
Abstract
This investigation was performed to assess the supplementation of probiotics on cytokine expression and lymphocyte subpopulation in Campylobacter coli challenged chickens. Thirty-six individuals were equally separated into four experimental treatments: C = untreated chickens, LB = probiotic control (Lactobacillus fermentum), Cc = Campylobacter-challenged control, LBCc = probiotic + Cc. All chicks were slaughtered and cecum samples were collected on day 4 postinfection. Gene expression analysis, using reverse transcription quantitative PCR (RT-qPCR), revealed significant differences in cytokine transcript expression between untreated and probiotic-treated chickens. In addition, flow cytometry was used to quantitate the levels of lymphocyte subpopulations. Principal component analysis showed that probiotic administration induced an overall downregulation of cytokine expression. C. coli exposure provoked a similar response to that of L. fermentum but to a lesser extent. Colonization of C. coli in the presence of the probiotic evoked a complex response with an upregulation of some type II cytokines, including interleukin IL-4 and IL-13, which could explain the increased presence of antibodies in both lamina propria and epithelium. Moreover, despite that the percentage of CD8 intraepithelial lymphocytes (IELs) was found to be higher, downregulation of proinflammatory cytokines IL-15, IL-16, and interferon γ was observed. This suggests that the detected CD8 are not effector cells but induced IELs, which release antimicrobial peptides, and are ready to be primed upon encountering antigen. These outcomes demonstrate that probiotic administration promotes a humoral response to a C. coli infection while dampening any potential inflammation mediated by effector T cells in 1-week-old chicks.
Introduction
In humans, Campylobacter coli has been linked to gastrointestinal diseases and recent outbreaks have been reported in both developing and industrialized countries (Kuhn et al., 2017; Silva et al., 2018). Campylobacter spp. have been considered commensal bacteria in various wild and domestic animals, including chickens. However, because of its potential role as an avian infectious agent, it has been suggested to review such bacterial–host interaction (Awad et al., 2018). Expression of inflammatory cytokines is a key feature of pathogen-induced inflammatory processes, which lead to intestinal damage and high mortality (Hartley-Tassell et al., 2018).
Campylobacter jejuni elicits an inflammatory response starting in intestinal epithelial cells. Colonization by this enterobacteria induces expression of cytokines involved in the initiation (tumor necrosis factor α [TNF-α], interleukin 8 [IL-8]), promotion (IL-12, IL-23), and regulation (interferon γ [IFN-γ], IL-17) of inflammation, as well as in immune cell signaling (IL-4, IL-10) (Al-Banna et al., 2018). However, there are no abundant data regarding the inflammatory/immune response to infection with C. coli. This species of Campylobacter spp. has been linked to around one-third of cases reported in some European countries, and has been considered vigorous colonizers of the chickens' intestine (Doorduyn et al., 2010; European Food Safety, 2010). In any case, intensive consumption of chicken-related products is considered a major health risk factor (Skarp et al., 2016). Probiotics have been used to prevent intestine colonization by Campylobacter spp. (Mañes-Lázaro et al., 2017). In particular, Lactobacillus strains have been reported to elicit positive immunomodulatory effects by regulating expression of key immune mediators (Hu et al., 2015). Moreover, utilization of L. fermentum has been shown to reduce pathogenic inflammatory processes with a concomitant upregulation of the immune response in pathogen-challenged animals (Stašová et al., 2015). Hence, we decided to study the inflammatory and immune mechanisms underlying the intestinal response to infection with C. coli in the presence of the probiotic L. fermentum.
Materials and Methods
Chickens and experimental design
A total of 1-d-old 36 broiler cock chickens (COBB 500) were allocated in four groups of 9 animals. Chickens had access to water and feed (without probiotics, antibiotics, or anticoccidiostats) ad libitum throughout the experiment (8 d). Bacteria were administered per os. The first group was considered the control (untreated chickens, C), in which none of the bacteria were added; in the second (probiotic control [Lactobacillus fermentum], LB) and fourth groups (probiotic + Campylobacter-challenged control, LBCc), a suspension, of L. fermentum (109 colony-forming units [CFU]/0.2 mL) was administered on daily basis from days 1 to 7. A suspension of C. coli (108 CFU/0.2 mL) was administered on the fifth day to the third (Cc) and fourth (LBCc) groups (Table 1).
Schema of Experimental Design
C, control group; Cc, Campylobacter coli; dpi, days postinfection; LB, Lactobacillus fermentum.
