
Abstract
Retail meats are one of the main routes for spreading antimicrobial-resistant bacteria (ARB) from livestock to humans through the food chain. In African countries, retail meats are often sold at roadside butcheries without chilling or refrigeration. Retail meats in those butcheries are suspected to be contaminated by ARB, but it was not clear. In this study, we tested for the presence of antimicrobial-resistant Escherichia coli from retail meats (n = 64) from roadside butcheries in Kampala, Uganda. The meat surfaces were swabbed and inoculated on PetriFilm SEC agar to isolate E. coli. We successfully isolated E. coli from 90.6% of these retail meat samples. We identified the phylogenetic type, antimicrobial susceptibility, and antimicrobial resistance genes prevalence between retail meat isolates (n = 89). Phylogenetic type B1 was identified from 70.8% of the retail meat isolates, suggesting that the isolates originated primarily from fecal contamination during meat processing. Tetracycline (TET)-resistant isolates with tetA and/or tetB gene(s) were the most frequently detected (28.1%), followed by ampicillin (AMP) resistance genes with bla TEM (15.7%,) and sulfamethoxazole–trimethoprim (SXT) resistance genes with sul2 (15.7%). No extended-spectrum beta-lactamase-producing isolates were detected. A conjugation assay showed that resistance to AMP, TET, and SXT could be simultaneously transferred to recipients. These findings suggest that antimicrobial-resistant E. coli can easily be transferred from farms to tables from retail meats obtained from roadside butcheries.
Introduction
The spread of antimicrobial-resistant bacteria (ARB) is a global concern in both human and veterinary medicine (World Health Organization, 2015). ARB in food-producing animals presents a risk of disseminating ARB from animals to humans through the food chain (Economou and Gousia, 2015; Founou et al., 2016). As fecal bacteria in livestock can easily be transferred to retail meats in a slaughterhouse during meat processing, retail meats are one of the most important routes for spreading ARB from livestock to humans, or “farm-to-table” (Aslam et al., 2003; Schroeder et al., 2004). Many researchers have reported the detection and prevalence of ARB in retail meat samples (Johnson et al., 2009; Eyi and Arslan, 2012; Zhao et al., 2012; Martínez-Vázquez et al., 2018). Some studies have reported isolating clinically important ARB, such as extended-spectrum beta-lactamase (ESBL)-producing bacteria, from retail meat samples (Ye et al., 2018). However, most of these studies were conducted in developed countries with improved food chains and appropriate cooling systems, and ARB studies of retail meats from developing countries remain limited (Messele et al., 2017).
Retail meats in developed countries are usually distributed in cold chains (Nastasijević et al., 2017). In general, beef carcasses are chilled immediately after slaughtering in chilled rooms for 24–96 h. They are kept in chilling rooms and then cut into primary cuts for distribution. Some studies reported that the numbers of lactic acid bacteria and Pseudomonas on the chilled meat surfaces increased, whereas the number of Enterobacteriaceae (Salmonella Enteritidis) slightly decreased (Ercolini et al., 2006; Chenoll et al., 2007; Sabike et al., 2015). Conversely, roadside butcheries in developing countries purchase their retail meats from slaughterhouses in the morning and sell them on the same day without chilling because these butcheries are rarely equipped with a refrigerator or freezer (Fig. 1). In these countries, retail meats are commonly transported directly from the slaughterhouses to retailers on bicycles or light trucks. Thus, we considered that retail meats from roadside butcheries clearly reflect the prevalence of ARB among livestock in Uganda. To demonstrate the possibility of spreading ARB through retail meats from roadside butcheries, here we report the prevalence and the characteristics (phylogenetic type, antimicrobial susceptibility, and resistance genes) of antimicrobial-resistant Escherichia coli, a major fecal indicator bacterium, isolated from retail meats in Uganda (Schroeder et al., 2004; Johnson et al., 2009).
