteristics of the prophage regions are listed in Table 3. The five intact prophage regions were in the genomes of both strains S17-13 and S17-20. The gene encoding Stx2e was located in an intact prophage of ∼87.2 kb in length. The stx2e-phages in strains S17-13 and S17-20 were similar to bacteriophage P27 (GenBank accession no. AJ298298), as determined by a BLAST search.
The Characteristics of the Prophage Regions in Strains S17-13 and S17-20
The prophage carrying the gene encoding Shiga toxin 2e.
Phylogenetic analysis and whole-genome map comparison
The analysis showed that strains STEC/ETEC S17-13, STEC S17-20, avian pathogenic E. coli (APEC) O78 (Mangiamele et al., 2013; Huja et al., 2015), and APEC ACN001 (Wang et al., 2016) were located within the same evolutionary branch (Fig. 1), indicating that these strains may have originated from a common ancestor. The genome sequences of S17-13 and S17-20 were compared with three APEC genome sequences using Mauve (version. 2.4.0). The results showed that the distribution of genetic material in the genomes of these strains was highly similar (Fig. 2).
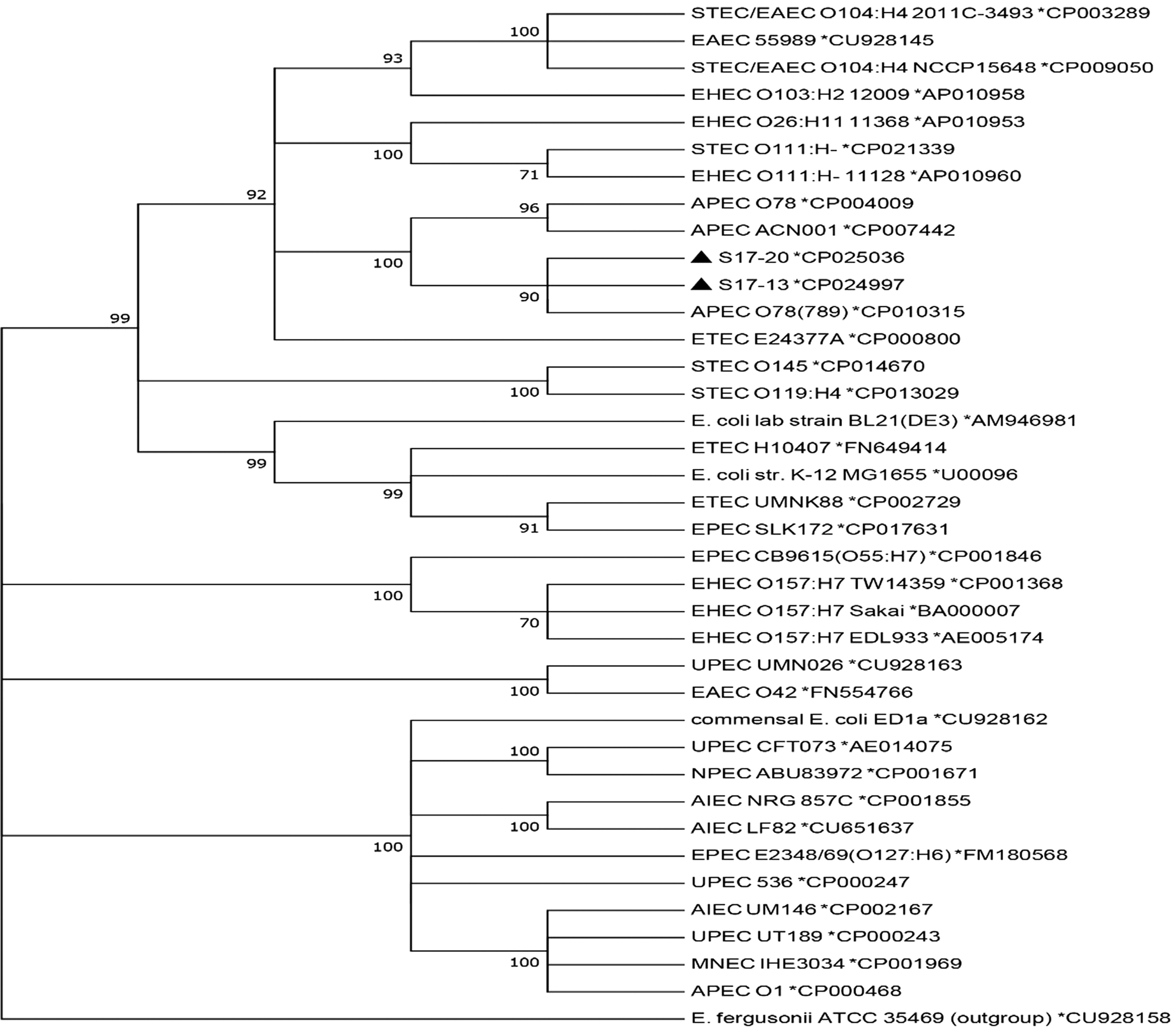
Phylogenetic analysis of completely sequenced Escherichia coli strains. The pathotype of each E. coli strain is indicated in front of the strain name. The number after the * is the GenBank accession number of the genome sequence of each strain. Filled triangles indicate two strains isolated and sequenced in this study. AIEC, adherent-invasive E. coli; APEC, avian pathogenic E. coli; EAEC, enteroaggregative E. coli; EHEC, enterohemorrhagic E. coli; EIEC, enteroinvasive E. coli; EPEC, enteropathogenic E. coli; ETEC, enterotoxigenic E. coli; MNEC, meningitis-associated E. coli; NPEC, nonpathogenic E. coli; STEC, Shiga-toxigenic E. coli; UPEC, uropathogenic E. coli.
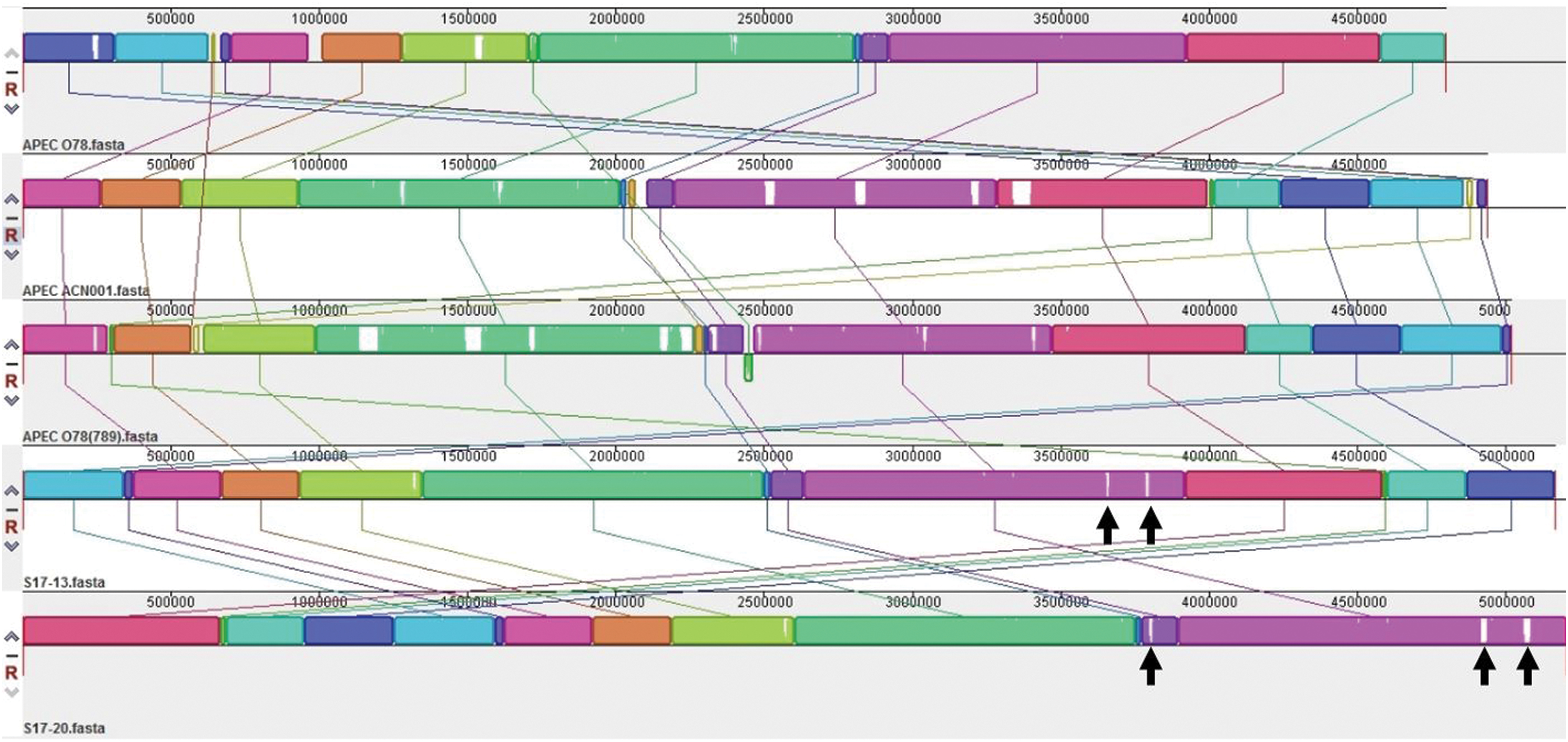
Whole-genome map comparison of S17-13 and S17-20 strains with three APEC strains. The areas in color are common among the compared genome sequences. White areas are unique. Arrows indicate the major differences between the genomes of S17-13 and S17-20. APEC, avian pathogenic E. coli. Color images are available online.