Strain preparation
Lyophilized L. fermentum strain CCM7514 (provided by Radomíra Nemcová, Assoc. Prof. DVM, PhD), from intestine of domestic chickens, was resuspended in 1 mL phosphate-buffered saline (PBS), pH 7.4, and grown at 37°C for 48 h in De Mann–Rogosa–Sharpe (MRS) broth (Merck, Germany) inside an anaerostate (BBL GasPak™ Plus, Albany, NY). Solitary colonies (minimum five colonies) were inoculated into 50 mL MRS broth and incubated 24 h at 37°C. After cultivation, 450 mL of MRS broth were added and the culture was centrifuged at 2268 × g at 4°C for 45 min. The sediment was resuspended in 50 mL PBS to make a decimal dilution to determine the number of bacteria. Each chicken was orally inoculated with 109 CFU/0.2 mL of L. fermentum from the first day until the end of the experiment. C. coli (supplied by the Czech Culture Collection, Brno, Czech Republic) was grown in blood-free selective agar base CM0739 supplemented with SR0155 (containing Cefoperazone and Amphotericin B; Oxoid Ltd., United Kingdom) in a jar with microaerobic conditions (5% O2, 10% CO2, 85% N2, CampyGen; Oxoid Ltd.). Incubation was carried out at 42°C for 2 d. The majority of C. coli strains produced gray moist colonies that were harvested and diluted in PBS to the specific viable concentration (1 × 108 CFU/mL). Bacterial number was controlled by measuring optical density at 550 nm. Bacterial numbers were determined according to a standard microbiological method using serial dilution and posterior plating (Karaffová et al., 2017). By the fourth day, each chicken was inoculated with 108 CFU/0.2 mL PBS of C. coli per os.
Chicken sample collection
By the eighth day of the experiment (4 d after C. coli challenge), all individuals from each group were slaughtered and cecal samples were collected. For reverse transcription quantitative PCR (RT-qPCR), sections were kept in RNAlater (Thermo Scientific, Waltham, MA) and stored at −80°C. For flow cytometric analyses, sections from the same samples were used to assess the following subset of T cells: CD3, CD4, CD8, and CD45, together with immunoglobulins IgA and IgM following a previously described methodology (Bucková and Revajová, 2014). Lymphocytes were posteriorly isolated and purified (Solano-Aguilar et al., 2000). The experiments were performed following the protocol no. 863/17-221, according to the guidelines established by the Ethical Commission of the University of Veterinary Medicine and Pharmacy in Košice, and under the approval of the National Veterinary and Food Administration of the Slovak Republic.
RNA extraction, reverse transcription, and quantitative polymerase chain reaction assays
The RNeasy mini kit was used (Qiagen, Hilden, Germany) following the manufacturer's instructions. RNA concentrations and purity were assessed by spectrophotometric analysis using a NanoDrop 200c (Thermo Scientific). The synthesis of cDNA was carried out using iScript cDNA Synthesis Kit (Bio-Rad, Hercules, CA) following the user's manual. Expression levels of 84 different genes involved in mucosal immunity was determined using a RT2 Profiler PCR Array (Qiagen) and Maxima SYBR green qPCR Master Mix kit (Thermo Scientific). Three samples were analyzed simultaneously, and the average was used for statistical analyses. For normalization of expression, five housekeeping genes encoding the following proteins were used: β-actin (ACTB), hexose-6-phosphate dehydrogenase (H6PD), hydroxymethylbilane synthase (HMBS), ribosomal protein L4 (RPL4), and ubiquitin-C (UBC). The Ct values of studied genes were normalized to an average Ct value of the reference genes (ΔCt), and calculated as 2−ΔCt. Expression levels were used for descriptive and statistical analyses. Relative expression was calculated and expressed as 2−ΔΔCt log2-fold change (Livak and Schmittgen, 2001).
Flow cytometric analysis
Cells were measured and analyzed on a FACScan flow cytometer (BD, Heidelberg, Germany), with 488 nm air-cooled argon ion laser. At least 10,000 cells were collected and evaluated. Three samples were analyzed simultaneously, and the average was used for statistical analyses. The proportions of individual subpopulations were expressed as relative percentages.