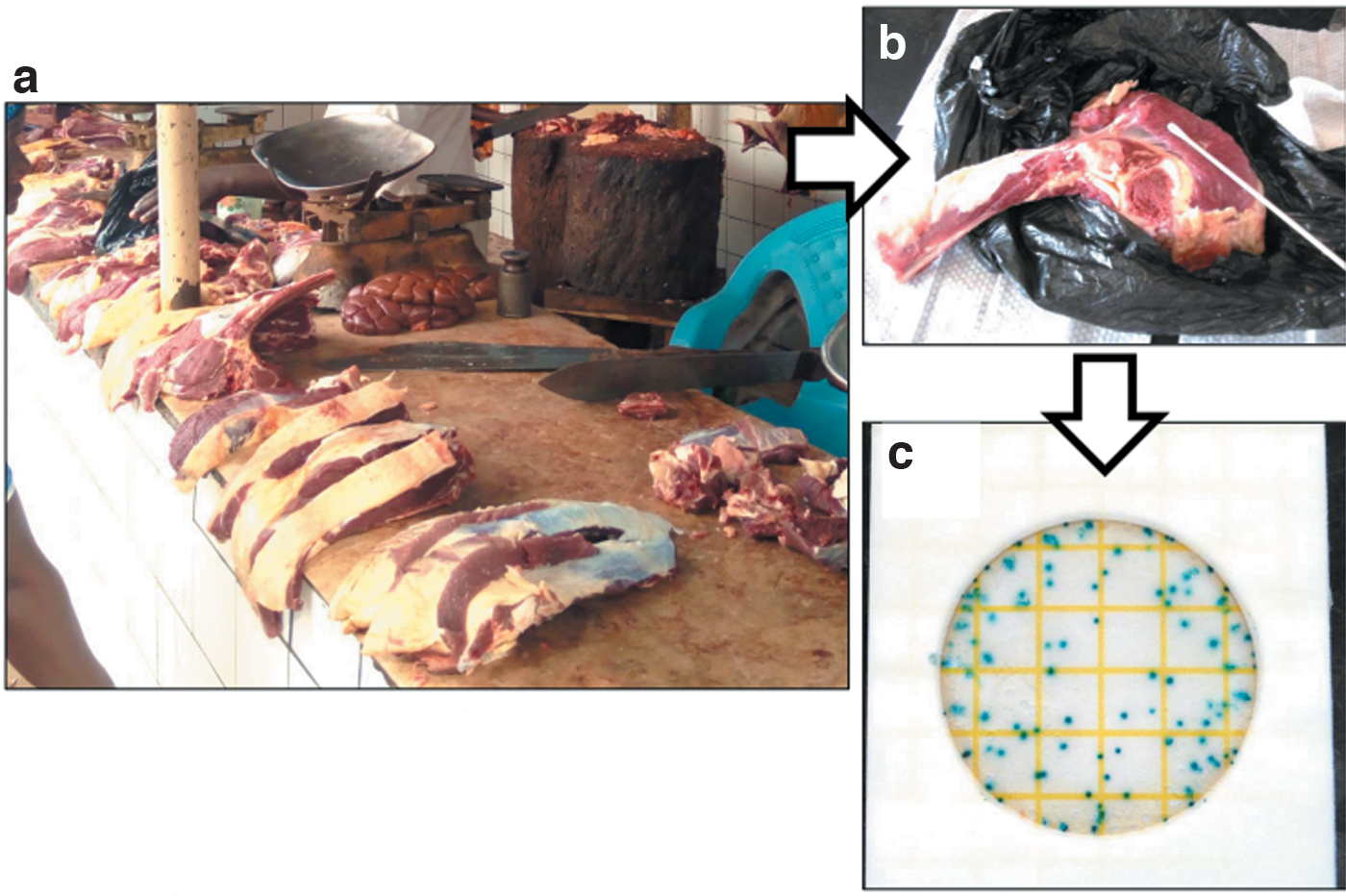
Sampling and processing of retail meat samples. Retail meat samples were purchased from roadside butcheries
Materials and Methods
Between February and October 2018, 64 retail meat samples (60 beef ribs and 4 goat ribs) were purchased at roadside butcheries (n = 64) in Kampala, Uganda (Fig. 1). One piece (quarter or half kilogram) was bought at each of the butcheries. Retail meats in the roadside butcheries were slaughtered on the same day at slaughterhouses in Kampala. We could not trace which slaughterhouse had processed the meat we bought. The samples were transported to the laboratory at ambient temperature and processed within 3 h after collection. The meat sample surfaces (∼30 × 10 cm) were swabbed completely with sterilized cotton swabs moistened by sterilized 0.85% saline solution (Nissui Pharmaceutical Co., Ltd., Tokyo, Japan) and suspended in 1 mL of saline solution. After serial dilution, 1 mL of the solution was inoculated on a PetriFilm SEC plate (3M Company, MN) and cultured at 37°C overnight. Up to two representative blue colonies (presence of β-galactosidase) were subcultured on ES Colimark agar media (Eiken Chemical Co., Ltd., Tokyo, Japan). Single blue colonies (presence of β-glucuronidase) were defined as E. coli isolates and were checked through E. coli-specific polymerase chain reaction (PCR) (Wang et al., 1996).