Discussion
The genomics and phylogeny of hybrid E. coli strains have not been studied widely. This study mainly investigated the genome of a swine-derived STEC/ETEC hybrid strain and its phylogenetic position in relation to other E. coli genomes. The information obtained from this study contributes significantly to our understanding of the genomic potential, virulence factors, pathogenicity, and evolution of a hybrid pathogenic E. coli strain.
The varying combination of multiple virulence genes harbored by the hybrid STEC/ETEC strain S17-13 showed that this strain contained a mixture of several different E. coli pathogroup-associated characteristics. The presence of virulence genes stx2e, estIa, estIb, eastA, and the F18 fimbriae genes, fedABCDEF associated with different pathogroups, confirmed the hybrid status in E. coli S17-13. Although S17-13 was positive for astA encoding EAEC heat-stable enterotoxin I, unlike the toxin name indicates, astA can be harbored by strains of STEC, EAEC, EPEC, ETEC, EIEC pathogroups (Paiva de Sousa and Dubreuil, 2001) and even extraintestinal pathogenic E. coli (Toval et al., 2014). In addition, strain S17-13 harbored other virulence genes, some of which have been associated with ETEC, EHEC, and UPEC (Supplementary Table S1).
E. coli virulence factors such as enterotoxins, invasion factors, adhesion factors, and Shiga toxins can be encoded by several mobile genetic elements, which mainly include plasmids, bacteriophages, transposons, or pathogenicity islands. Bacterial plasmids are able to transfer the drug resistance and virulence traits between different strains, and thus play an important role in horizontal gene exchange. We have performed the conjugation experiments to confirm horizontal transfer of plasmid. However, due to the wild-type virulence plasmids do not carry selective markers. It is difficult to screen out conjugon after conjugation experiments.
The Shiga toxin gene stx is located in an intact prophage region, which means that the Stx-phages and their ability to transfer genes horizontally may play an important role in the evolution of E. coli and the development of STEC variants (Schmidt, 2001). According to phylogenetic analysis, strains STEC/ETEC S17-13, STEC S17-20, APEC O78 (Mangiamele et al., 2013; Huja et al., 2015), and APEC ACN001 (Wang et al., 2016) are located in the same evolutionary branch, and these four strains might originate from a common ancestor. We can make a hypothesis about the evolutionary pattern of hybrid STEC/ETEC strain (Fig. 3). First, an ancestral E. coli acquired the virulence genes of APEC and evolved into an APEC strain; meanwhile, the other ancestral E. coli acquired the stx2e gene by horizontal transfer of a prophage and evolved into STEC. Second, STEC S17-20 may acquire the estIa, eatIb, and fedABCDEF genes by the horizontal transfer of plasmid and evolve into STEC/ETEC S17-13.
