
Abstract
Due to the increasing number of studies reporting the detection of antimicrobial-resistant isolates of Listeria monocytogenes, we sought to determine the antimicrobial susceptibility of L. monocytogenes isolates collected in Italy and find potential correlations to their serotypes and multilocus sequence types (MLST). The antimicrobial susceptibility of 317 L. monocytogenes isolates collected from food, humans, and the environment from 1998 to 2009 was assessed by minimum inhibitory concentration (MIC). Serotyping and MLST was also performed on all isolates. Potential correlations among antimicrobial resistance profiles, serotyping, and MLST were statistically evaluated. Twenty-four percent of L. monocytogenes isolates were resistant to oxacillin, 28.7% intermediate to clindamycin, and 24.3% to ciprofloxacin. The majority of isolates with elevated MIC to oxacillin was of environmental origin and belonged to serotype 4b/4e and ST2. Isolates with intermediate MIC values to clindamycin and ciprofloxacin were mostly of food and human origin and belonged to serotype 4b/4e and ST9. Regarding the time frame of isolate collection, comparing the last 3 years (2007–2009) to previous years (1998–2006), an increase was observed in the percentage of resistant and intermediate isolates per year. This trend strongly suggests the need for increasing attention on the prevalence of antimicrobial resistance in L. monocytogenes in Italy. To predict future resistance trends, the monitoring of clinical intermediate resistance might represent a useful tool especially for antibiotics associated to multiple-step mechanisms of acquired resistance. A specific focus should be addressed to antimicrobial-resistant isolates of serotype 4b, repeatedly associated with food-borne outbreaks.
Introduction
In the past decades, Listeria monocytogenes has been implicated in several outbreaks of food-borne illness in humans with a statistically significant increasing trend of listeriosis since 2008 (Schlech et al., 1983; EFSA and ECDC, 2015). While it has been established that food-borne transmission constitutes the main acquisition route of listeriosis (Farber and Peterkin, 1991; Swaminathan and Gerner-Smidt, 2007), most healthy humans are not significantly affected by the intake of small numbers of L. monocytogenes in foods (Pouillot et al., 2015). However, the risk of developing clinical listeriosis is higher for people with existing chronic illness, suppression of the immune system, pregnancy, or extreme youth or older age (younger than the age of 1 year or older than 60 years) (Swaminathan and Gerner-Smidt, 2007). For these vulnerable persons, the mortality rate remains high (∼16%) (Scallan et al., 2011). Patients affected by listeriosis are usually treated with aminopenicillin or benzylpenicillin in conjunction with an aminoglycoside. Vulnerable and immunocompromised patients at risk of listeriosis might follow an empiric treatment with broad spectrum antibiotics such as piperacillin/tazobactam or carbapenems (Thønnings et al., 2016).
Listeria spp. has been generally reported susceptible to almost all antimicrobials, except for cephalosporins, fosfomycin, and macrolides, against which Listeria spp. has been described as naturally resistant or with reduced susceptibility (Charpentier and Courvalin, 1999; Troxler et al., 2000). However, reports of L. monocytogenes resistant to a wide range of antimicrobial agents have been published. Since the first report of a multiresistant clinical isolate in France in 1988, antimicrobial-resistant isolates collected from humans, food, plants, and the environment have been described worldwide (Charpentier et al., 1995; Srinivansan et al., 2005; Granier et al., 2011; Ruiz-Bolivar et al., 2011; Bertsch et al., 2014; Moreno et al., 2014; de Vasconcelos Byrne et al., 2016; and Li et al., 2016). In humans, resistance of L. monocytogenes against tetracycline and ciprofloxacin has been described (Morvan et al., 2010). In food, high prevalence of oxacillin and clindamycin resistance was described recently among isolates from meat and fish production chains (Walsh et al., 2001; Lyon et al., 2008; Chen et al., 2010; Gómez et al., 2014). Similarly, significant percentages of resistance against ampicillin, penicillin G, and tetracycline were reported in L. monocytogenes isolated from meat, fish, and dairy production chains (Aras and Ardıç, 2015; Jamali et al., 2015; Kevenk and Gulel, 2016).
The emergence and spread of antimicrobial-resistant Listeria spp. has been attributed to the overuse of antibiotics in disease treatments and growth promotion in domestic livestock (Charpentier et al., 1995; Walsh et al., 2001).