PCR-based phylogenetic typing of E. coli was performed to classify the isolates into seven groups and subgroups: A0, A1, B1, B22, B23, D1, and D2 (Escobar-Páramo et al., 2004). E. coli belonging to groups A0, A1, and B1 are usually commensal and nonpathogenic strains. Groups B22 and B23 include extra-intestinal pathogenic strains related to urinary tract infections, and groups D1 and D2 include intra-intestinal pathogenic strains.
The minimum inhibitory concentrations (MICs) for 12 antimicrobial agents (ampicillin [AMP], cefazolin [CFZ], cefotaxime [CTX], gentamicin [GEN], kanamycin [KAN], tetracycline [TET], minocycline [MIN], nalidixic acid [NAL], ciprofloxacin [CIP], colistin [CST], chloramphenicol [CHL], and sulfamethoxazole-trimethoprim [SXT]) were tested by broth microdilution method using frozen plates (Eiken Chemical Co., Ltd.) according to the manufacturer's instructions. The breakpoint MICs of these antimicrobial agents were determined according to the Clinical and Laboratory Standards Institute guideline M100 ED29 (Clinical and Laboratory Standards Institute, 2019). Breakpoint to CST was defined as ≥4 mg/L according to the European Committee on Antimicrobial Susceptibility Testing guideline (European Committee on Antimicrobial Susceptibility Testing, 2019).
To prevent analysis of duplicated strains, E. coli isolates from the same meat samples with the same phylogenetic type and antimicrobial susceptibility pattern were omitted from further analysis. Colony-direct PCR was performed using Quick Taq HS DyeMix (Toyobo Co., Ltd., Osaka, Japan) to detect the following antimicrobial resistance genes: bla TEM, bla SHV, and bla OXA for AMP-resistant strains; tetA, tetB, tetC, tetD, tetE, and tetG for TET-resistant strains; and sul1, sul2, and sul3 for SXT-resistant strains (Colom et al., 2003; Phuong Hoa et al., 2008; Vignaroli et al., 2012).
We tested the transferability of antimicrobial resistance genes from the retail meat isolates to other E. coli through conjugation assay by broth mating method (Potron et al., 2011). AMP-TET-SXT-resistant isolates were used as donors, and sodium azide-resistant E. coli J53 was used as the recipient. The donor and recipient were mixed at a 1:1 ratio (final concentration of both 107 colony-forming units [CFU]) in phosphate-buffered saline and incubated at 37°C for 18 h. We focused on the transfer of AMP resistance because (1) AMP is reported to be one of the most commonly used first-line antibiotics in East Africa; and (2) usage of AMP is recommended by the WHO Integrated Management of Childhood Illness for treatment of bacterial infections in infants (Ampaire et al., 2016). After mating, the solution was inoculated on LB agar (Nakarai Tesque, Inc., Kyoto, Japan) containing 100 mg/L of sodium azide with or without 50 mg/L of AMP. After overnight cultivation at 37°C, the number of colonies on selective plates was counted manually. Then, the CFU of transconjugants and recipients in the mating solution transfer frequencies were determined as the number of transconjugants per recipient. The transfer of antimicrobial resistance gene(s) was confirmed by PCR using representative recipient colonies.
Results
We isolated 103 E. coli isolates from 58 retail meat samples (58/64; 90.6%). Among them, 14 had the same phylogenetic type and antimicrobial susceptibility pattern and were thus suspected to be duplicated clones and omitted from further analysis. Therefore, we used 89 E. coli isolates in this study. The phylogenetic patterns of retail meat isolates were as follows: A0, 4.5% (4/89); A1, 7.9% (7/89); B1, 70.8% (63/89); B22, 2.2% (2/89); B23, undetected; D1, 13.5% (12/89); and D2, 1.1% (1/89).
Antimicrobial susceptibility testing showed that TET-resistant isolates were the most frequently detected (28.1%, 25/89), followed by AMP-resistant isolates (15.7%, 14/89) and SXT-resistant isolates (15.7%, 14/89; Fig. 2). Only a few isolates showed resistance to GEN (3.4%, 3/89), NAL (2.2%, 2/89), CIP (1.1%, 1/89), or CHL (1.1%, 1/89). Resistance to cephalosporins (CFZ and CTX), KAN, MIN, or CST was not detected. As for multidrug resistance, 10 isolates (11.2%) showed simultaneous AMP-TET-SXT resistance (Table 1).
