
Abstract
Campylobacter jejuni is the leading cause of human foodborne enteritis worldwide. Poultry products are regarded as the main source of human campylobacteriosis. Strategies are being developed to reduce colonization of poultry by Campylobacter. The membrane transport protein CjaA was reported to stimulate mucosal immune responses, which can reduce the C. jejuni load in chickens. In this study, oral immunization of broilers with food-grade Lactococcus lactis NZ3900/pNZ8149 carrying the C. jejuni cjaA gene was examined for the ability to reduce colonization of broilers by Campylobacter. The Usp45 signal peptide and the Escherichia coli heat-labile enterotoxin B subunit (LTB) gene fragments were inserted into the upstream and downstream of the cjaA gene for secretory expression and immune enhancement, respectively. The cjaA gene and the fusion cjaA-ltb gene were both expressed in recombinant L. lactis, and the single cjaA gene was secretory expressed in the recombinant strain. Oral administration of two recombinant L. lactis strains expressing the cjaA gene and the fusion cjaA-ltb gene both stimulated specific anti-CjaA serum IgY responses significantly. While the average intestinal sIgA responses in these groups were higher compared with the control groups, they were not significantly different. Chicken challenge experiments showed that the colonization levels of C. jejuni in the groups provided oral immunization with two recombinant L. lactis-delivered CjaA strains were significantly lower than that of the control group at 5 d postinoculation, but there was no significant difference in C. jejuni colonization among all groups at 9 d. These results indicated that recombinant L. lactis with secretory expression of CjaA is a promising live vector vaccine against C. jejuni colonization of chickens. The immunization regimen requires further optimization to ideally stimulate detectable levels of intestinal sIgA to enhance the level of inhibition of C. jejuni colonization.
Introduction
As one of the main foodborne bacterial pathogens, Campylobacter jejuni is the leading cause of human bacterial enteritis worldwide (Snelling et al., 2005). This organism is strongly associated with acute, self-limited gastrointestinal illness as well as Guillain-Barré syndrome (Hegde and Ahluwalia, 2005). C. jejuni is considered to be a commensal organism of poultry; most cases of the disease are caused via consumption of contaminated chicken products (Young et al., 2007). Thus, poultry are the major reservoir of C. jejuni and the main source for human campylobacteriosis (Young et al., 2007; Lin, 2009; Skarp et al., 2016). Reducing the colonization level of C. jejuni in poultry plays an essential role in reduction of the incidence of campylobacteriosis (Rosenquist et al., 2003; Jagusztyn-Krynicka et al., 2009; Newell et al., 2011).
At the poultry farm level, vaccination is still a promising approach to reduce intestinal C. jejuni loads (Meunier et al., 2017; Poly et al., 2018). To meet a successful C. jejuni vaccine for chicken, some properties such as cost-effectiveness and ease of delivery to massive numbers of chicken should be taken into account (de Zoete et al., 2007). For this purpose, the live vectored vaccines delivering the protective antigen of C. jejuni may be the most promising candidates. Live vectors such as attenuated Salmonella strains, oocysts of Eimeria tenella, or Lactococcus lactis have been used for delivering the antigen candidates of C. jejuni (Clark et al., 2012; Łaniewski et al., 2014; Kobierecka et al., 2015, 2016; Meunier et al., 2016).
Lactic acid bacteria have been extensively studied as a live vector for heterologous protein delivery to induce specific mucosal and systemic immune responses upon oral administration (Wells and Mercenier, 2008; Bermudez-Humaran, 2009; Bahey-El-Din and Gahan, 2010; Wells, 2011; Plavec and Berlec, 2019). Kobierecka et al. (2016) first evaluated the L. lactis IL1403 strain as a mucosal delivery vehicle, and observed a moderate reduction of C. jejuni colonization in chicken. To ensure safety and effective expression of heterologous genes, a commercial food-grade expression platform L. lactis NZ3900/pNZ8149 (MoBiTec GmbH, Germany) was used in this study (Mierau and Kleerebezem, 2005). Furthermore, Escherichia coli heat-labile enterotoxin subunit B (LTB), as one of the most powerful mucosal adjuvants known (Newsted et al., 2015; Sun et al., 2017), and the usp45 signal peptide assisting for secretory expression (Borrero et al., 2011) were coexpressed with CjaA in tandem to elicit robust mucosal and systemic immune responses.