In the present study, the antimicrobial susceptibility of 317 L. monocytogenes isolates collected in a 11-year period (1998–2009) from humans, food, and environmental surfaces was assessed along with serotype and multilocus sequence type (MLST) to: (1) evaluate the occurrence of antimicrobial resistance and intermediate resistance in L. monocytogenes over time; (2) correlate antimicrobial resistance phenotype with origin, serotype, and sequence type.
Materials and Methods
L. monocytogenes isolates
A total of 317 L. monocytogenes isolates recovered between 1998 and 2009 from food (n = 239, including 162 meat products, 38 dairy products, 17 seafood, 22 ready-to-eat products), humans (n = 43), and environment (n = 35, including 17 food-processing environment, 6 animal feces, 4 soil, 4 water, and 4 plants) were analyzed in this study.
All the isolates were previously identified using confirmation tests: Gram stain, mobility at 25°C in motility agar, catalase test, oxidase test, fermentation of xylose and rhamnose, hemolysis test on sheep blood agar plates, CAMP test against Staphylococcus aureus and Rhodococcus equi, and the API Listeria® (BioMérieux, Marcy l'Etoile, France).
Serotyping
Serotyping was carried out using commercial specific antisera (Denka Seiken, Tokyo, Japan) against the serovars 1/2a, 1/2b, 1/2c, 3a, 3b, 3c, 4c, 4d/4e, and 4b/4e, following the manufacturer's instructions (Ueda et al., 2002).
Multilocus sequence typing
Genomic DNA was extracted using GenomicPrep® cell and tissue isolation kit (Amersham, Piscataway, NJ) following the manufacturer's instructions. DNA concentrations were determined at a wavelength of 260 nm using the spectrophotometer DU640 (Beckman, Fullerton, CA). The final concentration of DNA was adjusted to 10 ng/mL with RNAse, DNAse, and Protease-free water (Sigma-Aldrich, Milan, Italy).
Seven DNA sequences belonging to the abcZ, bglA, cat, dapE, dat, ldh, and lhkA genes were amplified by polymerase chain reaction (PCR) and sequenced. All genes were amplified using previously published primers (Salcedo et al., 2003), except for the ldh gene, for which the primers suggested from a modified MLST scheme (
