
Abstract
An Escherichia coli strain isolated from the feces of swine at a pork slaughterhouse in Henan province China was found to possess two colistin-resistance genes, mcr-1 and mcr-3, plus 16 additional resistance genes. Genes mcr-1.1 and mcr-3.1 were identified on IncHI2 and IncX1 type plasmids, respectively. Transconjugants (containing mcr-3, mcr-1&mcr-3) were obtained that were 64- and 512-fold higher than the minimum inhibitory concentration of colistin on the recipient bacteria (E. coli C600), respectively. The IncX1 plasmid containing mcr-3.1 displayed a very specific structure compared with previous mcr-3. Variable and stable regions were similar across different plasmids, multiple insertion sequences and transposases.
Polymyxin is a polypeptide antibiotic extracted from bacteria that is classified as polymyxin B or polymyxin E (colistin) and has bactericidal effect against almost all Gram-negative bacilli (Li et al., 2005). Polymyxin has been widely used in China as an antibiotic, since the approval of a colistin sulfate premix from Japan as feed additive in 1997, and as a veterinary drug since 2002.
However, the extensive use of colistin in China and worldwide has led to the continuous emergence of Gram-negative pathogens with colistin and multidrug resistance. As a result, colistin is at risk of losing its status as “the last line of defense” against drug-resistant bacteria (Giamarellou, 2016). In 2015, Chinese scholars identified mcr-1 as a new drug-resistance gene (Liu et al., 2016). As more colistin-resistance genes were identified, the Chinese Ministry of Agriculture issued a notice to officially terminate the use of colistin as a feed additive on July 26, 2016.
Mobile genetic elements such as plasmids, transposons, integrons and insertion sequence elements play essential roles in the dissemination of antibiotic resistance in Gram-negative bacteria. In particular, plasmids can self-replicate and act as vectors for other mobile elements with antibiotic-resistance genes, including the mcr-1 and mcr-3 genes identified in a large-scale screen of polymyxin-resistant genes in China (Yin et al., 2017), which are also present on diverse Enterobacteriaceae plasmids, including IncI2, IncX4, IncHI1, IncHI2, IncFI, IncFII, IncP and IncK (Wang et al., 2017).
Multidrug-resistant strains harboring mcr-3 were first identified in 2017, when mcr-1 and mcr-3, along with the carbapenemase gene bla NDM-5, were found in an Escherichia coli clinical strain (WCHEC020123) belonging to phylogenetic group A and sequence type 206 (ST206) (Liu et al., 2017). A second independent case of the coexistence of mcr-1, mcr-3, and bla NDM-5 was reported for E. coli strain WCHEC025943 (ST410) recovered from hospital sewage (Long et al., 2019). This strain showed an even broader antimicrobial resistance spectrum than the multidrug-resistant strain WCHEC020123. In this study, we report another multidrug-resistant E. coli strain co-expressing mcr-1, mcr-3, and bla CTX-M-14 isolated from the feces of swine at a pork slaughterhouse in Henan province, China, in May 2017.
The isolate (YZUC2624) was screened for the presence of mcr-1 and mcr-3 by polymerase chain reaction using the primers described by Liu et al. (2016) and Yin et al. (2017), and it was found to belong to ST77. The microbroth dilution method (in triplicate) was used to determine the susceptibility of the strain to various antibiotics according to the Clinical and Laboratory Standards Institute (CLSI) guidelines and breakpoints (M100-S25), veterinary CLSI (VET01-A4/VET01-S2), and European Committee on Antimicrobial Susceptibility Testing (EUCAST) (Chen et al., 2013; Desai et al., 2015; Liu et al., 2015). In addition to the colistin resistance conferred by mcr-1 and mcr-3 (minimum inhibitory concentration, MIC = 64 mg/L), YZUC2624 was resistant to nine classes of drugs (Supplementary Table S1), demonstrating an even broader antimicrobial resistance spectrum than strains WCHEC020123 and WCHEC025943.