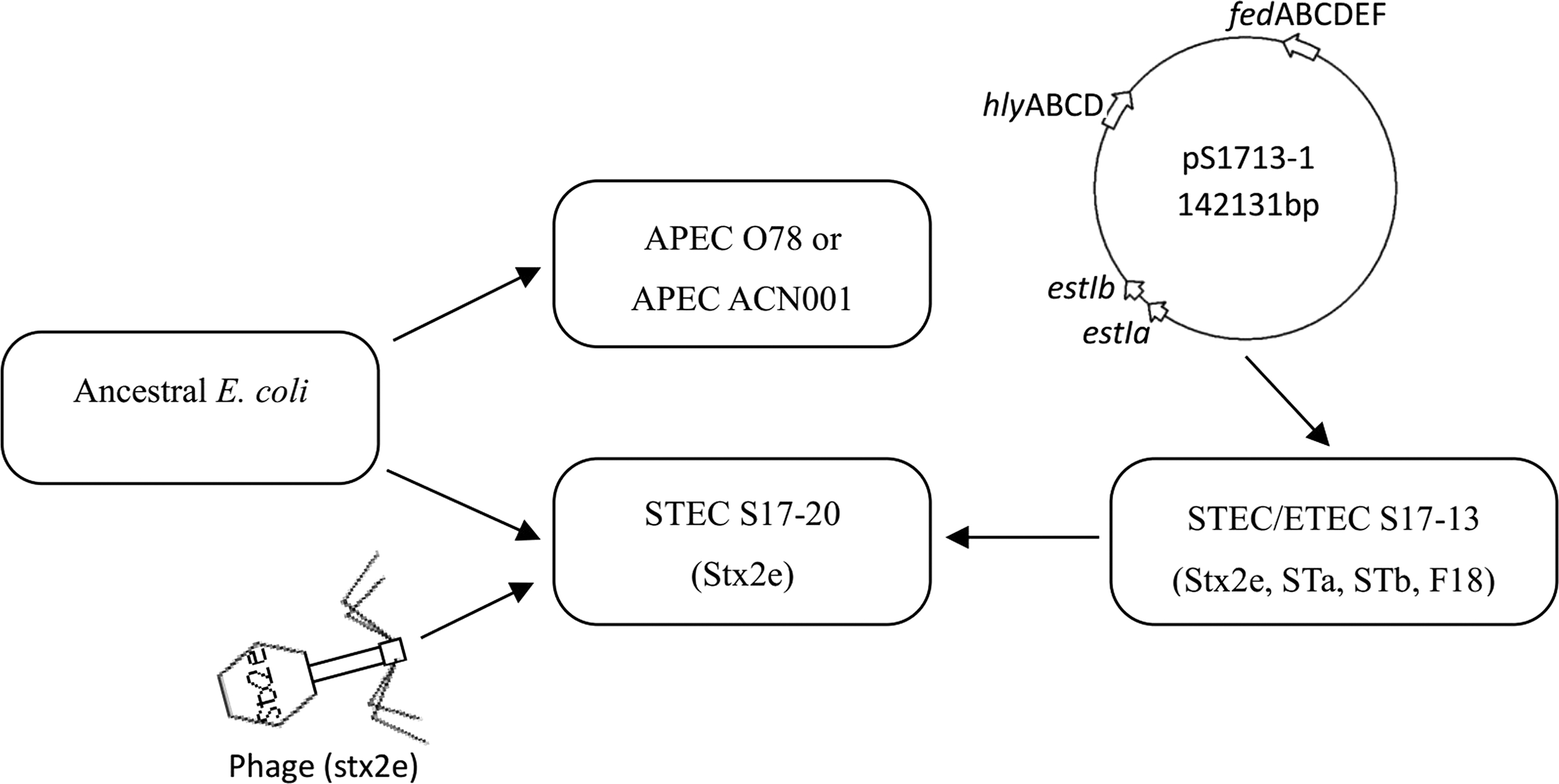
Proposed scheme of the evolutionary pattern of hybrid STEC/ETEC. APEC, avian pathogenic E. coli; ETEC, enterotoxigenic E. coli; STEC, Shiga-toxigenic E. coli.
Conclusions
This is the first report about a new hybrid STEC/ETEC strain isolated from piglets with diarrhea in China. We investigated the virulence gene contents of two E. coli strains and their phylogenetic positions in relation to other E. coli genomes. The stx2e-prophage and plasmid carrying both ST toxin and colonization factor genes may play an important role in the evolution of the STEC strain and the STEC/ETEC hybrid strain. The information obtained from this study contributes significantly to our understanding of the evolutionary pattern of pathogenic E. coli.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Key Research and Development Program of China (grant number 2017YFD0500104).
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.