In this study, the antigen candidate cjaA gene, the usp45 signal peptide, and the ltb gene fragments were inserted into the expression plasmid pNZ8149 individually or in combination. The recombinant plasmids were transformed into the food-grade L. lactis strain NZ3900 for oral administration to chickens. The immunogenicity and protective efficacy of the L. lactis-vectored CjaA vaccines were evaluated after oral immunization and challenge trial in chicken.
Materials and Methods
Bacterial strain, plasmids, and culture conditions
The major bacterial strains and plasmids used in this study are listed in Supplementary Table S1. C. jejuni NCTC 11168 was grown at 42°C in a microaerophilic incubator as described previously (Xu et al., 2015). L. lactis NZ3900 was grown in M17 medium (Difco, BD) supplemented with 0.5% glucose at 30°C without shaking. E. coli was grown in Luria–Bertani broth with shaking (200 rpm) or on agar (Difco, BD) at 37°C. When needed, culture media were supplemented with ampicillin (100 μg/mL), kanamycin (50 μg/mL), or nisin (1 ng/mL).
Polymerase chain reaction
The major polymerase chain reaction (PCR) primers used in this study are listed in Supplementary Table S2. PCR components and conditions were performed as described previously (Xu et al., 2010). PCR products were purified by the QIAquick PCR Purification Kit (Qiagen) for cloning and sequence.
Construction of L. lactis vaccine strains
Three recombinant plasmids pNZ8149s, pNZ8149s-cjaA, and pNZ8149s-cjaA-ltb were constructed as shown in Figure 1A. To construct pNZ8149s, an 81-bp signal peptide fragment of the usp45 gene was amplified from the plasmid pGH-SPusp45 using the primers SPusp45F/SPusp45R (Supplementary Table S2) and inserted into pNZ8149 with double digestion of NcoI and SphI. To construct pNZ8149s-cjaA, a 768-bp gene fragment of cjaA (containing the stop codon) was amplified from C. jejuni NCTC 11168 using the primers CjaA-2F/CjaA-2R (Supplementary Table S2) and inserted into pNZ8149s with double digestion of SphI and SpeI. To construct pNZ8149s-cjaA-ltb, a 765-bp gene fragment of cjaA (deleting the stop codon) was amplified using the primers CjaA-2F/CjaA-3R (Supplementary Table S2) and inserted into pNZ8149s with double digestion of SphI and SpeI. Then, a 387-bp gene fragment of LTB was amplified from the plasmid pGH-LTB using the primers LTBF/LTBR (Supplementary Table S2) and inserted into the recombinant plasmid with double digestion of SpeI and XbaI. The three recombinant plasmids pNZ8149s, pNZ8149s-cjaA, and pNZ8149s-cjaA-ltb were transformed into the host strain L. lactis NZ3900 via electroporation to obtain the recombinant L. lactis strains NZ3900-pNZ8149s, NZ3900-sCjaA, and NZ3900-sCjaA-Ltb, respectively.