Statistical analysis
A principal component analysis (PCA) was carried out as a tool of exploratory data analysis. Significant differences between the evaluated groups were assessed by a one-way analysis of variance and confirmed using Tukey post hoc test. Relationships between the various indicators were determined using Pearson's r correlation coefficient. These analyses were carried out with MATLAB® version 9.9.9341360 (R2016a). The heatmap and biplot were rendered with Python's plotting library, Matplotlib 3.0.3. (Supplementary Dataset S3).
Results
Effects of L. fermentum and C. coli exposure
PCA was carried out to study the differences and correlations between the evaluated inflammatory cytokines (Fig. 1). The outcomes show that expression of inflammatory cytokines in the control explained most of the variation for component 1, whereas the experimental groups did so for component 2. Exposure to any of the bacterial treatments triggered the downregulation of inflammation-related genes. IL-15 was the highest in the control group; nonetheless, expression of this interleukin was significantly reduced after L. fermentum exposure (Fig. 2, Table 2, and Supplementary Table S1 and Supplementary Dataset S1). Similarly, IL-16 and IL-13RA1 abundance was reduced after exposure to the probiotic, whereas infection with C. coli did not modify significantly the abundance of such transcripts (Fig. 2 and Table 2).
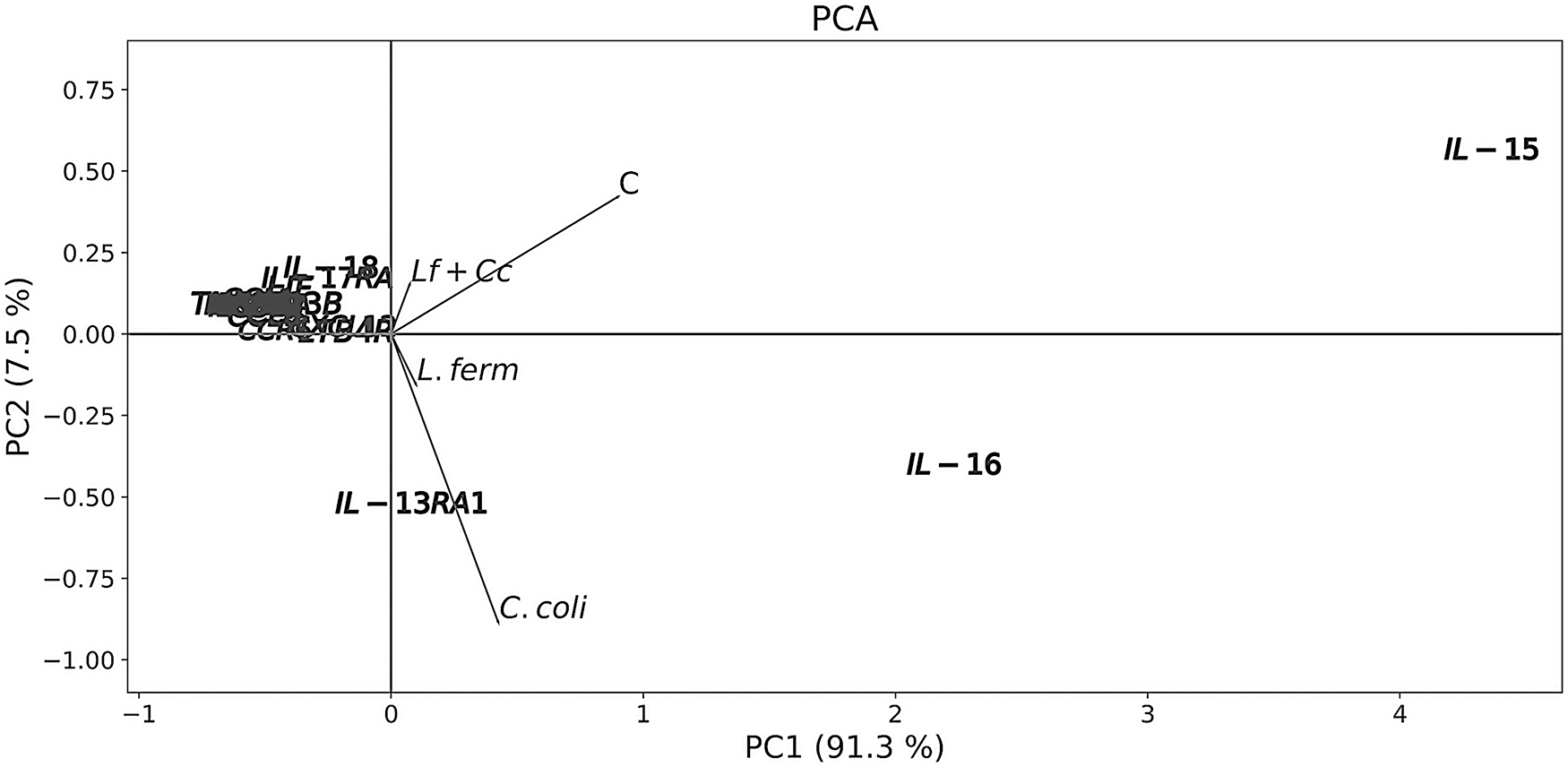
Ordination by PCA of cecum-based cytokines of broiler chickens exposed to Lactobacillus fermentum and/or Campylobacter coli. PCA, principal component analysis.
