
Abstract
Necrotizing enterocolitis (NEC) is a serious inflammatory intestinal disorder with a high mortality rate, which occurs most commonly in newborn infants. Cronobacter sakazakii, a common contaminant in infant formula, is associated with NEC. However, its role in NEC pathogenesis is unknown, and there are still no effective treatments for NEC. Currently, natural bioactive products have been investigated for their beneficial effects in preventing microbial infection. In this study, a neonatal mouse intestinal inflammation model was used to examine the protective effects of citral (a natural bioactive product) on C. sakazakii-induced intestinal inflammation and damages. It was shown that citral reduced the number of C. sakazakii cells in ileal tissues, and mice treated with citral had a significantly higher body weight than C. sakazakii-infected mice. Citral treatment also ameliorated serious ileal tissue damages, including epithelial sloughing, villous rupture, and enterocyte apoptosis. C. sakazakii infection upregulated the messenger RNA transcription levels of several inflammation-associated genes, increased production of IL-6 and TNF-α, and activated the NF-κB and MAPK signaling pathways in ileal tissues. Citral treatment mitigated these inflammatory responses. The apoptotic index and activities of caspase 3, 8, and 9 increased in murine ileum after C. sakazakii infection, but citral inhibited both enterocyte apoptosis and activations of these caspase. These findings suggest that citral has protective effects on C. sakazakii-induced intestinal inflammation in newborn mice, and it may play a future role in the management of C. sakazakii-associated infections and diseases.
Introduction
Necrotizing enterocolitis (NEC) is a severe inflammatory intestinal disorder that affects primarily preterm neonates and is associated with high mortality rates, ranging from 30% to 60% (Fredriksson and Lilja, 2019). The incidence of NEC which occurs in approximately 15% of premature infants weighing <1000 g in United States neonatal intensive care units has not changed over the last three decades (Samuels et al., 2016). NEC is characterized by intestinal epithelial necrosis, full-thickness intestinal destruction, bacterial invasion and translocation, inflammatory infiltration, intestinal epithelial barrier breakdown, sepsis, and death (Schmolzer et al., 2006; Thompson and Bizzarro, 2008). Several factors, including prematurity, formula feeding, and gut bacteria colonization (Lin et al., 2008b), may cause NEC by stimulating inflammatory cytokines, alterations in intracellular signaling pathways, and enterocyte apoptosis (Hunter et al., 2009; Zhou et al., 2017; Yan et al., 2019). Notably, the Gram-negative Cronobacter species have been isolated from the blood, peritoneal fluid, feces, and alimentary canal of NEC-affected infants (Chan et al., 1994; Almajed and Forsythe, 2016). Powdered infant formulas contaminated with Cronobacter sakazakii have been implicated in outbreaks of NEC (van Acker et al., 2001), suggesting that this pathogen may play an important role in NEC.
C. sakazakii is an important emerging opportunistic foodborne pathogen that is prevalent in milk-based powdered infant formula (Kandhai et al., 2004). Several researches showed that C. sakazakii could induce serious infection and intestinal inflammatory responses (Guner et al., 2011; Weng et al., 2014). The United States Food and Drug Administration (FDA) issued several warnings regarding C. sakazakii infection in newborns (Hunter et al., 2008). Although it has been proved that antibiotics provided protective effects in treating NEC (Silverman et al., 2017), drug-resistant species have emerged due to the widespread usage of antibiotics, presenting a huge challenge for treating NEC (Fjalstad et al., 2018). Besides antibiotics, several clinical trials suggested that a regimen combining the probiotics Bifidobacteria infantis and Lactobacillus acidophilus provided a protective effect for NEC (Lin et al., 2008a; Braga et al., 2011), and several probiotic strains have also been suggested as a beneficial role in preventing C. sakazakii-induced intestinal inflammation (Khailova et al., 2009; Kutylowksi and Yahia, 2019). However, the administration of live organisms to immunocompromised premature infants is prohibited by the FDA (Lin et al., 2008a). Some other treatments for NEC, including supportive care, radiologic monitoring, and surgical intervention, were also investigated (Elfvin et al., 2015; Shah et al., 2017). Natural antimicrobials from plant source have been widely explored for their direct antimicrobial activity or anti-infective potential (Aziz and Karboune, 2018). Our previous work showed that citral had a more potent antibacterial effect on C. sakazakii compared with some other natural antibacterial substances (Shi et al., 2016).
Citral (GRAS 182.60), the main bioactive component of Litsea cubeba oil, has been reported to exhibit multiple beneficial biological effects in vitro, including antiviral, anti-inflammatory, antineoplastic, antioxidant, and bacteriostatic (Hierro et al., 2004; Ortiz et al., 2010). It is also shown that citral exhibited anti-virulence effects for C. sakazakii in vitro (Shi et al., 2017). In this study, we extended the previous in vitro studies into a newborn mouse inflammation model to determine whether citral can protect against intestinal inflammation in newborn mice that are exposed to C. sakazakii.
Materials and Methods
Bacterial strain
C. sakazakii (strain No. 29544) (toxin genotype ST-8, serotype O1) was obtained from American Type Culture Collection (ATCC, Manassas, VA), and prepared as described previously with some modifications (Li et al., 2015). Briefly, C. sakazakii was prepared by the cultivation at 37°C overnight in tryptone soya broth (TSB; LandBridge, Beijing, China), after which the bacterial cells were recovered by centrifugation at 4°C, 8000 × g for 5 min, then washed, resuspended, and adjusted in phosphate-buffered saline (PBS, pH = 7.4) for animal experiment.