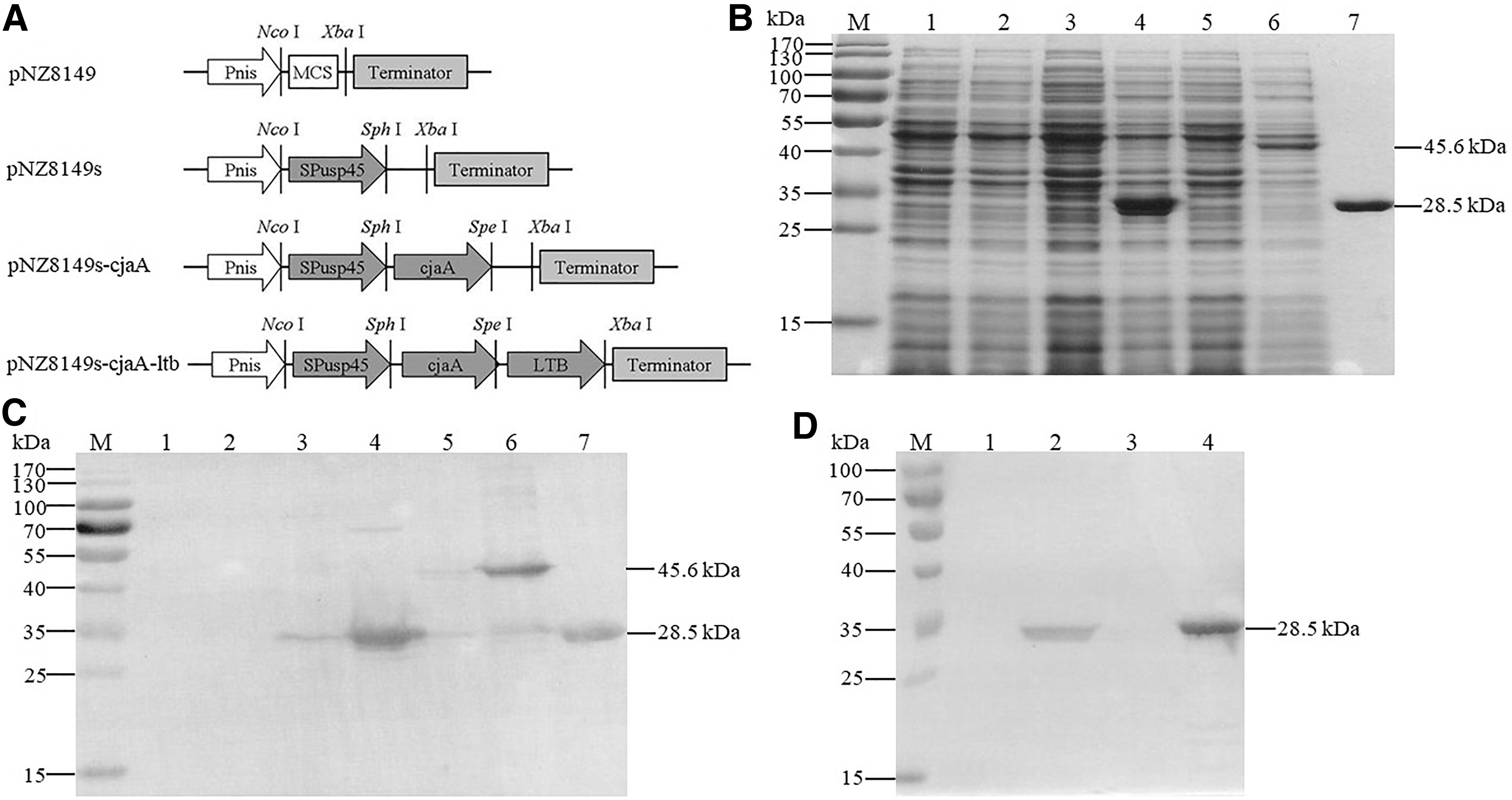
Analysis of rCjaA expression and secretion from recombinant Lactococcus lactis.
Production of recombinant CjaA in E. coli and generation of polyclonal antisera
A histidine-tagged rCjaA was produced in E. coli BL21 (DE3) using pET-28a vector (Novagen). Briefly, a 768-bp fragment covering 255-aa CjaA was PCR amplified from C. jejuni NCTC 11168 using the primers CjaA-1F/CjaA-1R (Supplementary Table S2). The amplified product and the vector pET-28a were both digested with NcoI and XhoI and ligated onto each other. After cloning and expressing successfully, the rCjaA was purified by the AKTA purifier 100 system (GE Healthcare, Bucks, United Kingdom) using a HisTrap FF affinity chromatography column. The highly purified rCjaA was used for production of rabbit polyclonal antisera against CjaA. The rabbit CjaA polyclonal antisera were prepared by Beijing Genomics Institute (Beijing, China).