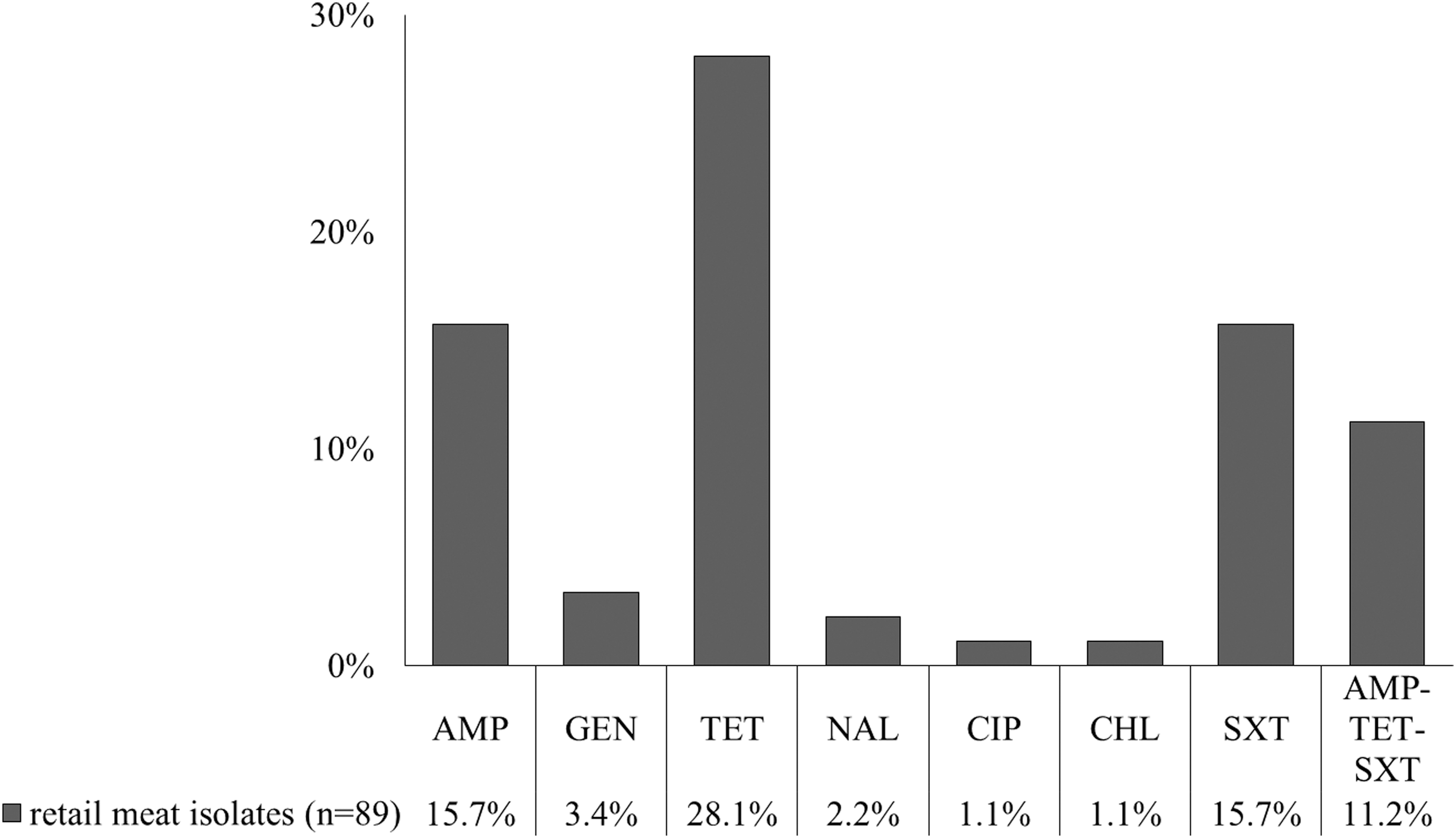
Frequency of resistant isolates (n = 89) to respective antimicrobial agents. AMP, ampicillin; CHL, chloramphenicol; CIP, ciprofloxacin; GEN, gentamicin; NAL, nalidixic acid; SXT, sulfamethoxazole–trimethoprim; TET, tetracycline.