Antimicrobial susceptibility testing
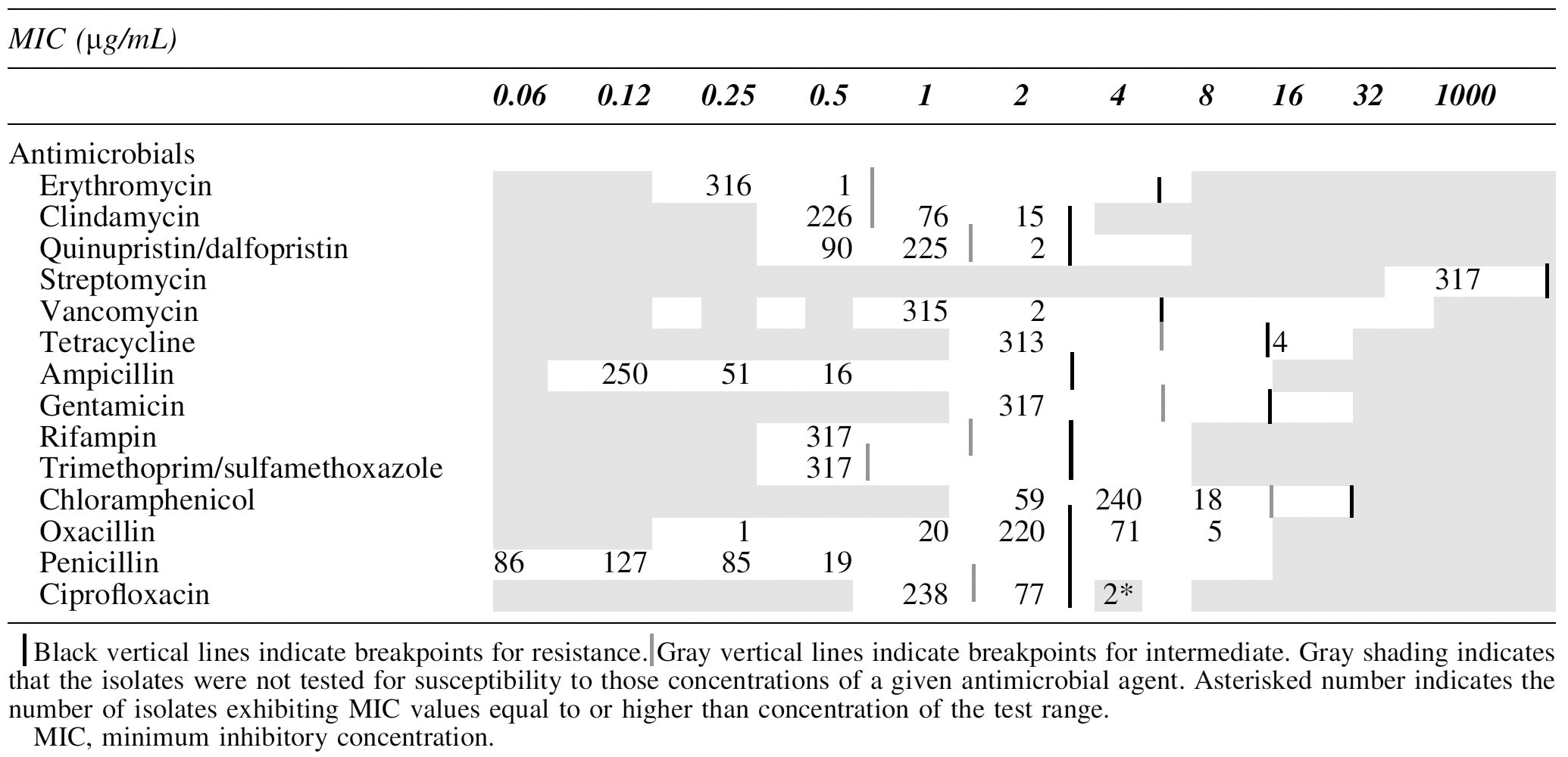
Isolates were subjected to minimum inhibitory concentration (MIC) using Sensititre® plates GPN4F and GPALL1F (Trek Diagnostic Systems, Cleveland, OH) specific for Gram-positive bacteria and including a wide variety of antibiotics at a full range of MIC: erythromycin (0.25–4 μg/mL), clindamycin (0.5–2 μg/mL), quinupristin/dalfopristin (0.5–4 μg/mL), streptomycin (1000 μg/mL), vancomycin (1–32 μg/mL), tetracycline (2–16 μg/mL) ampicillin (0.12–8 μg/mL), gentamycin (2–16 μg/mL), rifampin (0.5–4 μg/mL), trimethoprim/sulfamethoxazole (0.5/9.5–4/76 μg/mL), chloramphenicol (2–16 μg/mL), oxacillin +2% NaCl (0.25–8 μg/mL), penicillin G (0.06–8 μg/mL), and ciprofloxacin (1–2 μg/mL). The MIC against L. monocytogenes isolates was assessed following the broth microdilution method reported in Clinical and Laboratory Standards Institute (CLSI) M45-A3 document accordingly CLSI breakpoints were used for MIC categorization (CLSI, 2016). In brief, isolated colonies were suspended in sterile demineralized water to achieve a 0.5 McFarland. From the bacterial suspension, 10 μL were transferred to 90 μL Mueller–Hinton Broth with lysed horse blood (Trek Diagnostic Systems) and inoculated onto the MIC plates, which were incubated 24 h at 35°C. MIC values were manually recorded. Escherichia coli ATCC 25922 and Enterococcus faecalis ATCC 29212 were used as quality control strains. Each experiment was performed in duplicate.
As CLSI breakpoints for L. monocytogenes only include ampicillin, penicillin, and trimethoprim/sulfamethoxazole, and L. monocytogenes belongs to the Bacillales order, CLSI breakpoints for Bacillus were used for the following antimicrobials: erythromycin, clindamycin, vancomycin, tetracycline, gentamicin, rifampin, chloramphenicol, and ciprofloxacin (document CLSI M45-A3) (CLSI, 2016; Yan et al., 2016). CLSI breakpoints for Staphylococci, Enterococci, and Corynebacteri were used for oxacillin, streptomycin, and quinupristin/dalfopristin, respectively (CLSI M45-A3, CLSI VET01-S2) (CLSI, 2013, 2016).