SDS-polyacrylamide gel electrophoresis (SDS-PAGE) and immunoblotting
The overnight cultures of all L. lactis strains were inoculated into fresh GM17 medium with a dilution of 1:100, grown to an OD600 of 0.4–0.5 at 30°C and induced by nisin (1 ng/mL). After incubation for 3 h, bacterial pellets and supernatants were harvested by centrifugation. Supernatants were passed through 0.22 μm filters and concentrated using Amicon Ultra-4 Centrifugal Filter Units (Millipore Sigma, St. Louis, MO). The concentrated supernatants were dialyzed by phosphate buffer saline (PBS) (pH 7.2) for further use. The pellets were resuspended in PBS and lysed by ultrasonication on ice. The lysates and the concentrated supernatants were used for SDS-PAGE and subsequent immunoblotting following the standard procedures.
Animal experiment approval
The chicken experiment was in accordance with the Guidelines of Animal Care and Use Committee of the Institute under the approval of Institute of Animal Husbandry and Veterinary Medicine (permit no.: 2014-05). All efforts were made to alleviate animal suffering and improve their quality of life.
Oral immunization and challenge of chickens
One-day-old specific pathogen free (SPF) chickens were purchased from Beijing Boehringer Ingelheim Vital Biotechnology Co., Ltd. and randomly divided into 5 groups with 18 birds per group in separate isolators. The chickens were allowed to acclimatize for 3 d, and flocks were determined to be negative for Campylobacter by culturing cloacal swabs before use for immunization (Xu et al., 2010). Four groups were orally inoculated with L. lactis strains of NZ3900-sCjaA-Ltb, NZ3900-sCjaA, NZ3900-pNZ8149s, and NZ3900-pNZ8149 with a dose of 2 × 1010 CFU of bacteria per chicken, respectively. The chickens in the control group were orally inoculated with the GM17 culture medium. The birds were immunized from day 5 to day 11, and then reimmunized from day 19 to day 25. To evaluate the persistence of the orally inoculated recombinant L. lactis in the chicken intestine, the chicken fecal samples were collected at day 12, 14, 16, and 18 from all the groups. Total DNA was extracted from the fecal samples for amplifying the partial fragment of pNZ8149 using the primer pairs pNZ8149F/pNZ8149R (Supplementary Table S2). All chickens were weighed at day 5, 19, and 33 to evaluate the growth performance during the immunization period. At day 33, the blood samples were collected from the wing vein of each chicken to monitor anti-CjaA-specific IgY antibody and the intestinal lavage samples were collected from euthanized chickens (6 birds/group), as described previously (Kobierecka et al., 2016), to monitor the sIgA antibody response.
At day 33, all the remaining chickens were orally challenged of C. jejuni NCTC 11168 with a dose of 1.5 × 106 CFU/chicken. For each group, six chickens were euthanized and cecal contents were collected on day 38 and 42, 5 and 9 days postinoculation (DPI), respectively. The cecal contents from each chicken were weighed and diluted in MH broth. The diluted samples were cultured on MH agar plates with Campylobacter-specific selective supplements (SR117E; Oxoid, Basingstoke, England) for enumeration of C. jejuni as described previously (Xu et al., 2010).