Animal experiment
Chinese Kunming Mice (Mus musculus Km) purchased from the Laboratorial Animal Center of Xi'an Jiaotong University (Xi'an, Shaanxi, China) (SCXK 2013–003), were kept under specific pathogen-free conditions. This study was carried out in accordance with recommendations of the “Guide for the Care and Use of Laboratory Animals: Eighth Edition, ISBN-10: 0-309-15396-4 (National Research Council, 2010).” The protocol was approved by the Laboratorial Animal Center of Xi'an Jiaotong University. Citral (high performance liquid chromatographically pure >99%; LGC Labor GmbH, Wesel, Germany) for animal experiments was 0.54 mg/mL (prepared in PBS), which is the minimum inhibitory concentration of citral against C. sakazakii ATCC 29544 (Shi et al., 2016). In addition, the preliminary experiments showed that citral at this concentration did not affect the normal growth, body weight, and intestinal morphology of the neonatal murine pups. Therefore, this concentration was chosen for this study. The murine pups were divided randomly into four groups (n = 30 per group) with different treatments: (1) PBS (normal saline), (2) Citral, (3) C. sakazakii infection (CS), and (4) Citral+CS. All pups were daily fed with 0.1 mL clean formula feed three times. The intestinal infection methods described by Emami et al. (2011) were followed with some modifications. In brief, from postnatal day 3 (P3), each pup was given PBS or citral through oral gavage once per day at night for eight sequential days. At P7 morning, each mouse pup in groups C and D received a single oral dose of 1 × 107 colony-forming unit (CFU) C. sakazakii for infection. All pups were weighed daily from P3 to P10. At P10 night (2 h after PBS/citral treatment), all pups were euthanized, and the intestinal tissues between the duodenum and rectum were collected.
The number of C. sakazakii in intestinal tissues
The fresh intestinal tissues were homogenized in cold sterile PBS (1:10, w/v). Serial dilutions were prepared and plated onto Chromogenic Cronobacter sakazakii Agar (LandBridge) and incubated at 37°C for 24 h to determine the number of C. sakazakii per mg of ieal tissue.
Intestinal histological assessment
The intestinal tissues (jejunum, ileum, and colon) were fixed in cold 4% paraformaldehyde/PBS overnight and then embedded in paraffin at 4°C. The paraffin-embedded tissues were sliced into 3 μm sections, mounted onto glass slides, dewaxed with xylene and ethanol, and stained using hematoxylin and eosin (Equal, Shannxi, China). The stained sections were observed with a light microscope at magnification of 100 × . The intestinal histopathology was analyzed by researchers who were blinded to the treatment conditions, following criteria (Table 1) set as previously described (Ran-Ressler et al., 2011).
Criteria for Intestinal Histopathological Scores
Real-time quantitative polymerase chain reaction
The total RNA in the intestinal tissues was extracted following the instructions of the Tiangen RNA Extraction Kit (Code No. DP430, Beijing, China).Then, RNA was converted to complementary DNA using a PrimeScript™ RT Reagent Kit (TaKaRa, Beijing, China). The primer sequences used in this study are shown in Table 2. Real-time quantitative polymerase chain reaction (PCR) was performed with QuantiTect SYBR Green 2 RT-PCR Master Mix (TaKaRa) for 40 cycles on the iQ5 Real-time PCR System (Bio-Rad), and the cycle threshold (Ct) values of real-time PCR were detected. The relative amount of each studied messenger RNA (mRNA) was normalized to glyceraldehyde-3-phosphate dehydrogenase (GAPDH) as a housekeeping gene, and the data were analyzed by the 2−ΔΔCt method (Livak and Schmittgen, 2001).
Primer Sequences for RT-qPCR
RT-qPCR, real-time quantitative polymerase chain reaction.
Apoptosis assay
Activity of caspase 3, 8, and 9
The tissues were ground on ice with lysate into homogenates, and then centrifuged (18,000 × g, 15 min, 4°C) to collect the supernatant. The caspase 3, 8, and 9 activities in the ileal tissues were determined according to the instructions of the kits from Beyotime Biotechnology Co., Ltd (Shanghai, China). After being incubated at 37°C for 4 h, the samples were measured with a microplate reader (Victor X3; PE, Singapore) at an absorbance of 405 nm to measure the caspase 3, 8, and 9 activities in the intestinal cells.
TdT-mediated dUTP Nick-End Labeling
Paraffin-embedded intestinal tissue sections were processed for TdT-mediated dUTP Nick-End Labeling (TUNEL), which stains the fragments of cellular genomic DNA exposed in the late stage of apoptosis, using a One-Step TUNEL Assay Kit along with 4′,6-Diamidino-2-Phenylindole (DAPI) (Beyotime) according to the supplier's instructions. Digital images of the TUNEL-labeled sections were obtained with a fluorescence microscope (DM5000 B; Leica, Germany). The fluorescence intensities were analyzed by Image Pro Plus 6.0 software to evaluate the apoptotic index of ileal cell apoptosis.
Sodium dodecyl sulfate–polyacrylamide gel electrophoresis and western blot
Intestinal tissues were homogenized on ice in lysis buffer supplemented with 1 mM phenylmethanesulfonyl fluoride, 2% protease and phosphatase inhibitor cocktail (Beyotime), then cleared by centrifugation at 12,000 × g for 5 min at 4°C. Equal amount of proteins (30 μg) were subjected to 12% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and electrotransferred onto a nitrocellulose membrane. Blots on the membrane were blocked in TBST (Tris-buffered saline, Tween 20) with 5% bovine serum albumin (Beyotime) for 2 h at room temperature and incubated with primary antibodies (1:1000 in TBST; Beyotime) at 4°C overnight. Subsequently, the membranes were washed three times with TBST for 20 min each, and incubated with horseradish peroxidase-conjugated secondary antibodies (1:1000 in TBST; Beyotime) for 90 min at room temperature. After washing, the blots were detected with ECL reagents (Beyotime) and ChemiDocXRS+System (Bio-Rad). Western blotting for GAPDH was performed to ensure equal sample loading, and the relative expressions of MAPK p38, phosphor-p38 (p-p38), NF-κB p65, and IκBα were detected.