Conjugation experiments were conducted using streptomycin-resistant E. coli C600 as the recipient. The transconjugants were selected on MacConkey agar plates supplemented with streptomycin (128 mg/L) and colistin (4 mg/L), revealing that the mcr-3 gene in YZUC2624 was transferable. The E. coli C600-pYZUC2624.2 transconjugant harbored only mcr-3 with frequencies of 5 × 10−5–5 × 10–6/donor (Fig. 1), and was resistant to six classes of drugs, whereas the E. coli C600-pYZUC2624.1&2 transconjugant harbored both mcr-1 and mcr-3 with frequencies of 4 × 10–3/donor, and was resistant to eight classes of drugs, with 64- and 512-times higher MICs than that of the parental strain E. coli C600, respectively (Supplementary Table S1).
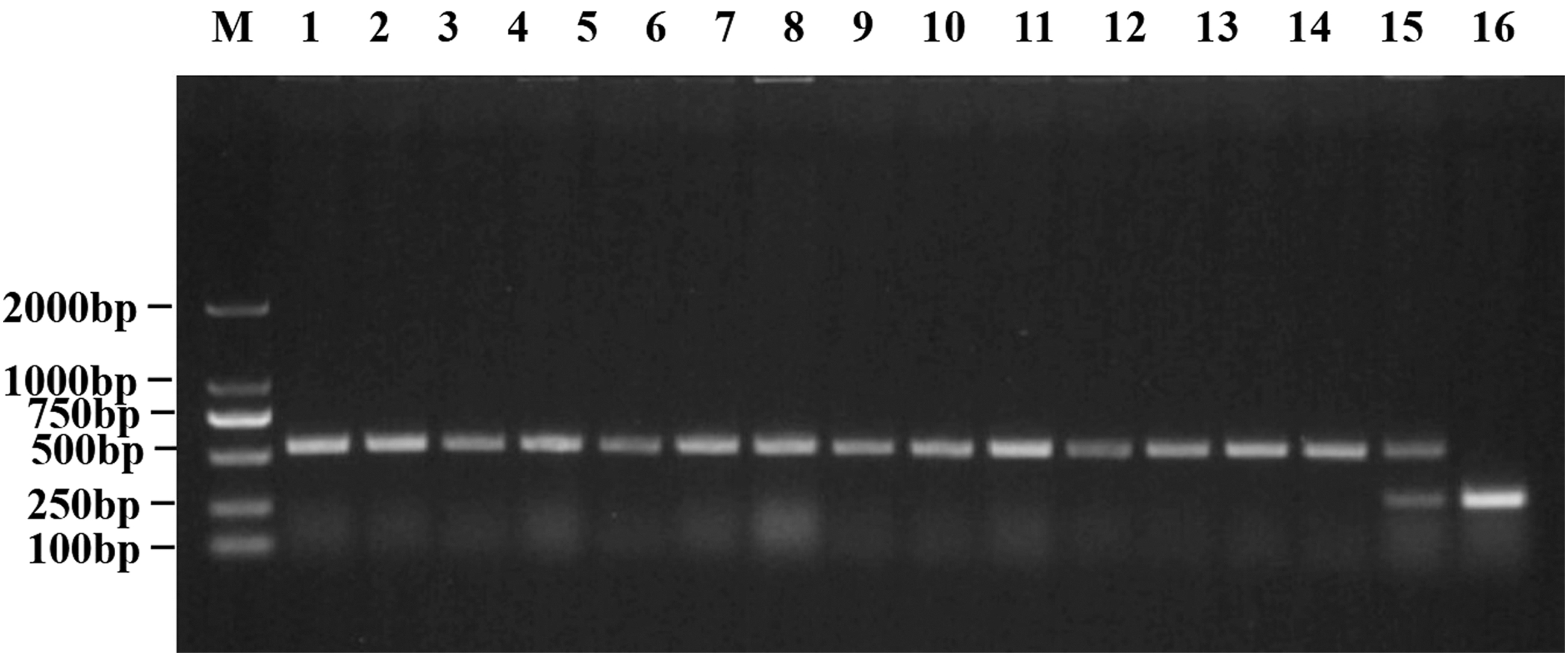
The plasmids carrying mcr-1 and mcr-3 were transferred to Escherichia coli C600 recipient by conjugation. Polymerase chain reaction was used to detect the carrying of mcr-1 and mcr-3 genes in the transconjugants. The results of agarose gel electrophoresis are shown in the figure. Lane M: DL2000, lane 1–14: transconjugants harbored only mcr-3 (542 bp), lane 15: transconjugant harbored both mcr-1 and mcr-3, lane 16: the positive control of mcr-1 (309 bp).