Enzyme-linked immunosorbent assay
The anti-CjaA-specific antibody level in sera and intestinal lavage samples was determined by an indirect enzyme-linked immunosorbent assay (ELISA), as described previously (Kobierecka et al., 2016), with some modifications. Briefly, 96-well microtiter plates were coated with purified rCjaA at a concentration of 1.6 μg/mL overnight at 4°C. Sera and intestinal lavage samples were diluted at 1:10 in ELISA dilution buffer as primary antibody. Optimal dilutions of 1:10,000 for horseradish peroxidase-conjugated anti-chicken IgY or IgA antibodies (Sigma-Aldrich) were used as a secondary antibody. The optical density values at 450 nm were measured using an ELISA reader (Synergy H1; BioTek Instruments, Inc.).
Statistical analysis
Statistical analysis was performed using a one-way analysis of variance (ANOVA). Duncan's multiple range test was used to compare the differences among the treatment groups. A p-value of less than 0.05 was taken to indicate statistical significance.
Results
Production of highly purified rCjaA and antiserum
The rCjaA had a molecular mass of ∼28.5 kDa as determined by SDS-PAGE (Supplementary Fig. S1), in accordance with the calculated molecular mass of CjaA. Approximately 3 mg of the highly purified 6xHis-tagged rCjaA was used for production of rabbit polyclonal antisera.
Production of rCjaA in L. lactis
Introducing the usp45 signal peptide into the recombinant plasmids was used to obtain an extracellular form of CjaA and CjaA+LTB (Fig. 1A). Expression of rCjaA and rCjaA-LTB in L. lactis transformants was analyzed by SDS-PAGE and Western blot using anti-CjaA-specific sera (Fig. 1B/C). With induction of nisin, ∼28.5 and 45.6 kDa proteins were produced in NZ3900-sCjaA and NZ3900-sCjaA-Ltb, respectively (Fig. 1B), in accordance with the calculated molecular mass of CjaA and CjaA+LTB. Western blot analysis also showed that the rCjaA and rCjaA-LTB proteins reacted with the anti-CjaA sera specifically (Fig. 1C). Moreover, Western blot analysis of the concentrated supernatants showed that only the band of ∼28.5 kDa secreted by NZ3900-sCjaA was observed (Fig. 1D). No band of the fusion protein CjaA+LTB was detected in the supernatant of NZ3900-sCjaA-Ltb (Fig. 1D).
Body weight of chickens
The mean body weight of each group is shown in Supplementary Figure S2. No statistically significant differences in body weight were observed within the five groups throughout the trial.
Persistence of recombinant L. lactis in the chicken intestine
The positive sample with a 321-bp PCR product was counted in each group. All chickens were detected positive on 1 d, and about half of the chickens were detected positive on 7 d after the oral immunization (Table 1), which indicated that the recombinant L. lactis only persists in chicken intestine for a certain period.
Persistence of Recombinant Lactococcus lactis in the Chicken Intestine
Persistence of recombinant Lactococcus lactis in the chicken intestine was evaluated by amplifying the partial fragment of pNZ8149 using the primer pairs pNZ8149F/pNZ8149R.
The chicken feces samples were collected after the first consecutive 7-d (day 5–11) oral immunization.
Positive samples/total samples.
Serum and intestinal antibody responses induced by oral immunization
The ELISA results showed that the anti-CjaA sera IgY levels in the groups of NZ3900-sCjaA-Ltb and NZ3900-sCjaA were significantly higher than those in the groups of NZ3900-pNZ8149s, NZ3900-pNZ8149, and culture medium (p < 0.01) (Fig. 2). Moreover, the sera IgY level of the NZ3900-sCjaA group was significantly higher than that of the NZ3900-sCjaA-Ltb group (p < 0.05) (Fig. 2). While the mean levels of intestinal sIgA detected in NZ3900-sCjaA- and NZ3900-sCjaA-Ltb-immunized groups were higher compared with the other treatment groups, no statistically significant differences were detected (Fig. 2).