Statistical analysis
Chi-square test was performed using StatView 5.0 software (SAS 155 Institute Inc., USA) to compare nominal variables between serotypes, lineages, and sequence types.
Results
Serotype, lineage, sequence type
Serotyping of the 317 L. monocytogenes strains identified the following serotypes: 1/2a (43%; n = 136), 1/2c (26%; n = 84), 4b/4e (15%; n = 46), 1/2b (12%; n = 38), 3a (2%; n = 7), 3b (1%; n = 3), 3c, 4c, and 4d/4e (0.32%; n = 1).
In the population of L. monocytogenes, there are three lineages: the lineage I includes isolates of serotype 4b/4e, 1/2b, 3b, 4d/4e, and 3c, the lineage II isolates of serotype 1/2a, 1/2c, and 3a, and the lineage III isolates of serotype 4a and 4c. Of the 317 strains, 72% (227/317) belonged to the lineage II, 28% (89/317) to the lineage I, and 0.31% (1/317) to the lineage III.
MLST analysis of the 317 strains identified 52 different sequence types. The most representative STs were ST9 (25%; n = 80), ST121 (17%; n = 54), ST2 (7%; n = 21), ST3 (5%; n = 16), and ST8 (4%; n = 14). The results of MLST analysis, serotyping and Lineage distribution of the 317 isolates, and their source are summarized in Table 1.
Characteristics of the 317 Listeria monocytogenes Isolates
ST, sequence type.
Antimicrobial susceptibility
The MIC distribution of the 317 isolates is reported in Table 2. Using the breakpoints given, all isolates were susceptible to erythromycin, streptomycin, vancomycin, ampicillin, gentamicin, rifampin, trimethoprim/sulfamethoxazole, chloramphenicol, and penicillin. From all isolates tested, 54% (170/317) were intermediate or resistant to certain antimicrobials. In detail, of the 317 isolates tested, 91 (29%) were intermediate to clindamycin, 77 (24%) to ciprofloxacin, and 2 (1%) to quinupristin/dalfopristin; 76 (24%) were resistant to oxacillin, 4 (1%) to tetracycline, and 2 (1%) to ciprofloxacin. Among all intermediate or resistant isolates, 59% (101/170) were intermediate or resistant to only one antimicrobial, 33% (56/170) to two antimicrobials, and 8% (13/170) to three antimicrobials (Table 3).
Minimum Inhibitory Concentration Distribution of the 317 Listeria monocytogenes Strains
Antimicrobial Resistance Patterns of Listeria monocytogenes Strains
Underlined names represent intermediate susceptibility.
CIP, ciprofloxacin; CLI, clindamycin; OXA, oxacillin; SYN, quinupristin/dalfopristin; TET, tetracycline.
In relationship to the origin, the percentage of oxacillin-resistant isolates was significantly higher in the environment in comparison to food and humans (46%, 19%, and 35%, respectively) (p < 0.05) (Table 4), while the percentages of both clindamycin and ciprofloxacin intermediate strains were significantly higher in human isolates in comparison to food and the environment (60%, 23%, and 29%, respectively, for clindamycin intermediate isolates and 70%, 19%, and 3%, respectively, for ciprofloxacin intermediate isolates) (p < 0.001) (Table 4).
Antimicrobial Resistance and Origin of the 317 Listeria monocytogenes Strains
I, number (%) of intermediate strains; R, number (%) of resistant strains; S, number (%) of sensitive strains.
In relationship to the lineage, the percentage of oxacillin-resistant strains was significantly higher in isolates of lineage I (58/89; 65%) compared with lineage II (18/227; 8%) (p < 0.001) (Table 5) (Fig. 1). No significant differences were found between the percentages of clindamycin and ciprofloxacin intermediate strains belonging to the two lineages (Fig. 1) (Table 5).