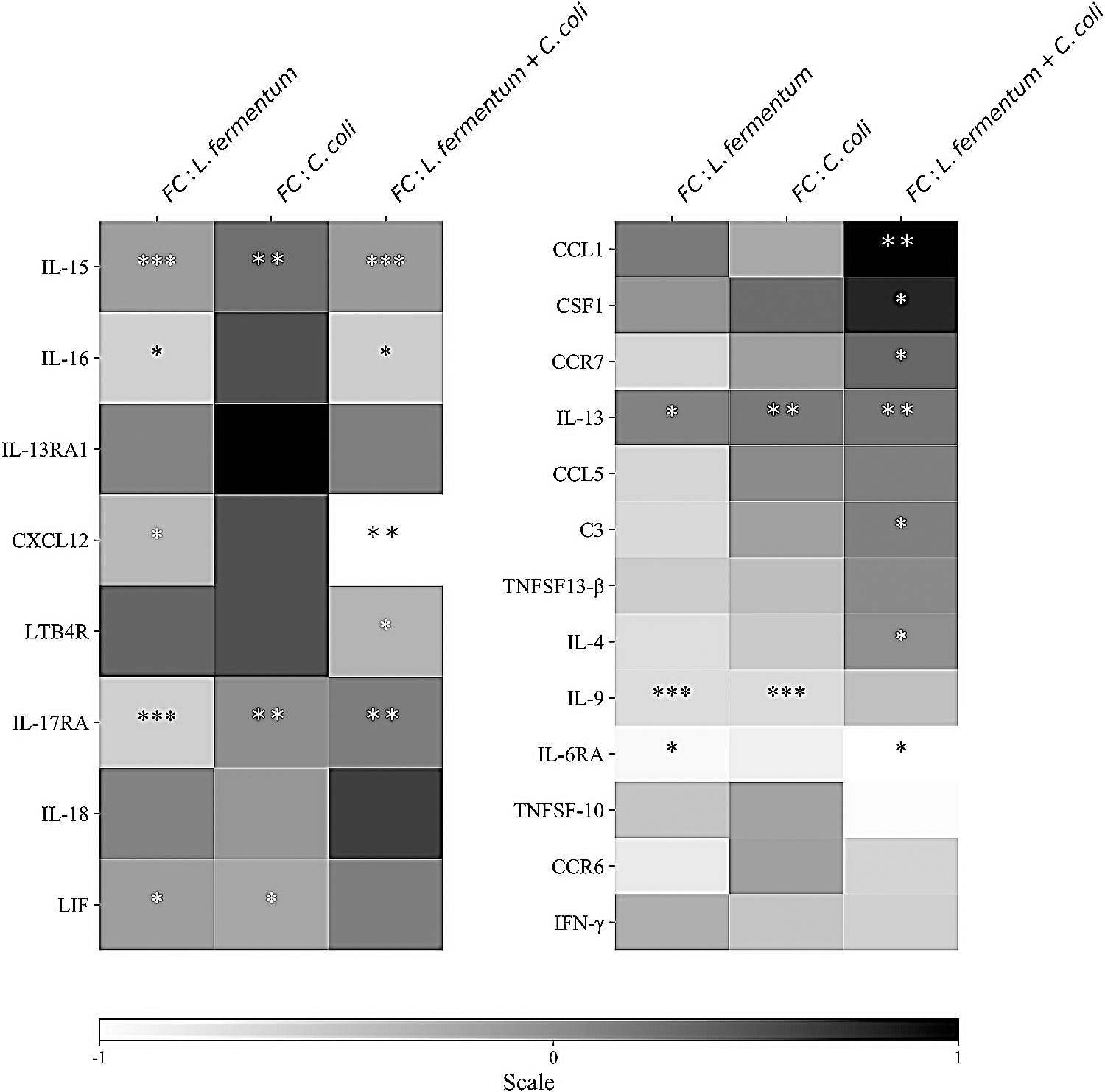
Heat map showing the transcriptional fold change of inflammatory cytokines in the cecum of broiler chickens after administration of the probiotic L. fermentum and exposure to the pathogen C. coli, either alone or in combination. The color scale, −1 (white) to +1 (black), indicates mRNA expression levels relative to the values of the control group, with white designating downregulated genes and black indicating upregulated genes. FC, fold change. *Significant differences with the control group (*p < 0.05; **p < 0.01;***p < 0.001).