Total genomic DNA from E. coli YZUC2624 was extracted using a TIANGEN® TIANamp Bacteria DNA Kit (Tiangen Biotech, Beijing) according to the protocol recommended by the manufacturer. A 300-bp library for Illumina paired-end sequencing was constructed and was immediately sequenced using a MiSeq platform. To obtain the complete sequence of pYZUC2624.1 and pYZUC2624.2, the PacBio RSII single-molecule real-time (SMRT) sequencing strategy was adopted. The mcr-1-harbouring plasmid and mcr-3-harbouring plasmid was extracted from the transconjugant E. coli C600-pYZUC2624.2 and E. coli C600-pYZUC2624.1&2 using a TIANprep Mini Plasmid Kit (TIANGEN).
Template libraries were prepared using a SMRTbell Template Preparation Kit (Pacific Biosciences, Menlo Park, CA) and sequencing was performed using a PacBio RSII instrument (Pacific Biosciences). The raw sequence data were assembled using RS Hierarchical Genome Assembly Process protocol version 3.0 in SMAT Analysis version 2.3.0 (Pacific Biosciences) (Stoesser et al., 2014). Gene prediction and annotation were performed using RAST software tools (Aziz et al., 2008). Resistance-related genes, plasmid replicon types, and sequence types (STs) were analyzed using ResFinder 2.1, PlasmidFinder 1.3, and MLST 1.8* multilocus sequence typing servers, respectively, which are available at the Center for Genomic Epidemiology at the Danish Council for Strategic Research (
DNA sequencing, de novo assembly, and annotation identified 18 resistance genes distributed across three plasmids (Supplementary Table S2): IncHI2-type plasmid pYZUC2624.1 (225,059 bp; GenBank accession no. MK962305), containing four resistance genes (mcr-1.1, bla CTX-M-14, fosA3, and sul1); IncX1-type plasmid pYZUC2624.2 (45,354-bp; GenBank accession no. MK962306), containing eight resistance genes [Fig. 2: mcr-3.1, aadA1, aadA2, bla TEM-1B, cmlA1, floR, qnrS1, and tet(A)]; and IncR1-type plasmid pYZUC2624.3, with 76,622 bp containing six resistance genes [aac(3)-IId, aadA2, Inu(F), sul3, tet(A), and dfrA12]. Comparison of pYZUC2624.1 with eight strains harboring mcr-1-carrying IncHI2 plasmids using BRIG30 (Fig. 3) showed sequence coverage of 80–88%, which was highest in the backbone structure region, and significant differences in the regions harboring drug-resistance genes.

Plasmid map of plasmid pYZUC2624.2 (GenBank accession no. MK962306). The innermost circle is GC content (52% GC), the second circle is GC skew, the third circle of arrows represent the position and transcriptional direction of the open reading frames. Resistance genes are indicated by fuchsia arrows. The remaining sequence segments are indicated by red arrows. GC, percentage of guanine and cytosine. Color images are available online.