Enzyme-linked immunosorbent assay
The intestinal tissues (50 mg) were homogenized in 500 μL of ice-cold PBS (containing 2% anti-protease cocktail), and then centrifuged at 3000 × g, 4°C for 20 min. The protein concentrations in supernatants were measured using a Bradford Protein Assay Kit (Beyotime). The expression levels of cytokines (IL-6 and TNF-α) were measured using the Enzyme-Linked Immunosorbent Assay Kits (Xinle, Shanghai, China). The cytokine concentration in the tissues was calculated as pg per mg of protein (pg/mg protein).
Statistical analysis
Results were analyzed by one-way analysis of variance or Student's t-test using SPSS statistics 22.0 (SPSS, Inc., Chicago, IL). Results are shown as the mean ± standard deviation (SD). Mean values labeled with different lower-case letters are statistically different from one another (p < 0.05).
Results
Citral reduced the number of C. sakzakii in murine ileal tissues
As shown in Figure 1A, 567 ± 115 CFU of C. sakazakii per milligram of ileal tissue were detected in C. sakazakii-infected mice. In contrast, there were no C. sakazakii observed in ileal tissues from C. sakazakii-infected mice treated with citral. Thus, citral treatment significantly decreased the number of C. sakazakii cells in the ileum of mice.

The number of C.sakazakii cells and histopathological scores of ileal tissues in neonatal mice.
Citral mitigated the intestinal morphological damage induced by C. sakazakii
Mice infected with C. sakazakii had significantly less weight increment from P3 to P10 compared with normal mice (p < 0.05) (Supplementary Fig. S1). Notably, C. sakazakii-infected mice that had received citral treatment had a significantly higher body weight growth compared with infected mice.
As shown in Figure 2, the jejunum and colon displayed mild injuries, whereas the ileum showed extensive necrosis. Therefore, only the ileal tissue used for further studies focused on the ileal tissues. C. sakazakii infection resulted in substantial pathological damage, including villus cell loss, epithelial sloughing, intestinal epithelial cell rupture, and intestinal architecture alignment disruption, with an average histopathological grading of 2.75 (Fig. 1B). In contrast, C. sakazakii-infected mice that received citral retained an orderly and intact villus architecture like that of uninfected controls, and villus tip sloughing, blunting, and cell loss were ameliorated, showing a significantly lower average pathological scores of 0.9 (p < 0.05) (Fig. 1B).
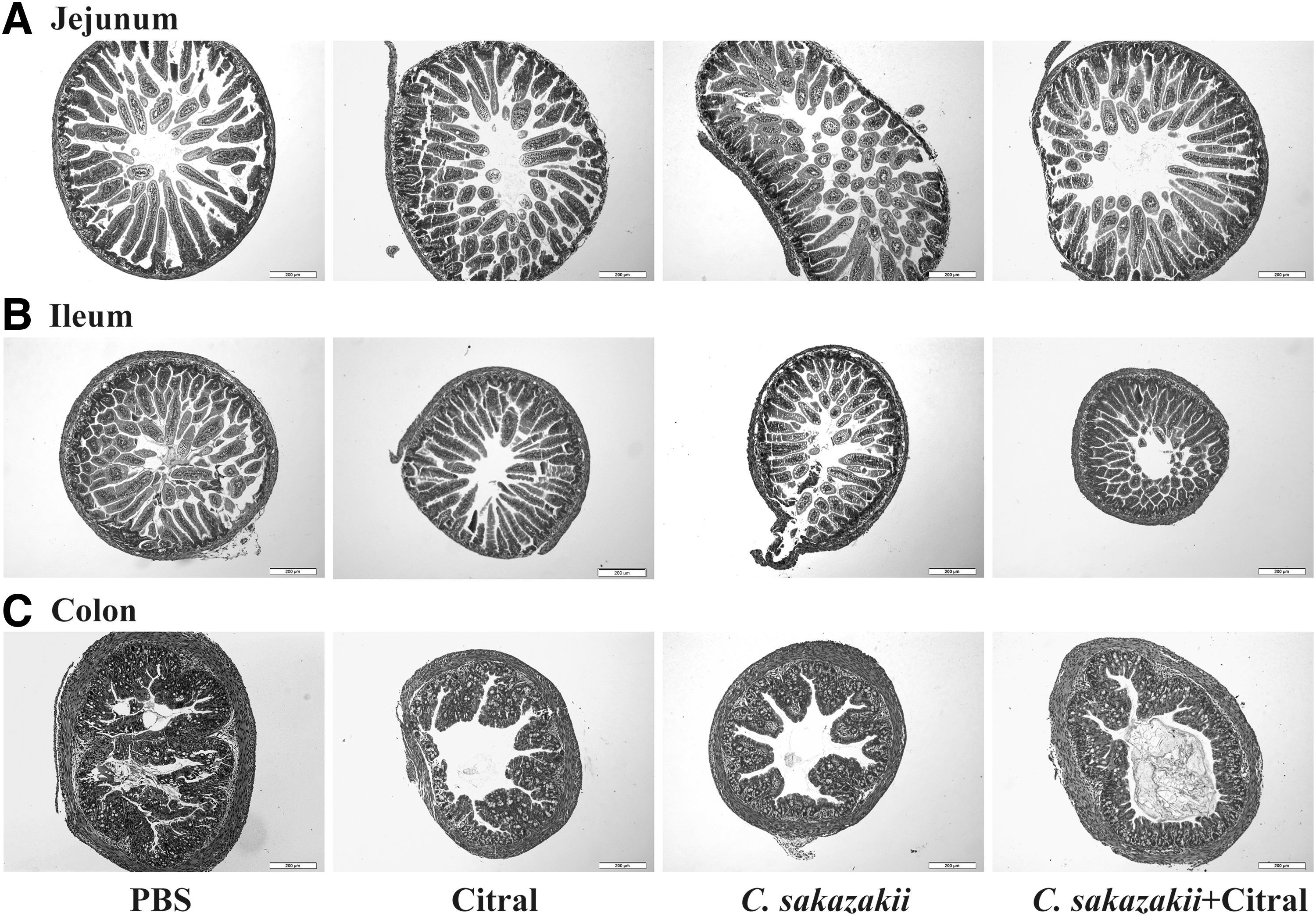
Hematoxylin and Eosin sections of intestinal tissues: jejunum
Citral affected the mRNA levels of seven inflammation-associated genes stimulated by C. sakazakii
The levels of seven inflammation-associated genes were determined using RT-qPCR (Table 3). Compared with normal pups, the mRNA levels of IL-6, PAF receptor, IFN-γ, IL-1β, TNF-α, NF-κB p65, and iNOS were found to be significantly higher in the ileal tissues of C. sakazakii-infected mice. In contrast, the mice treated with citral and infected with C. sakazakii had significantly lower mRNA transcription levels of inflammation-related genes. RT-qPCR was also employed to examine the levels of TLR4 mRNA, however, its levels were undetectable in all groups.