Transfer Frequency of Multidrug-Resistant Escherichia coli from Retail Meats (n = 10) and Antimicrobial Susceptibility of Those Donors and Recipients
All donors in this table were isolated from beef ribs.
AMP, ampicillin; MIC, minimum inhibitory concentration; SXT, sulfamethoxazole–trimethoprim; TET, tetracycline.
All AMP-resistant retail meat isolates (14/14) harbored the bla TEM gene. TET-resistant retail meat isolates harbored several tet genes: tetA, 68.0% (17/25); tetB, 32.0% (8/25); tetE, 4.0% (1/25); and tetG, 4.0% (1/25). Two isolates harbored both tetA and tetB. The most prevalent SXT resistance gene was sul2 (14/14, 100%), and 7 isolates (50.0%) simultaneously harbored sul2 and sul1. sul3 was not detected.
We tested the transferability of AMP resistance using multidrug (AMP-TET-SXT)-resistant isolates (n = 10). Among them, six isolates successfully transferred their AMP resistance to E. coli J53. The transfer frequencies of those isolates were 10−3 to 10−4 (Table 1). PCR for antimicrobial-resistant genes showed that transconjugants derived from those six multidrug-resistant donors harbored the AMP-resistant gene, bla TEM, and the TET- and SXT-resistant genes, tetA, tetB, sul1, and/or sul2. On the contrary, AMP-resistant determinants in four isolates did not transfer to E. coli J53. The transfer frequencies of those isolates were lower than 10−7 to 10−8 (Table 1).
Discussion
Enteric bacterial pathogens such as E. coli on retail meats are mainly originated from fecal contamination of animal intestinal contents during meat processing (Brashears and Chaves, 2017). Among the retail meat samples in this study, 90.6% (58/64) were positive for E. coli. This frequency was relatively high compared with those of similar studies. E. coli prevalence in retail beef was reported to be 25.0% (14/56) in northwest Turkey, 49.2% (29/59) in Mexico, 68.9% (2061/2991) in the United States, and 75.0% (27/36) in Thailand (Eyi and Arslan, 2012; Zhao et al., 2012; Martínez-Vázquez et al., 2018). The information on bacterial contamination in retail meats in East Africa is limited, but Azege et al. reported that 70.0% (21/30) of retail meats in the butcher shops in Ethiopia are contaminated by Salmonella (Azage and Kibret, 2017). We hypothesized that the widespread detection of E. coli in Ugandan beef was because of the distribution network of the retail meats because the meats were slaughtered and directly transported from slaughterhouses to roadside butcheries on the same day. In addition, cross-contamination of E. coli between meats could have occurred because little attention appeared to be paid to hygiene control at the roadside butcheries (Fig. 1) or during transportation, resulting in a high prevalence of E. coli on the meat surfaces.
PCR-based phylogenetic typing showed that group B1 was the most frequently detected phylotype from retail meats (70.8%), followed by groups D1 (13.5%) and A0 (7.9%). E. coli groups A0, A1, and B1 are commensal strains that include non- or low-pathogenic E. coli in the animal digestive tract (Carlos et al., 2010). The high prevalence of E. coli group B1 from retail meats is consistent with a previous report that group B1 was mainly isolated from the feces of herbivorous animals, such as cows, and in retail meats (Johnson et al., 2009; Carlos et al., 2010). In addition, our previous study reported that phylotype B1 was also prevalent among fecal E. coli isolates from Ugandan livestock (Okubo et al., 2019). In agreement with those reports, our results suggest that E. coli in the livestock digestive tract can be distributed from farms to consumers directly through retail meats, especially under the conditions of a poor temperature-controlled supply chain.
Antimicrobial susceptibility testing showed that TET resistance was most frequently detected in E. coli isolates from retail meats in Uganda. The main TET-resistant determinant was the tetA gene, followed by the tetB gene. Our previous study revealed that TET resistance is common among E. coli from Ugandan livestock, and most of these bacteria harbored tetA and/or tetB gene(s) (Okubo et al., 2019). Another global study reported that human-derived TET-resistant atypical enteropathogenic E. coli was significantly more common among isolates from East Africa (60%) and West Africa (72%) compared with those in Asia (41%), suggesting that TET resistance is common among E. coli in African countries regardless of the host (Ingle et al., 2018). Resistance to AMP and SXT was also detected, and the major resistance determinants were bla TEM and sul2, respectively. Those genes were frequently detected from Enterobacteriaceae in Ugandan livestock samples in previous studies (Odoch et al., 2018; Okubo et al., 2019), suggesting that the prevalence of TET-, AMP-, or SXT-resistant E. coli and its resistance genes were common features among livestock-derived bacteria in Uganda. AMP, TET, SXT, and their derivatives are common antibiotics in Ugandan livestock farms (Okubo et al., 2019). This indicates that ARB on retail meats are coincident with what antibiotics the farmers used in livestock farms.