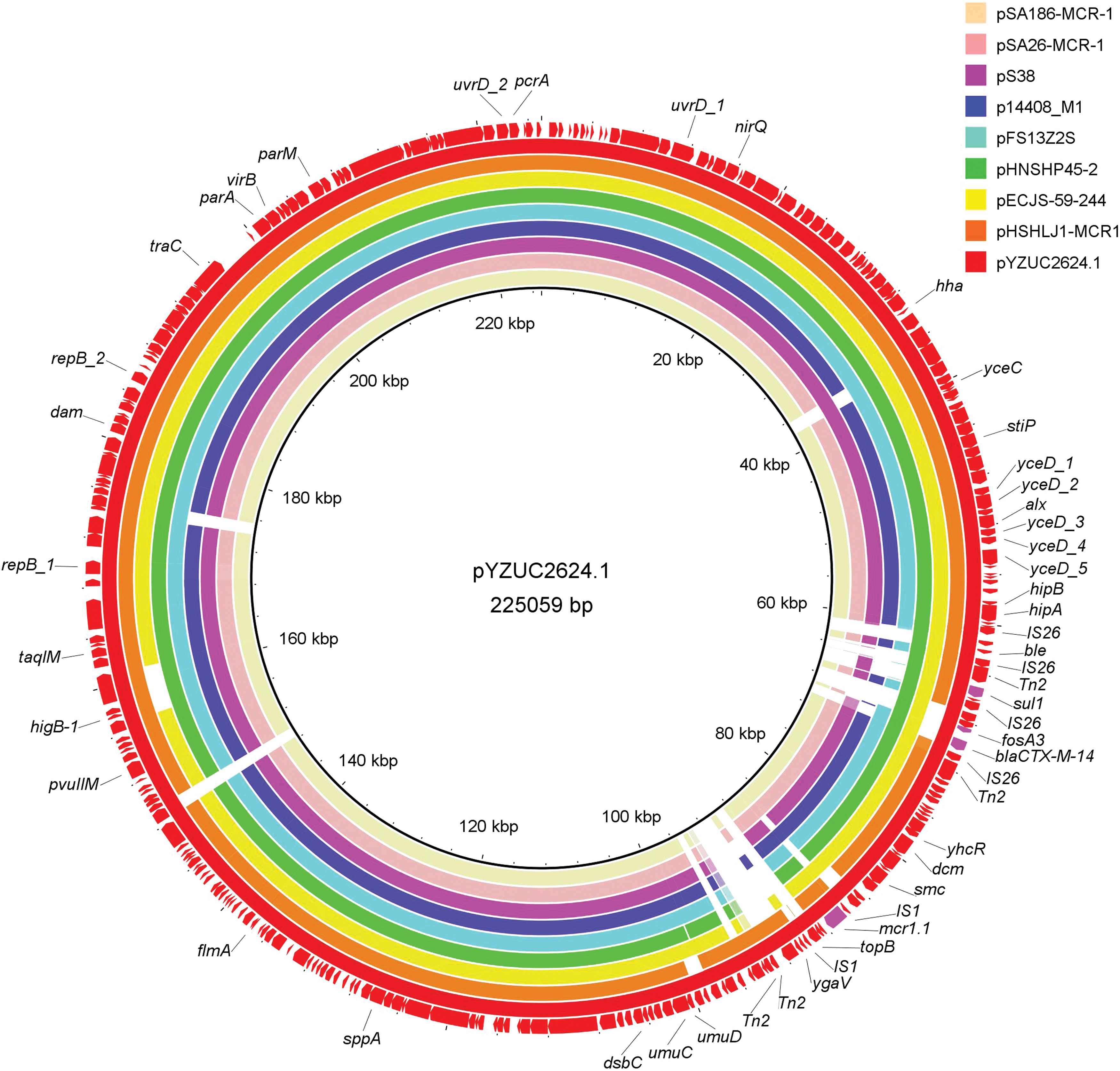
Alignment of mcr-1-carrying IncHI2 plasmids. Alignment was performed using BRIG30, pYZUC2624.1 (GenBank accession no. MK962305) was used as the reference. GenBank accession no. of these plasmids are CP022735, KU743384, KX129782, LT599829, KY421937, KU341381, KX084394, KX856066, and MK962305, respectively. Outer circle arrow represents the position and transcriptional direction of the open reading frames in pYZUC2624.1. Resistance genes are indicated by fuchsia arrows. The remaining sequence segments are indicated by red arrows. Color images are available online.
Linear comparisons of pYZUC2624.2 (no. MK962306) with other plasmids (Fig. 4) showed that the genetic environment of mcr-3.1 is similar to the mcr-3.22 environment of pUnnamed3 (Klebsiella, USA, CP041095), which has two additional insertion sequence (IS3 and IS26) compared with pYZUC2624.2. The environment of the other seven resistance genes on pYZUC2624.2 was most similar to that of pQnrS1_005237 (E. coli, China, CP026578), despite multiple misalignments. The IncX1 plasmid structure of pYZUC2624.2 was similar to that of pHYEC7–110 (E. coli, Hong Kong, China, KX518744).