Fold Changes in the Messenger RNA Transcription Levels of Different Inflammation-Associated Genes in the Newborn Mouse Ileal Tissues
The expression levels of genes in different groups are shown as the “Mean ± standard deviation” (n = 10 mice in each group, one-way analysis of variance), and are marked with letters. For each gene, there are significant differences in the gene expression levels between the groups that are marked with different letters (e.g., a, b, c, d) (p < 0.05).
CS, C. sakazakii; PBS, phosphate-buffered saline.
Citral attenuated the intestinal cell apoptosis induced by C. sakazakii
The activities of caspase 3, 8, and 9 in the murine ileal tissues are shown in Figure 3. Citral treatment significantly attenuated the caspase 3 activation induced by C. sakazakii infection. Similarly, the caspase 8 activity in the ileal tissues of C. sakazakii-infected mice ileum were 1.58-fold higher than that in the control group, and caspase 8 activity was decreased to almost normal level in the infected mice treated with citral. Furthermore, citral treatment also reduced the caspase 9 activity in the ileal enterocytes of infected mice.
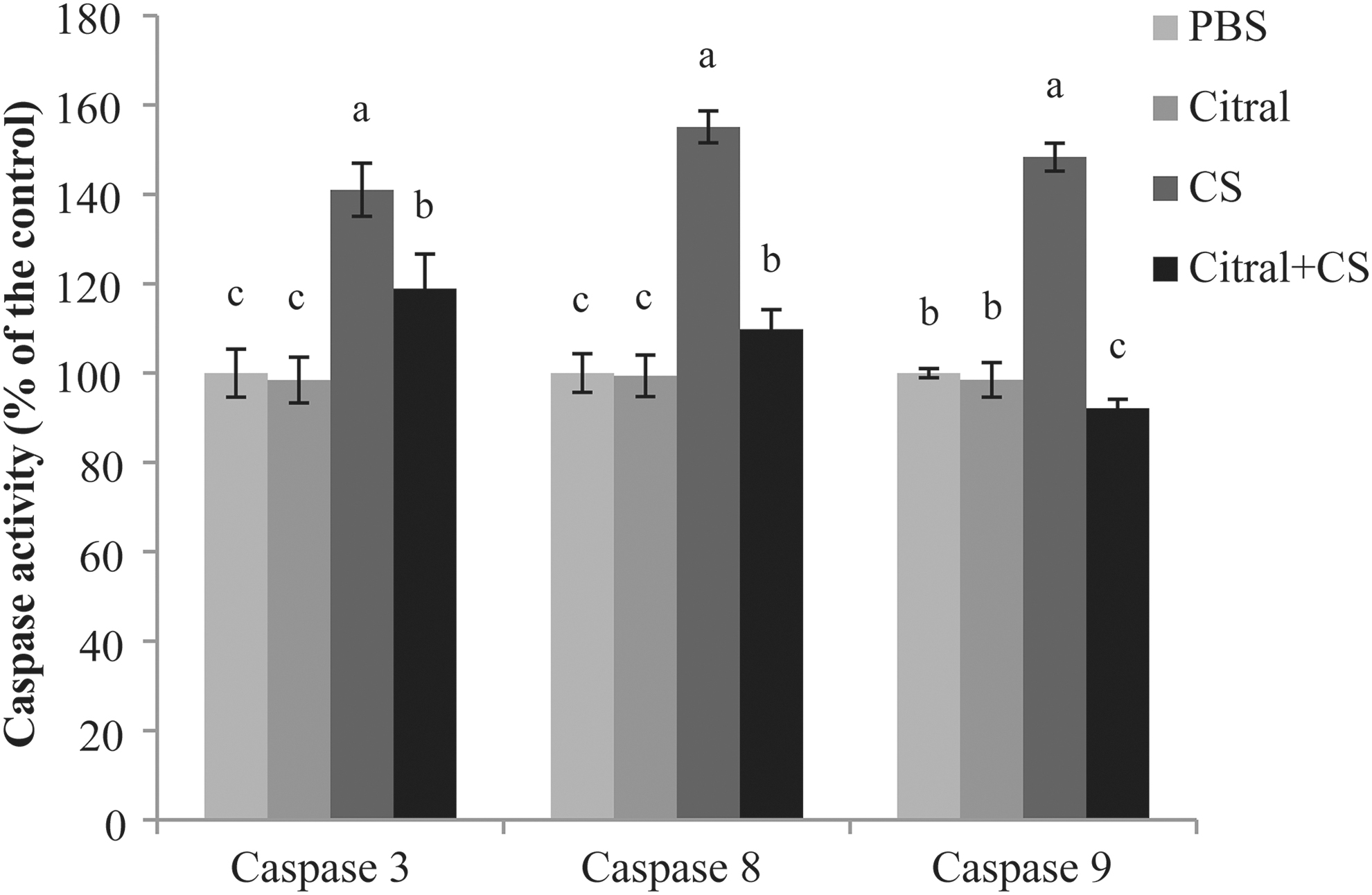
Caspase activity in ileal enterocytes from neonatal mice. Four different column colors stand for four different treatments (PBS, citral, C. sakazakii, or C. sakazakii+citral), taking caspase activity of PBS treatment mice as control for each caspase. Error bars represent the standard deviation (n = 10 mice per group), mean values with different lower-case letters (e.g., a, b, c) are statistically different from one another (one-way analysis of variance, p < 0.05). CS, C. sakazakii; PBS, phosphate-buffered saline.