mRNA Expression Levels of Cecal Inflammatory Cytokines in Probiotic and Pathogen-Treated Broiler Chickens
Values are represented as mean ± SD.
Significant differences with the control group ( * p < 0.05; ** p < 0.01; *** p < 0.001).
Significant differences with the L. fermentum treatment (▪ p < 0.05; ▪▪ p < 0.01; ▪▪▪ p < 0.001).
Significant differences with the C. coli treatment (▴ p < 0.05; ▴▴ p < 0.01; ▴▴▴ p < 0.001).
IL, interleukin; IFN-γ, interferon-γ; TNF, tumor necrosis factor.
Some cytokines were observed to be clustered in groups, depending on their contribution to the accounted variability of the components (Fig. 1 and Table 2). In group 1 (CXCL12 and LTB4R), transcript abundance was the highest in the control, being reduced by inoculation with L. fermentum (Fig. 2, Table 2, and Supplementary Table S1). Alternatively, transcript abundance of LIF, IL-18, and IL-17RA (second group) was downregulated at least 2-fold, except for IL-18 whose levels were as high as the ones in the control (Fig. 2, Table 2, and Supplementary Table S1). Expression levels of some cytokines belonging to the third group proved to be below the mean for component 1 (values were lower than those of group 2) and, in general, over the mean for component 2 (Fig. 1), which appears to be linked to the general rise in transcript abundance in the L. fermentum + C. coli group (Fig. 2, Table 2, and Supplementary Table S1).
Effects on lymphocyte levels
Intraepithelial and lamina propria-based lymphocytes were determined using flow cytometry. The percentage of positive intraepithelial CD8 lymphocytes (IELs) was significantly greater in all experimental groups. On the contrary, the percentage of positive CD8 in the lamina propria increased significantly only when chickens were infected with the pathogen after previous exposure to the probiotic (Fig. 3 and Supplementary Dataset S2). In this group, the percentage of positive IgA and IgM plasma cells in the lamina propria was greater compared with the control. In addition, the percentage of IgM cells was higher than that of the control when L. fermentum and C. coli were administered separately. Differences in the percentages of intraepithelial IgA were also found between the L. fermentum + C. coli and the other groups (Fig. 3). Furthermore, percentages of cells positive to CD3, CD4, and CD45 markers, in both the epithelium and lamina propria, did not change significantly in any of the treatments (Fig. 3).

Flow cytometric analysis. Percentage ± SD of lymphocyte subsets in samples from the lamina propria and epithelium of L. fermentum and C. coli-challenged broiler chickens. *Significant differences with the control group (*p < 0.05; **p < 0.01;***p < 0.001). ▪Significant differences with the L. fermentum treatment (▪ p < 0.05; ▪▪ p < 0.01; ▪▪▪ p < 0.001); ▴Significant differences with the C. coli treatment (▴ p < 0.05; ▴▴ p < 0.01; ▴▴▴ p < 0.001).
Correlation analysis between cecal inflammation and immune indicators
Pearson's r correlation coefficient analysis (Fig. 4 and Supplementary Table S2) revealed positive correlations among the subpopulation of lymphocytes. IgM plasma cells from lamina propria proved to be correlated to intraepithelial IgA, and to both lamina propria and intraepithelial CD8 lymphocytes. Likewise, CD8 cells from both locations demonstrated to be positively correlated. Positive correlations were additionally found between different cytokines and percentages of intraepithelial and lamina propria-based IgA (Fig. 4 and Supplementary Table S2). Similarly, IgM levels in the lamina propria proved to be positively correlated with levels of IL-13. Conversely, negative correlations were observed between lamina propria, intraepithelial CD8 cells, and LTBR4. Moreover, the data also revealed significant correlations between some cytokines (Fig. 4 and Supplementary Table S2).