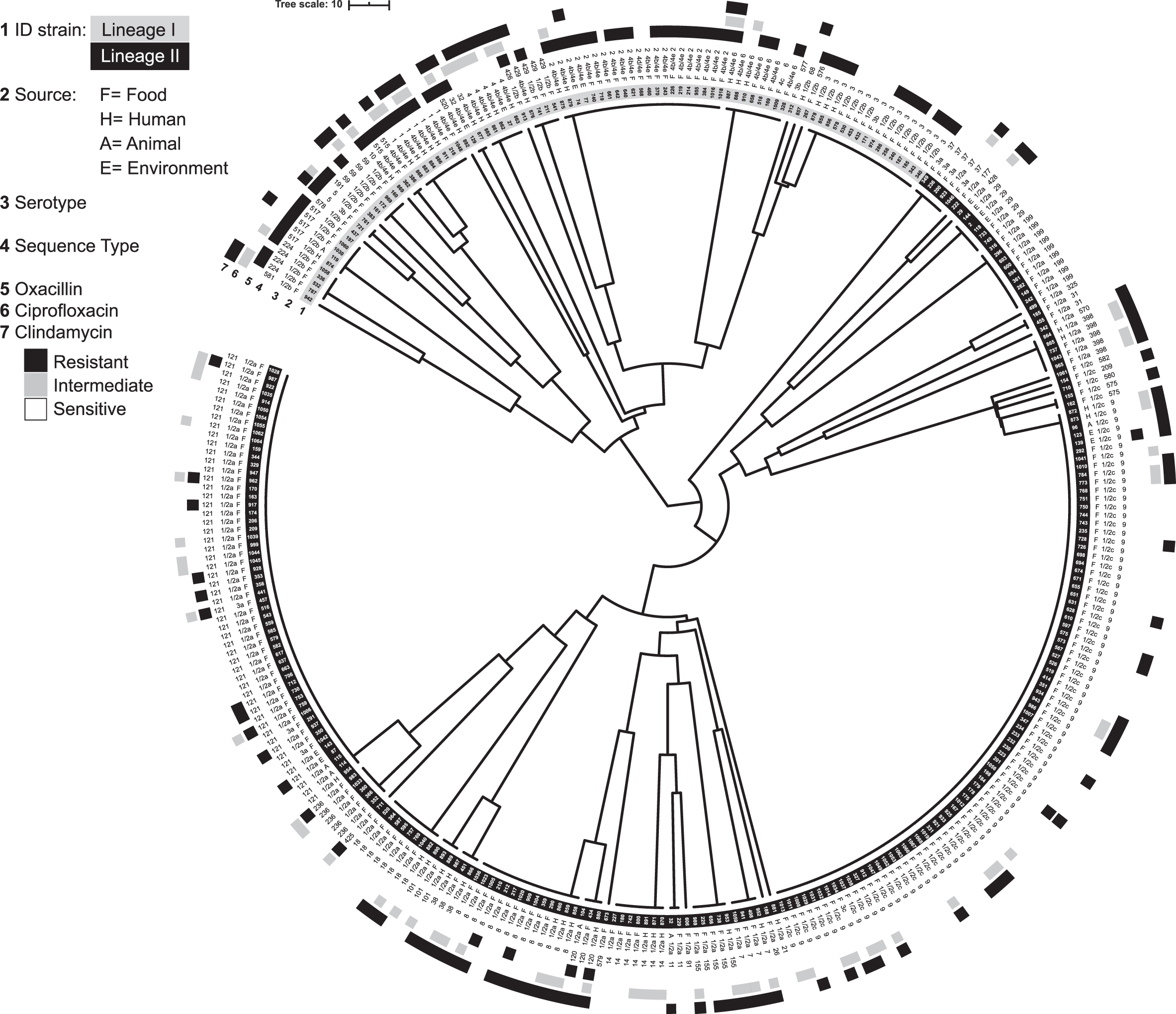
Unweighted pair group method with arithmetic mean tree based on multilocus sequence type profiles. Rings are as follows: strain ID (1), source (2), serotype (3), sequence type (4), oxacillin susceptibility (5), ciprofloxacin susceptibility (6), and clindamycin susceptibility (7).
Antimicrobial Resistance and Lineage of the 317 Listeria monocytogenes Strains
I, number (%) of intermediate strains; R, number (%) of resistant strains; S, number (%) of sensitive strains.