As for multidrug resistance, 10 isolates showed AMP-TET-SXT resistance. Among them, two isolates (strains RM20B and RM23A) were classified as phylogenetic types D2 and D1, respectively. Phylotypes D1 and D2 include intra-intestinal pathogenic strains (Escobar-Páramo et al., 2004), thus those multidrug-resistant D1 and D2 E. coli can be a risk factor for serious food poisoning. In addition, six isolates seemed to encode bla TEM gene on the same plasmid with tet genes and/or sul genes because those resistance genes were transferred to its recipients simultaneously. The spread of AMP-TET-SXT-resistant E. coli was detected at fecal isolates from Ugandan livestock (Okubo et al., 2019). These results suggest that multidrug-resistant E. coli can be spread from food-producing animals to consumers through retail meats.
The spread of bacteria with resistance to clinically important antimicrobial agents, such as ESBL-producing E. coli, is a global concern. Several review articles reported that ARB, especially ESBL-producing Enterobacteriaceae, in livestock is a risk factor to human health because these bacteria can be spread from farms to consumers through the food chain (Economou and Gousia, 2015; Founou et al., 2016). Among our isolates, no cephalosporin-resistant E. coli, which are suspected to be ESBL producers, were detected from retail meats. A previous cross-sectional study on E. coli derived from dairy cattle in Uganda reported that only 4 of 385 E. coli isolates (1.03%) were identified as ESBL producers harboring bla CTX-M-15 or bla CTX-M-27 genes (Ball et al., 2019). In contrast, a previous report from a national hospital in Uganda reported that 59.7% (28/42) of E. coli derived from blood specimens were resistant to CTX, and 70.2% (33/42) were resistant to ceftriaxone (Kajumbula et al., 2018). Another study in a rural area of Uganda reported that 11.8% of E. coli and Klebsiella pneumoniae from clinical stool samples (n = 21) showed resistance to the third- and fourth-generation cephalosporins (Stanley et al., 2018). The gap in frequency of cephalosporin-resistant E. coli between Ugandan animal-related samples and human samples suggests that ESBL-producing E. coli do not originate from livestock or retail meats but from human communities in Uganda.
In conclusion, we detected antimicrobial-resistant E. coli from retail meat from roadside butcheries in Uganda. Our results showed that >90% of the retail meats at roadside butcheries were contaminated with fecal-associated E. coli, and some were resistant to AMP, TET, and/or SXT. These findings suggest that ARB in livestock can easily be transferred to humans through the food chain. Little attentions appeared to be paid for hygiene conditions in those roadside butcheries. Thus, sanitary control is important for protecting consumers from foodborne diseases and preventing the spread of ARB. But it is not realistic to regulate or restrict all the roadside butcheries. Instead, we need to reduce ARB at the farm level. We considered that the prudent use of antimicrobials in food-producing animals is the most crucial thing to stop the further emergence and dissemination of ARB.
Footnotes
Acknowledgments
The Uganda National Council of Science and Technology (UNCST) approved the study protocol (Permit No. A_522). The authors thank I. Makhuwa and H.N. Opolot, Science Officer, Research Registration, Clearance and Analysis Unit, UNCST, for their help with this research project. The authors also thank Traci Raley, MS, ELS, from Edanz Group (
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was funded by the Japan Society for the Promotion of Science KAKENHI (Grant No. 16H02767). Eddie M. Wampande was supported through the DELTAS Africa Initiative grant #DEL-15-011 to THRiVE-2. The DELTAS Africa Initiative is an independent funding scheme of the African Academy of Sciences' (AAS) Alliance for Accelerating Excellence in Science in Africa (AESA) and supported by the New Partnership for Africa's Development Planning and Coordinating Agency (NEPAD Agency) with funding from the Welcome Trust grant #107742/Z/15/Z and the U.K. government.