TUNEL staining identifies apoptotic cells by labeling fragmented DNA with FITC (green fluorescence). The green fluorescence in the ileum sections of C. sakazakii-infected mice was extremely intensive, revealing that apoptosis was serious in the ileal tissues of the C. sakazakii-infected mice. Significant reductions of the green fluorescence signals were observed in the ileal tissues from C. sakazakii-infected mice treated with citral (Fig. 4). Therefore, citral significantly mitigated the enterocyte apoptosis in the ileal tissues that were induced by C. sakazakii.
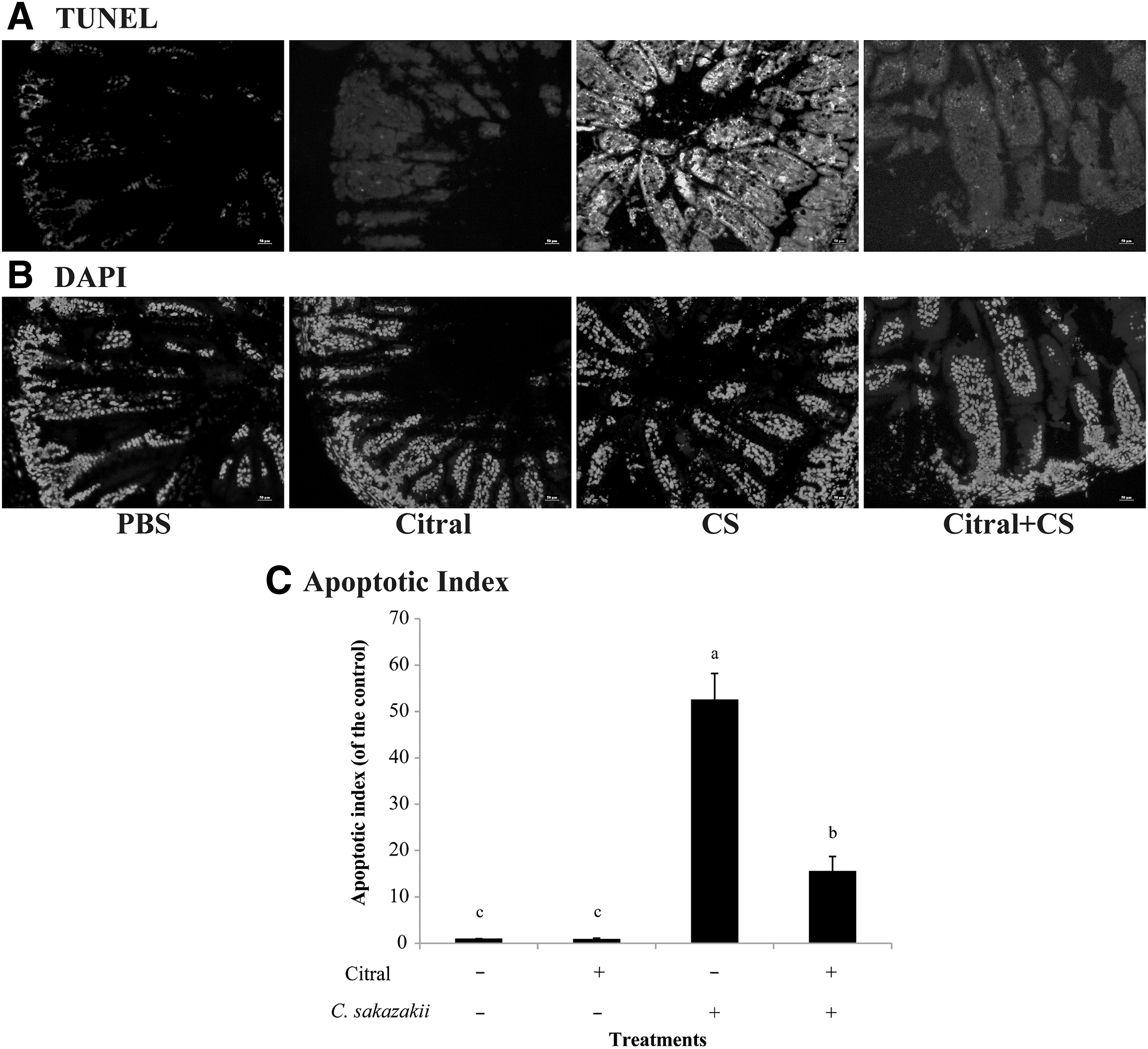
TUNEL labeling of ileum sections from the newborn mice in different groups. Representative images (at 400 × ) of TUNEL
Citral affected the expression levels of cytokines IL-6 and TNF-α stimulated by C. sakazakii
As shown in Figure 5A, IL-6 levels in mice subjected to both citral treatment and C. sakazakii infection were significantly lower than those in mice infected with C. sakazakii (9.30 ± 0.05 pg/mg vs. 11.65 ± 1.30 pg/mg, respectively). Similar results were observed for TNF-α (Fig. 5B); TNF-α protein expression levels in ileal tissues from C. sakazakii-infected mice were 126.26 ± 0.22 pg/mg; C. sakazakii-infected mice treated with citral had a significantly lower level of TNF-α expressions (81.83 ± 10.44 pg/mg).