In relationship to the serotype, the percentages of oxacillin-resistant as well as clindamycin and ciprofloxacin intermediate strains were significantly higher in isolates of serotype 4b/4e (72%, 35%, and 33%, respectively) (p < 0.05) (Table 6).
Antimicrobial Resistance and Serotype of the 317 Listeria monocytogenes Strains
I, number (%) of intermediate strains; R, number (%) of resistant strains; S, number (%) of sensitive strains.
Isolates were collected in a broad time period ranging from 1998 to 2009. Comparing the last 3 years (2007–2009) to previous years (1998–2006), a clear positive trend of increase was observed in the percentage of resistant and intermediate isolates per year. In particular, the percentage of oxacillin-resistant and clindamycin intermediate strains increased from 22% to 28% and from 26% to 35%, respectively, and the percentage of ciprofloxacin intermediate strains increased from 21% to 34% (Table 7). The increase was statistically significant only for ciprofloxacin intermediate strains (p < 0.05).
Antimicrobial Resistance of the 317 Listeria monocytogenes Strains Over Time
I, number (%) of intermediate strains; R, number (%) of resistant strains; S, number (%) of sensitive strains.
Discussion
The Listeria genus was described as susceptible to antibiotics active against Gram-positive bacteria, including ampicillin or penicillin (combined with aminoglycosides), trimethoprim (alone or combined with sulfamethoxazole), tetracyclines, erythromycin, and gentamicin (Rota et al., 1996; Teuber, 1999; Yücel et al., 2005). High susceptibility of isolates to ampicillin (100%), penicillin (100%), tetracyclines (99%), gentamicin (100%), erythromycin (100%), and trimethoprim-sulfamethoxazole (100%) were observed in the present study (Table 1). Similar results were also previously reported (Conter et al., 2009; Yan et al., 2010; Barbosa et al., 2013; Wang et al., 2013; Madeo et al., 2015).
Few isolates showed resistance to tetracycline (4/317; 1.3%) and ciprofloxacin (2/317; 0.6%), whereas a consistent number of isolates showed resistance to oxacillin and intermediate resistance to clindamycin or ciprofloxacin (24%, 28.7%, and 24.3%, respectively). Also, intermediate resistance to quinupristin/dalfopristin was observed in 2 (0.6%) L. monocytogenes isolated from water and plant (environmental source).
In our study, resistance to oxacillin was 24% for L. monocytogenes isolated from selected sources. Oxacillin resistance has been already associated to L. monocytogenes with even higher percentages in research studies from Brazil, United States, and Poland (Davis and Jackson, 2009; Wieczorek et al., 2012; Camargo et al., 2015). In Staphylococcus infections, oxacillin susceptibility test is used as a marker for methicillin-resistant Staphylococcus aureus. This resistance is associated with the mecA gene coding for an alternative penicillin-binding protein namely PBP2A with low-binding affinity to β-lactam antibiotics (Reichmann and Pinho, 2017). Further studies need to be performed to verify whether similar molecular mechanisms of oxacillin resistance are true also for Listeria.
In the present study >20% of L. monocytogenes isolates showed intermediate MIC values for clindamycin and ciprofloxacin. These intermediate values are particularly interesting. Acquired resistance to both antimicrobials is a multistep process linked to the subsequent acquisition of multiple point mutations in core genes. In particular, clindamycin resistance was associated with methylation and mutations in the 23S rRNA (Kehrenberg et al., 2005; Poehlsgaard et al., 2005; Kulczycka-Mierzejewska et al., 2018). Ciprofloxacin resistance was associated to mutations in the quinolone resistance determining region of gyrA, overexpression of efflux pump gene llde and mutation in the fepR gene, and regulator of the efflux pump protein gene fepA in L. monocytogenes (Lampidis et al., 2002; Godreuil et al., 2003; Moreno et al., 2014; Jiang et al., 2018; Wilson et al., 2018).
Intermediate MIC values suggest a probable future shift to resistance phenotypes in case of acquisition of additional mutations. Significant percentages of clindamycin and ciprofloxacin resistance have been already described worldwide (Davis and Jackson, 2009; Wieczorek et al., 2012; Camargo et al., 2015; Noll et al., 2017).