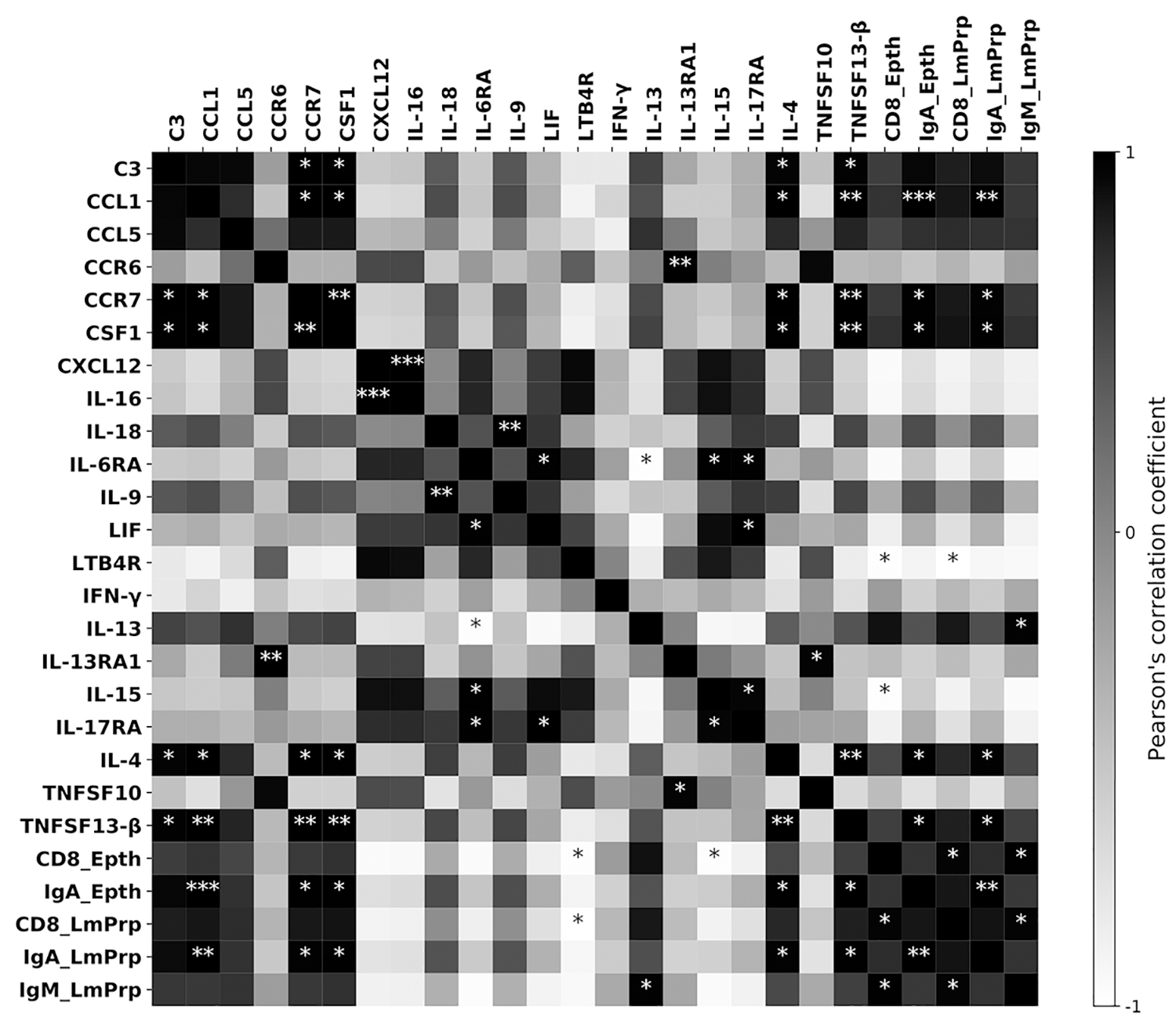
Heat map based on the Pearson's r correlation coefficient matrix among the indicators examined. The r scores are denoted in grayscale from −1 (white) to 1 (black). *Significant correlations (*p < 0.05; **p < 0.01; ***p < 0.001).
Discussion
L. fermentum has been shown to lessen pathogenic inflammatory reactions and modulate the expression of key immune mediators (Zhao et al., 2019). Specifically, its use in avian models has proved to ameliorate inflammation and immune response after pathogen exposure (Guo et al., 2015). In this study, our main goal was to assess the way in which L. fermentum could modulate the cytokine-mediated inflammatory reaction in the cecum of broiler chickens infected with C. coli. We examined the expression levels of a group of cytokines involved in an inflammatory response, and sought to determine the effects on immune cell subpopulations.