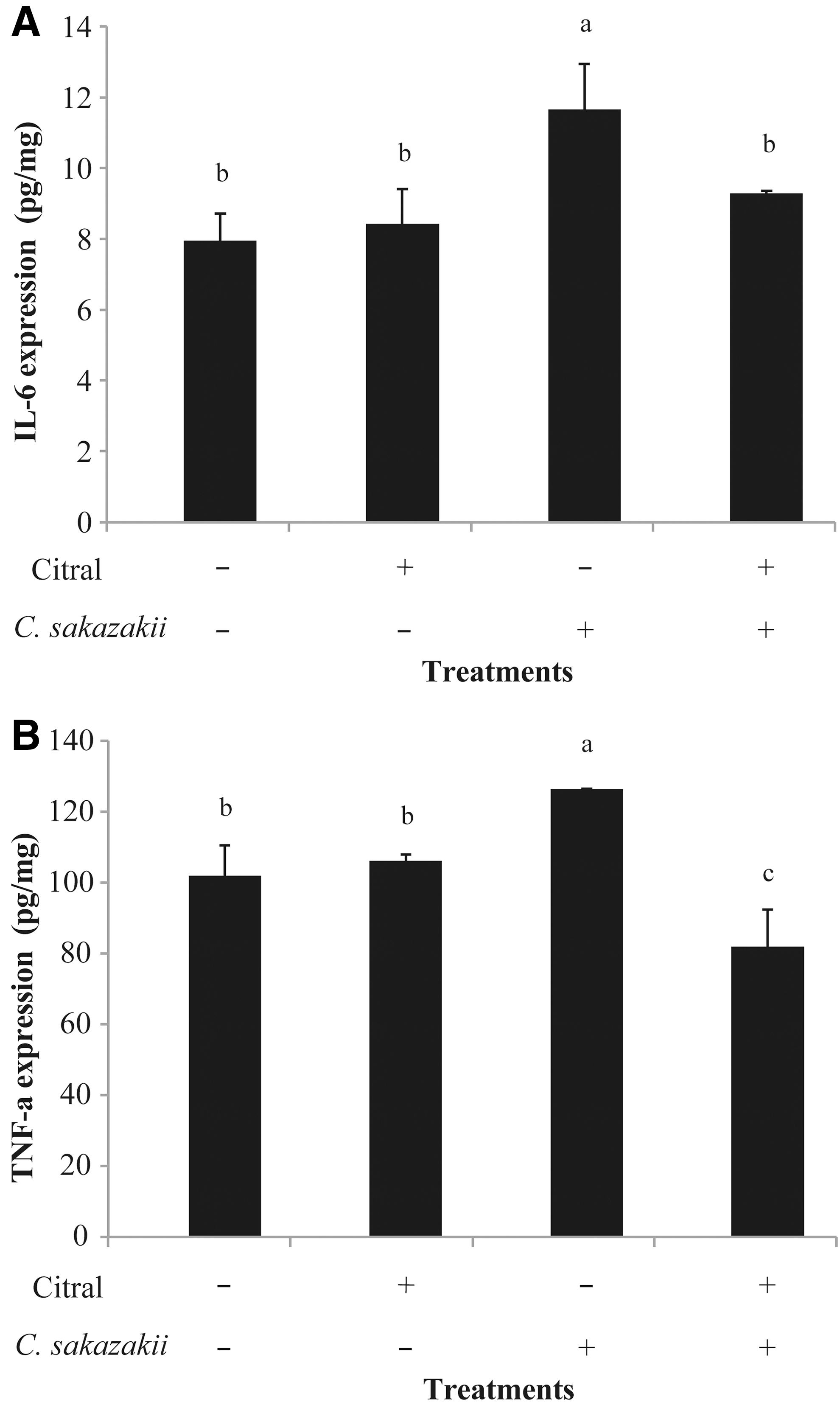
The protein expression levels of IL-6
Citral mitigated the signaling pathways activation induced by C. sakazakii
Western blotting assays were performed to examine the levels of key proteins in the NF-κB and MAPK signaling pathway in the murine ileal tissues (Fig. 6). Citral treatment attenuated the high expression levels of MAPK p38 and phosphor-p38 in murine ileal tissues that were induced by C. sakazakii infection. NF-κB p65 expression levels were significantly higher in the C. sakazakii-infected murine ileal tissues compared with control mice. However, the levels of IκBα in C. sakazakii-infected mice were substantially lower compared with control mice, suggesting an activation of NF-κB pathway. Citral treatment attenuated the increased levels of NF-κB p65 and increased IκBα protein expression in the C. sakazakii-infected mice.