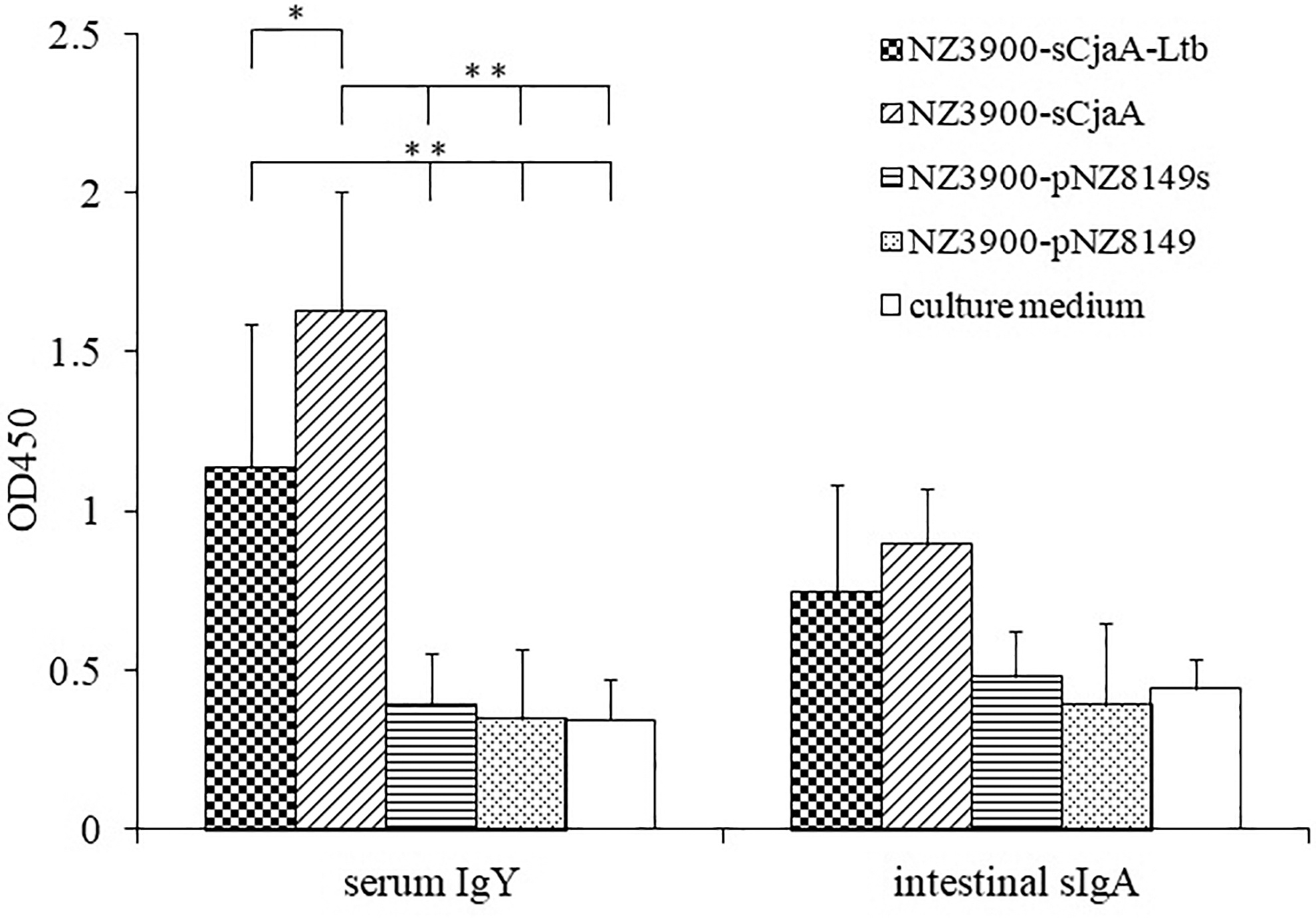
Serum IgY and intestinal sIgA antibody titers to rCjaA. The serum IgY and intestinal sIgA antibody titers of chickens from the five groups were determined at 33 d by indirect ELISA. Sera and intestinal lavage samples were diluted at 1:10 in ELISA dilution buffer as primary antibody. Each bar is the average of OD450 readings from six individual serum samples or intestinal lavage samples with standard error. Two asterisks indicate that the anti-CjaA sera IgY levels in the groups of NZ3900-sCjaA-Ltb and NZ3900-sCjaA were significantly higher than those in the other three groups, respectively (p < 0.01). An asterisk indicates significant difference (p < 0.05) between the NZ3900-sCjaA-Ltb group and NZ3900-sCjaA group. ELISA, enzyme-linked immunosorbent assay.