In this study, treatments making use of L. fermentum proved to reduce the abundance of inflammation-related transcripts, corroborating the positive influence of probiotics (Penha Filho et al., 2015). Moreover, this research shows that previous inoculation with L. fermentum, before infection with C. coli, elicited a complex inflammatory response. Such outcomes have been hitherto reported. In fact, L. fermentum AD1 administration was shown to modulate mRNA abundance of inflammatory mediators in C. jejuni-challenged chickens (Stašová et al., 2015).
Earlier studies have demonstrated that C. coli modulates expression of proinflammatory and regulatory cytokines in human and murine cells ( Klančnik et al., 2015; Hamza et al., 2017). We observed that infection with C. coli significantly reduced the expression of five cytokines, along with the upregulation of IL-13. These outcomes demonstrate that the reaction to C. coli appears less striking compared with the one caused by C. jejuni, which triggers a marked inflammatory reaction (Connerton et al., 2018).
L. fermentum administration increases levels of antibodies such as IgA and IgM (Rocha et al., 2012;Salim et al., 2013). In comparison, we found that levels of IgA were significantly increased in the lamina propria and epithelium when both bacteria were administered. The percentage of lamina propria IgM plasma cells proved to be significantly increased by all treatments. Other studies have demonstrated that administration of Lactobacillus strains modulates levels of IgM and IgG (Mountzouris et al., 2010). Our observations suggest that infection with C. coli in the presence or absence of the probiotic induces an increased humoral response. In particular, the percentage of IgA was observed to be augmented only when infection proceeded inoculation with L. fermentum. IgA is known to play a key role in protecting gut mucosa by neutralizing or preventing toxins, viruses, or bacteria from binding the mucosal surface. Moreover, they are able to promote pathogen phagocytosis and opsonization, which involves bacterial agglutination and complement activation (Havelaar et al., 2009). This response seems to be linked to the differential expression of various type II cytokines observed in this group. Microorganism-associated molecular patterns (lipoteichoic acid, wall teichoic acid, and peptidoglycan) of Lactobacillus species are known to activate receptors in epithelial and dendritic cells. These Toll-like and NLRP receptors activate downstream signaling, which eventually induces cytokine production that promotes the polarization of Th2 cells and their characteristic cytokine production (Bron et al., 2012).
Inoculation with C. coli induced an increased presence of IELs, which was no different from the effects elicited by L. fermentum. This response seems mild in comparison with the one prompted by C. jejuni. Several reports have indicated a significant increase of CD8, CD4, and various immunoglobulins ( Pielsticker et al., 2016; Han et al., 2017); in fact, this reaction pattern has supported the notion that C. jejuni does not play a commensal role in chickens at all. The percentage of IEL cells was significantly greater in all experimental groups than in control conditions. Nonetheless, levels of lamina propria CD8 cells were significantly greater only when chickens were inoculated with C. coli in the presence of L. fermentum. These results corroborate previous findings where higher percentages of these T cells have been detected after administration of various Lactobacillus strains (Bai et al., 2013). However, the percentages of CD4 found in these experiments were significantly augmented, as opposed to what is reported herein.
IL-15 proved to be negatively correlated to IELs; IL-15 has been seen to elicit important CD8 responses, including their generation and maintenance (Lim et al., 2012). In fact, tissue-resident (induced and natural) IELs, along with effector memory T cells (TEM), rely on IL-15 production (Konjar et al., 2017). Nonetheless, high levels of this cytokine have been observed to induce differentiation of these semi-activated cells into cytolytic factors-releasing effector T cells (Itsumi et al., 2009). This suggests that the 3-fold reduction in IL-15 expression might be required for avoiding polarization of IELs and TEM into effector cytotoxic cells. Despite this downregulation, IL-15 was the most abundant cytokine in all experimental treatments; commensal bacteria are known to regulate IL-15 production by signaling involving epithelial-based TLR2 and MyD88 (Konjar et al., 2017). Proinflammatory cytokines (IL-16, IFN-γ, and TNF), typically released by effector T cells, were observed to be downregulated by the presence of the lactic acid bacteria, which insinuates that the detected percentages of CD8 cells might be related to induced IEL and TEM cells, and not to cytotoxic T cells. The detected CD8 IELs may contribute to the overall pathogen containment and clearance. Although these IELs are not effector cells, they can produce a range of antimicrobial and tissue repair factors, which seem crucial for mucosal protection. In addition, IELs can migrate to sites of infection, where they interact with other IELs; such costimulation promotes a collective release of antimicrobial peptides (Ganz, 2003). Proliferation of these cells may not require antigen processing, but instead could rely on the recognition of molecular patterns (peptide and nonpeptide) released by bacteria or epithelial cells upon damage or stress (Tanaka et al., 1995).