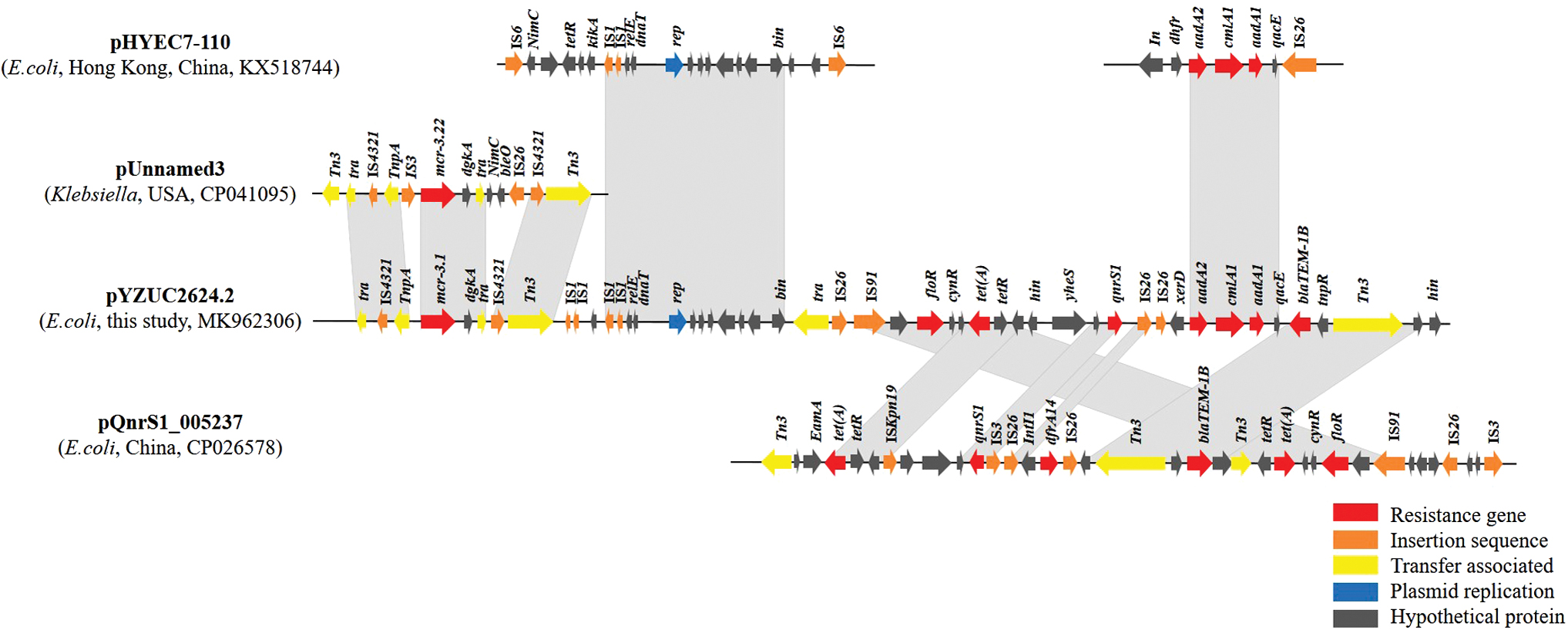
Linear comparisons of pYZUC2624.2 (GenBank accession no. MK962306) with other plasmids. Regions >99% identity are shaded in gray. Solid lines indicate the plasmid backbone. The extents and directions of antibiotic-resistance genes and other genes are indicated. Resistance genes are in red, insertion sequences are in orange, transfer-related proteins are in yellow, replication proteins are in blue, and hypothetical proteins are in black. Color images are available online.
The co-expression of multiple clinically important antimicrobial resistance genes presents a particular challenge for clinical treatment and infection control in health care settings. Notably, plasmid pYZUC2624.2 carrying mcr-3 is of the IncX1 type, whereas mcr-3 was found on IncP-type plasmids in the previous reports of multidrug-resistant E. coli carrying mcr-1 and mcr-3 (Liu et al., 2017; Long et al., 2019). This suggests the possible emergence of super-resistant strains, and the ability of horizontal transfer of mcr-3-containing plasmids can further increase their spread.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the National Key Research and Development Program of China (grant 2017YFD0500102), Jiangsu Training Program for College Students to Innovate and Start Enterprise (grant 201811117094X), Six Talent Peaks Project (SWYY-083), and the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD).
Supplementary Material
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.