Colonization of C. jejuni
At 5 DPI, chickens in the NZ3900-sCjaA and NZ3900-sCjaA-Ltb groups showed a reduced colonization level compared with the other three groups (Fig. 3). The NZ3900-sCjaA group had a significant reduction of cecal C. jejuni (p < 0.05) by 2.35 log10 and 2.05 log10 compared with the NZ3900-pNZ8149s and culture medium groups, respectively (Fig. 3). At 9 DPI, the majority of chickens in each group were colonized by NCTC 11168 with an average shedding level of 106–107 CFU/g feces and no statistically significant differences were observed within the five groups (Fig. 3). The results indicated that the rCjaA secreted by L. lactis NZ3900 plays an essential role in reducing colonization level at the early stage of C. jejuni colonization.
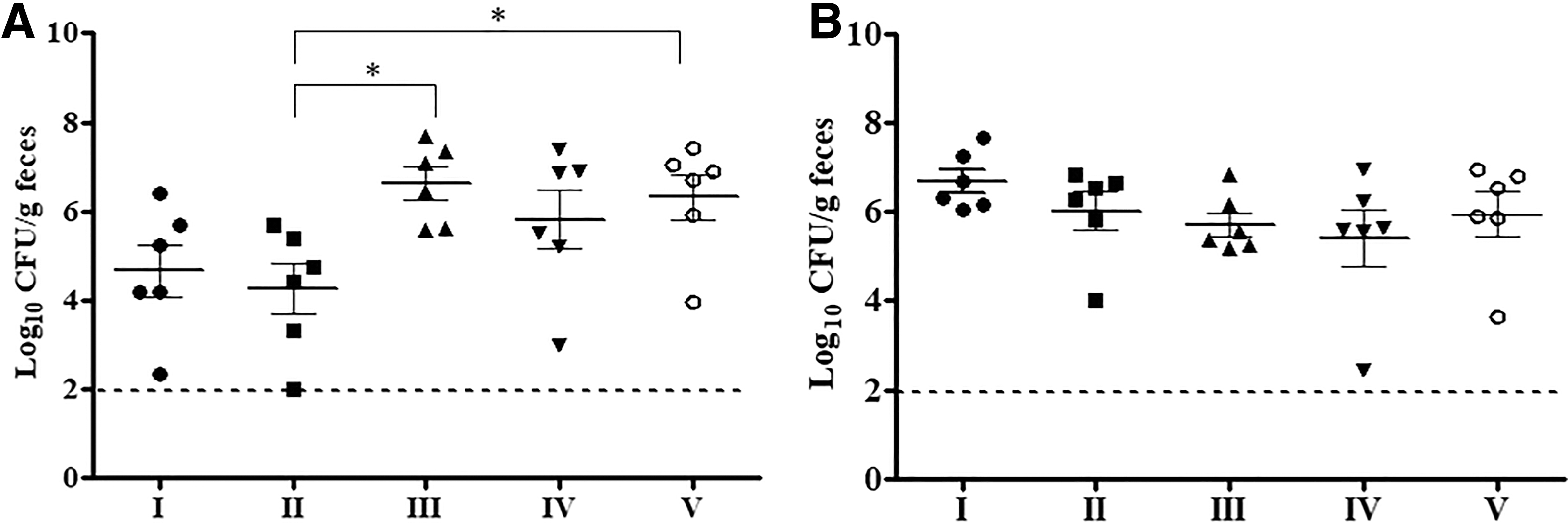
Effects of oral immunization with recombinant Lactococcus lactis on colonization of Campylobacter jejuni in chickens on 5 d
Discussion
Surface-exposed colonization proteins and capsular polysaccharides play essential roles in stimulating immune responses and protection against C. jejuni colonization (Neal-McKinney et al., 2014; Meunier et al., 2017; Nothaft et al., 2017; Poly et al., 2018). The membrane transport protein CjaA had been expressed in different live vectors to effectively stimulate the mucosal immune response and reduce C. jejuni colonization in chicken (Wyszynska et al., 2004; Clark et al., 2012; Kobierecka et al., 2016). Wyszynska et al. (2004) reported that avirulent Salmonella-delivered CjaA reduced C. jejuni colonization by 6 log10 in chicken. About 1–2 log10 reduction of C. jejuni colonization had been reported in the studies of E. tenella-delivered CjaA and L. lactis-delivered rCjaAD (Clark et al., 2012; Kobierecka et al., 2016). This study indicated that L. lactis NZ3900-delivered CjaA had a reduction of ∼2 log10 on C. jejuni colonization in chicken at 5 DPI. A quantitative microbial risk assessment showed that reducing the number of Campylobacter on chicken carcasses by 2 log10 causes a 30-fold reduction in the incidence of campylobacteriosis in humans (Rosenquist et al., 2003), which indicated that the recombinant L. lactis strain NZ3900-sCjaA is a promising vaccine candidate.