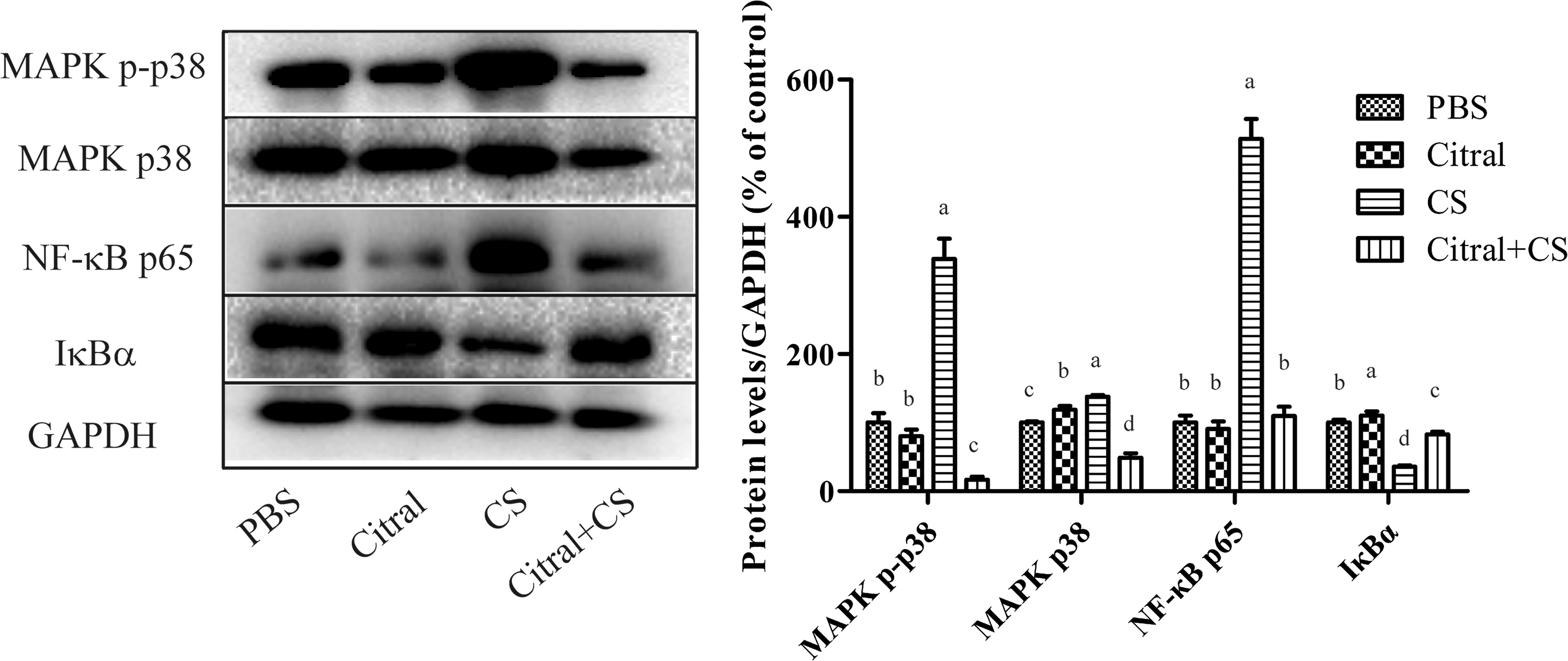
The protein expression of NF-κB p65, IκBα, MAPK p38, and phosphor-p38 in the ileal cells from the newborn mice. GAPDH was performed as the housekeeping protein to ensure equal sample loading. Representative western blot (left) and quantification (right) of MAPK p38, MAPK phosphor-p38, NF-κB p65, and IκBα expression in ileal cells from newborn mice treated with PBS, citral, CS, or citral+CS. Four different column colors stand for four different treatments, taking protein expression levels of PBS treatment mice as control for each protein. Error bars represent the standard deviation (n = 10 mice per group); mean values with different lower-case letters (e.g., a, b, c, d) are statistically different from one another (one-way analysis of variance, p < 0.05). CS, C. sakazakii; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; PBS, phosphate-buffered saline.
Discussion
C. sakazakii, a foodborne pathogen, has been linked to NEC, which is a devastating disease of neonates (Triulzi et al., 2006). Currently, natural antimicrobials have gained more and more attention because of their effectiveness (Nair et al., 2004). In this study, we investigated the anti-inflammatory effects of citral against C. sakazakii infection in newborn mice.
In this study, the mice were treated with citral from P3 to P10, C. sakazaii inoculation was performed at P7. Mice were pretreated with citral consistently for 4 d before C. sakazakii infection and 4 d following the bacteria infection. Generally, it will take a period of time for natural bioactive compounds to interact with and impact the host gut ecosystem and gut cell function (Feng et al., 2019). Therefore, the aim of our study is to determine the protective effects of citral against intestinal inflammation in newborn mice that are exposed to C. sakazakii.
Two recent studies in NEC patients found that, the permeability of the patients' intestinal tracts is increased, and the quantities of certain bacterial species in their intestinal tracts are changed at a large scale (Patel and DuPont, 2015; Maffei and Schanler, 2017). Due to these changes, it is possible that some Gram-negative bacteria may adhere to, invade, and proliferate in enterocytes and cause damage that is responsible for the onset of NEC (Hui et al., 2017). In this study, no C. sakazakii was detected in the intestinal tissues from infected mice treated with citral. It is possible that citral stimulated the immune system to eliminate the C. sakazakii bacteria.