Some genes were clustered in groups; group 1 included genes downregulated after exposure to treatments using L. fermentum. LTB4R, member of the TNF superfamily, proved to be negatively correlated with CD8, despite the fact that this factor positively influences the accumulation of CD8 cells in the presence of a pathogen (Sun et al., 2016). Similarly, cytokines from the second group (IL-17RA and IL-18) were notably downregulated after exposure to L. fermentum and C. coli; this reinforces the notion that the observed CD8 cells are in fact IEL and TEM cells and not effector cells, whose development is characteristically influenced by the aforementioned cytokines (Okamoto et al., 1999; Ng et al., 2015; Boari et al., 2018).
Cytokines of group 3 were mostly upregulated when colonization by C. coli occurred in the presence of L. fermentum; in particular, differential expression of Th2 cytokines (IL-13, IL-4, and IL-9) was detected. IL-13 and IL-4, for example, proved to be positively correlated to IgM and IgA, respectively. These cytokines are known to control inflammation mechanisms and activate humoral immunity. In fact, IgM and IgA levels have been observed to be modulated by expression of these factors (Al-Banna et al., 2018). In addition, the chemokines CCL1 and CCR7, along with the TNFSF13-β, were upregulated. These factors play important roles in maintaining gut homeostasis, which involves recruitment and maturation of dendritic and antigen-presenting cells. Deficiencies in CCR7 signaling are related to delays in production of IgA after Salmonella infection in mice (Hashizume-Takizawa et al., 2019). Likewise, TNFSF13-β has been observed to regulate immunoglobulin production in the intestine (Hashiguchi et al., 2018). Our results corroborate previous findings by demonstrating that the upregulation of these factors proved to be linked to the observed percentages of IgA in both epithelium and lamina propria. Moreover, upregulation of CCL5 and CSF-1 might suggest monocyte recruitment, which could represent a source of intestinal macrophages (Nieto et al., 2017). C3 levels were similarly increased, implying a potential role of the complement (Sünderhauf et al., 2017). Arguably, L. fermentum-induced pathogen phagocytosis, along with opsonization of mucosa-attached bacteria, seems to play a role in containing invasive C. coli.
Conclusion
Inoculation with probiotics has been widely used as a method for reducing gut colonization by unwanted microorganisms. In this study, we demonstrate that infection with C. coli, in the presence of L. fermentum, modulates the expression of inflammatory cytokines in the cecum of broiler chickens, which leads to a heightened humoral response that seems to be linked to an increased abundance in type II cytokines. Furthermore, downregulation of proinflammatory cytokines suggests that the detected CD8 cells might not yet be polarized into effector cells, limiting, therefore, the onset of inflammatory processes. In conclusion, these outcomes provide critical insights into the current understanding of the immunomodulatory effects of utilizing Lactobacillus strains for eluding colonization by Campylobacter spp. in broiler chickens.
Footnotes
Acknowledgments
The authors thank Radomíra Nemcová, Assoc. Prof. DVM, PhD, head of the Laboratory of Gnotobiology, and Jana Koščová, DVM, PhD, head of the Department of Microbiology and Immunology, for helping with the bacterial strain preparation.
Author's Contributions
M.S.: conceptualization, methodology, supervision, writing—original draft preparation; M.L.-Á.: conceptualization, methodology, writing—original draft preparation; C.L.-Á.: formal analysis, methodology, software; V.K.: RT-qPCR, funding acquisition; V.R.: methodology, flow cytometry, original draft preparation; S.G.: conceptualization; Z.Š.: conceptualization; R.H.: conceptualization, methodology, supervision.
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the Grant Agency for Science of the Slovak Republik VEGA (1/0112/18, 1/0355/19); Slovak Research and Developmental Agency (APVV-15-0165).
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Dataset S1
Supplementary Dataset S2
Supplementary Dataset S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.