An interesting statistical correlation has been found between resistance and lineages/serotypes. In particular, oxacillin resistance was significantly more frequently identified in lineage I and in particular in serotype 4b/4e. Similarly, the percentages of isolates with intermediate clindamycin or ciprofloxacin MIC values were significantly higher in serotype 4b/4e. A correlation between some L. monocytogenes serotypes and antimicrobial resistance has been previously reported although no explanation for these observations has been proposed yet. For instance, Barbosa et al. (2013) found that resistance to erythromycin was correlated with the serotype 4b, whereas no correlation was observed between ciprofloxacin resistance and L. monocytogenes serotypes (Safdar and Armstrong, 2003). On the contrary, other authors found that serotype 1/2a isolates were more frequently resistant to ciprofloxacin compared to serotype 4b (Safdar and Armstrong, 2003; Kovacevic et al., 2013). Besides, the association of antimicrobial resistance or reduced susceptibility to serotype 4b is of concern, since strains belonging to this serotype were associated to outbreaks (Althaus et al., 2017; Buchanan et al., 2017; Burall et al., 2017).
The two most frequently identified STs among intermediate and resistant isolates to clindamycin, ciprofloxacin, and oxacillin were ST9 and ST2, respectively. These two STs have been found in different food categories. The high level of exposure might be the reason of the high contribution to listeriosis cases of these STs (Henri et al., 2016).
Along with single antimicrobial resistance or reduced susceptibility, 8% of resistant isolates showed resistance to three or more antimicrobials and therefore fall within the definition of multidrug-resistant isolates (Table 3) (Magiorakos et al., 2012). Results from other studies reported multidrug resistance in L. monocytogenes isolated from food and animals to chloramphenicol, erythromycin, gentamicin, kanamycin, streptomycin, and rifampin (Charpentier et al., 1995; Walsh et al., 2001; Srinivasan et al., 2005). However, Godreuil et al. (2003) reported that only five of 488 clinical isolates of L. monocytogenes were susceptible to chloramphenicol and vancomycin (Walsh et al., 2001).
Although L. monocytogenes was noted to be relatively susceptible to a wide range of antimicrobials as few as 20 years ago, a number of more recent reports suggests that the rate of antimicrobial resistance in L. monocytogenes is increasing from 1% to 10% up to 50% (Vela et al., 2001; Walsh et al., 2001; Godreuil et al., 2003; Barbosa et al., 2013; Gómez et al., 2014; Thønnings et al., 2016; Arslan et al., 2019). Accordingly, in the present study, a median increase per year of the percentage of antimicrobial resistant or reduced susceptible isolates to oxacillin, clindamycin, and ciprofloxacin was observed in the period 2007–2009 in comparison to previous years (1998–2006). These results are consistent to a previous Italian study based on a large antimicrobial susceptibility survey conducted in 1995. In this study, all 148 isolates were susceptible to β-lactams, lincosamides, and tetracycline (Aureli et al., 2003). Conversely, in a more recent study performed in France on 4816 clinical L. monocytogenes strains isolated between 1926 and 2007, although low, resistance to specific antimicrobials (tetracyclines and fluoroquinolones) was found to be emerging (Morvan et al., 2010).
In France, a 10-year period study, mostly overlapping the years in the present work (1996–2006) isolated only 4 resistant isolates among the 202 food and environmental samples analyzed (Granier et al., 2011). This is in contrast with the observation of the present study, in which 170 isolates were identified resistant over 317 isolates analyzed. Although it must be said that in the present dataset, isolates of human origin were also included, their relative few number (43) does not significantly bias the comparison.
In comparison to previous studies, results suggest significant different trends of antimicrobial-resistant L. monocytogenes from food and the environment in European neighboring countries in the first decade of the century. In addition, in line with the concept of epidemiological cutoff values, evaluation of clinical intermediate resistance in L. monocytogenes might represent a useful tool to predict future occurrence of resistant strains against antibiotics associated with a multistep acquisition mechanism. If confirmed on additional L. monocytogenes isolates collected in more recent years, the Italian increase of resistant and intermediate L. monocytogenes isolates to oxacillin, clindamycin, and ciprofloxacin is a matter of concern. This observation is particularly relevant especially in case of antimicrobial-resistant isolates of serotype 4b, which is repeatedly associated with food-borne outbreaks.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Italian Ministry of Health (RC IZSPB 05/08).