Intestinal morphological damage has been typically regarded as a major disease marker for NEC (Thompson and Bizzarro, 2008). C. sakazkii caused serious damages to the intestinal villus structures, particularly in the ileum, and citral treatment significantly mitigated the infection-induced devastation of intestinal tissues. It was reported that, C. sakazakii can colonize and overload the gastrointestinal tract, leading to a loosening of the tight junctions among intestinal epithelial cells, which subsequently causes tissue injuries and acute inflammatory responses (Emami et al., 2012). C. sakazakii infection could induce the intestinal villi destruction, mucosal sloughing, submucosal edema, and goblet cell necrosis (Hunter et al., 2008; Emami et al., 2011). Similar to the previous study (Weng et al., 2014), C. sakazakii infection caused much severe damages in the ileal tissues than in the jejunum and colon tissues. Furthermore, the development of NEC can increase histopathological scores of the intestinal tracts to 2.0 (Khailova et al., 2009). In this study, citral significantly decreased the high histopathological scores of the murine intestinal tissues induced by C. sakazakii infection.
Similar to the previous study (Miyake et al., 2016), C. sakazakii infection negatively influenced mouse body weight growth but that citral treatment could reduce this effect. It was speculated that C. sakazakii infection led to the hypofunction of intestinal tissues, making it difficult for the body to absorb sufficient nutrition for normal growth and development.
Various inflammatory cytokines have been implicated in the pathogenesis of intestinal inflammatory responses and enterocyte apoptosis (Markel et al., 2006). C. sakazakii infection induced high mRNA and protein levels of cytokines IL-6 and TNF-α, which is similar to the clinical characteristics of NEC in the human infants (Neu and Walker, 2011); citral treatment significantly mitigated the infection-induced expression of IL-6 and TNF-α. Previous work also proved that the expression levels of IL-6 and TNF-α in gut tissue cells show a rapid upward trend in inflammatory responses (Hsueh et al., 2003; Zhang et al., 2016). From these findings, it is possible that citral stimulated the host immune systems to perform functions that prevent infection with C. sakazakii and partially regulate the inflammatory responses in murine enterocytes (Cruz-Cordova et al., 2012).
Previous work reported that the inflammatory response is associated with IFN-γ, which can stimulate iNOS and nitric oxide expression (Wang et al., 2017b. Citral was effective at inhibiting the increased levels of the iNOS and IFN-γ genes in ileal cells from C. sakazakii-infected mice in this study. TLR4 could activate the NF-κB signaling pathway when LPS interacts with the host cells (Li et al., 2017). However, we were unable to detect any TLR4 signals in our model through RT-qPCR, so the TLR4 mRNA expression in this system may be too limited to analyze. Altogether, these findings show that citral can significantly mitigate the cascading transcriptions of inflammation-associated cytokines in murine intestinal inflammatory responses to C. sakazakii infection.
The intestinal inflammation induced by C. sakazakii infection may be associated with the numerous damaged and dead intestinal tissue cells produced by infection (Blackwood et al., 2017). Apoptosis is a complex process, accompanied by the activation of a variety of regulatory factors, and the activation of caspase 8, 9, and 3, which are early markers of apoptosis, and execute different functions in cell apoptosis (Lin et al., 2017). Citral treatment reduced the caspase 3, 8, and 9 activities in the intestinal cells from C. sakazakii-infected mice. Previous work in a mouse model of NEC reported that TNF receptor activation could stimulate the activity of caspase 9 and 8, leading to the activation of caspase 3 and apoptosis (Brentnall et al., 2013). In the present study, citral reduced the high expression levels of TNF-α, caspase 3, 8 (extrinsic pathway), and 9 (intrinsic pathway) in the intestinal cells of C. sakazakii-infected mice. Thus, C. sakazakii may activate caspase pathways (both the extrinsic and intrinsic pathways), which then induce intestinal cell apoptosis, and citral may mitigate the intestinal cell apoptosis through the caspase pathway.
The attachment of C. sakazkii to enterocytes in the infected mice caused enterocyte apoptosis; this could be detected by TUNEL staining (Emoto et al., 2016). Citral treatment significantly attenuated the amount of ileal apoptotic cells in the C. sakazakii-infected mice. Thus, C. sakazakii infection induced ileal enterocyte apoptosis by activating the caspase pathway, and citral treatment significantly prevented enterocyte apoptosis in C. sakazakii-infected mice.
It has been shown that infection with some Gram-negative pathogens could activate the NF-κB signaling pathway in enterocytes (Zeng et al., 2006). C. sakazakii infection activated NF-κB p65 levels in the murine ileal tissues, and also decreased the IκBα expression, which may be the marker of NF-κB activation. Similarly, another study found that the secretion of TNF-α could activate the NF-κB signaling pathway, leading to the expression of proteins in the caspase family and inducing inflammatory responses and apoptosis (Wang et al., 2017c). The MAPK signaling pathway also regulates inflammatory cell infiltration, cytokine production, and apoptosis (Wang et al., 2017a). Citral was effective for reducing C. sakazakii-induced MAPK p38 and phosphor-p38 production. Thus, the inhibition of NF-κB and MAPK signaling pathways could be one of the possibilities by which citral exerts anti-inflammatory effects.
Conclusion
In summary, citral protected the mice from C. sakazakii-induced intestinal inflammation through reducing bacterial loads in intestinal tissues, mitigating morphological damages, reducing inflammatory cytokine expression, inhibiting signaling pathway activation, and preventing enterocyte apoptosis. It was possible that both the inhibitory activities of citral against C. sakazakii and the immunomodulatory effects of citral on the host contribute to the reduction of inflammatory response in newborn mice. Altogether, these findings suggest that citral may play a future role in the management of C. sakazakii-associated infections and diseases.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Fundamental Research Funds for the Central Universities (2452017228), National Natural Science Foundation of China (31801659 and 31772084), and General Financial Grant from the China Postdoctoral Science Foundation (No. 2017M623256).
Supplementary Material
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.