The localization of the expressed antigen in L. lactis plays an essential role in stimulating the mucosal immune responses. The antigen either secreted or surface displayed (cell wall-anchored) has a better effect on immunogenicity than that of intracellular location (Wells, 2011; Plavec and Berlec, 2019). Kobierecka et al. (2016) had reported that L. lactis harboring the surface-exposed rCjaAD could elicit specific anti-rCjaAD sIgA, but no correlation was noticed between the levels of the induced immune response and antigen localization. However, in this study, the NZ3900-sCjaA group with the secretory form of CjaA elicited a higher antibody level of sera IgY and sIgA compared with the NZ3900-sCjaA-Ltb group, which indicated that secretory form of CjaA plays an essential role in stimulating immune response.
L. lactis is a noncommensal and noncolonizing bacterium in animal intestine; it can survive passage through it for up to 5–7 d (Perdigon et al., 2001). In this study, persistence of L. lactis NZ3900 in chicken intestine indicated that the bacteria were completely shed to the environment in half of the chickens within 7 d. Unlike the traditional vaccination of one or two doses, frequent dosing over several weeks via oral administration may be required for an effective L. lactis vaccine (Wells, 2011). Frequent dosing may be a disadvantage for the L. lactis vaccine. However, it has been suggested that the risk of eliciting immunological tolerance can be diminished compared with the persistent strains.
The LTB adjuvant has proven to be a promising mucosal adjuvant for its nontoxicity and potent activity (Verweij et al., 1998; Millar et al., 2001). Sun et al. (2017) reported that the LTB protein was expressed as secretory form in the L. lactis NZ3900/pNZ8149 platform and significantly enhanced the mucosal sIgA antibody responses. In this study, the fused CjaA-LTB protein was coexpressed in L. lactis, but secretory expression failed to be detected by Western blot. The sera IgY level of the NZ3900-sCjaA-Ltb group was significantly lower than that of the NZ3900-sCjaA group. The immunostimulatory failure of LTB may be due to the intracellular expression, which needs to be investigated in the future work.
Footnotes
Acknowledgment
We are grateful to Dr. Maojun Zhang (National Institute for Communicable Disease Control and Prevention, Chinese Center for Disease Control and Prevention) for providing the C. jejuni NCTC 11168 strain.
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by grants from the National Key Research and Development Program of China (2018YFD0500506); the National Natural Science Foundation of China (31572527); and the Special Program on Science and Technology Innovation Capacity Building of BAAFS (KJCX201914).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.