
Abstract
Objectives:
The previous researches revealed that Vibrio parahaemolyticus has been detected in freshwater fish samples. However, the molecular characteristics of V. parahaemolyticus isolated from freshwater fish, including pathogenic and pandemic strains, are still unknown. This study aims to characterize and identify molecular properties of the bacterium. In addition, it identifies the source of V. parahaemolyticus from freshwater fish samples in Zhejiang Province, China.
Methods:
Four hundred and twenty-one freshwater fish samples (from fishing farms, retail markets, and restaurants) and 212 seafood samples (from retail markets) were collected in 10 cities of Zhejiang Province. V. parahaemolyticus strains were isolated from these samples and comparatively analyzed by multilocus sequence typing, serotyping, antimicrobial susceptibility test, and polymerase chain reaction, targeting common toxin genes (tdh, trh) and markers for pandemic strains (orf8, toxRS/new).
Results:
Sixty-eight V. parahaemolyticus strains were isolated from the 421 freshwater fish samples, and 89 V. parahaemolyticus isolates were identified out of 212 seafood samples. The detection rate of V. parahaemolyticus was significantly different (p < 0.05) between the fishing farms, the retail markets, and the restaurants. The isolates from freshwater fish samples were divided into eight O serotypes with three O3:K6 isolates, which contain three pandemic complexes (tdh +, orf8 +, toxRS/new +). A total of 53 different sequence types (STs) were identified among the 68 isolates, including 28 novel STs. Antimicrobial susceptibility results indicated that 76.5% of the strains were resistant to ampicillin. A third (3/9) of the isolates from fishing farm sources shared the same STs with their counterparts from retail markets. Compared with the isolates from the seafood samples collected in the same sampling sites, 13.2% (9/68) freshwater fish isolates overlapped with seafood isolates.
Conclusions:
Our study showed that V. parahaemolyticus population in freshwater fish is genetically diverse. The V. parahaemolyticus contaminates might have come from both fishing farm sources and cross-contamination from seafood in the closed area at the markets. Freshwater fish may work as a reservoir of pathogenic and pandemic V. parahaemolyticus isolates, indicating potential public health and food safety risks associated with the consumption of freshwater fish.
Introduction
V
China has a long history of freshwater fishing, and is the largest freshwater aquaculture country in the world. The Ministry of Agriculture, China announced 29,052,900 tons worth 84.5 billion U.S. dollars from freshwater aquaculture and 2,183,000 tons worth 6.6 billion U.S. dollars from freshwater fishing in 2017 (
As a halophilic bacterium, V. parahaemolyticus is found mostly in coastal areas. However, the prevalence of these organisms has gradually increased in inland cities in recent years, which is probably due to contamination of V. parahaemolyticus from freshwater products (Pei et al., 2016). V. parahaemolyticus isolates were obtained from freshwater crayfish, fish, and sediment (Chandran et al., 2011; Dong et al., 2016; Lee et al., 2018). Lee et al. (2018) showed a close relationship between isolates from freshwater fish and seafood samples. It is worth noting that the detection rate of V. parahaemolyticus in freshwater fish was more than 10% in China (Wang et al., 2017b). Molecular characteristics of V. parahaemolyticus isolated from freshwater fish, including pathogenic and pandemic strain, are still unknown, and therefore further investigations are required.
In 1996, V. parahaemolyticus serotype O3:K6 was first detected in India (Okuda et al., 1997). Since then, serotype O3:K6 was identified as a pandemic clone responsible for 50–80% of the infections all around the world (Chowdhury et al., 2013; Machado and Bordalo, 2016; Raszl et al., 2016; Guerrero et al., 2017). Thermostable direct hemolysin (TDH) and/or TDH-related hemolysin (TRH) are commonly associated with pathogenic strains of V. parahaemolyticus (Raghunath, 2015; Cai and Zhang, 2018). Hence, the genes encoding TDH and TRH are routinely used for detection of those strains. The tdh gene was observed in 80–90% of the clinical isolates and <1% of the environmental samples (Islam et al., 2004; Chen et al., 2016; Wang et al., 2017a), whereas the trh gene was detected in 1.0–4.0% of the clinical isolates (Chen et al., 2016; Wang et al., 2017a). The toxR gene is species specific, conserved among bacterial, and considered as signature molecular markers for V. parahaemolyticus.
Following the international spreading of the pandemic V. parahaemolyticus O3:K6 strains, surveys of the prevalence and characteristics of this bacterium in various food samples have been conducted in many countries. There are several serovariants of the pandemic strains of O3:K6, such as O4:K68, O1:K25, O1:26, and O1:KUT, which also have been recognized as pandemic clones. To identify pandemic clones, some virulence genes have been targeted as pandemic markers, such as orf8 and toxRS/new (Espejo et al., 2017). The orf8 gene encodes an adherence protein that increases the virulence of V. parahaemolyticus by adhering to host intestinal cells. The toxRS/new gene, which can be amplified by a group-specific PCR, encodes a transmembrane protein that plays an important role in the regulation of virulence genes. The orf8 and toxRS/new genes were observed in 60% of the clinical isolates and <1% of the environmental isolates (Yang et al., 2008; Chen et al., 2016; Wang et al., 2017a).
V. parahaemolyticus from clinical or environmental samples are always susceptible to the majority of antimicrobials tested; however, most of the isolates demonstrated a high resistance to ampicillin (AMP) (Han et al., 2007; Chen et al., 2016).
Multilocus sequence typing (MLST) is a technique for characterizing isolates of bacterial species using the DNA sequences of seven housekeeping genes. It is a useful method for characterizing, tracing, and analyzing genetic evolution of pathogens at the molecular level. Since MLST scheme for V. parahaemolyticus was first built in 2004, it has been increasingly used as a common tool for comparison of V. parahaemolyticus isolates from food as well as diarrhea patients (Rahman et al., 2017).
This research was performed in ten major cities of Zhejiang Province, which were located in eastern China. Zhejiang has over 640 km of coastline, which hosts the largest number of islands in China. Residents in this region consume uncooked or half-cooked fish regularly. Moreover, V. parahaemolyticus causes most of the food-borne illness among local citizens (Chen et al., 2019). To understand the incidences, sources and molecular characteristics of V. parahaemolyticus in freshwater fish, we obtained samples from freshwater fish, and V. parahaemolyticus isolates were analyzed for serotyping, expression of common virulence genes (tdh and trh) and pandemic markers (orf8 and toxRS/new), antibiotic susceptibility, and genetic diversity analysis by MLST.
Materials and Methods
Sampling
There are totally 11 prefecture-level cities in Zhejiang Province. One of the cities is an island city with no freshwater fish farms. Therefore, different sites (fishing farms, retail markets, restaurants) in the 10 cities of Zhejiang province were sampled from the second quarter to the fourth quarter of 2014 (Fig. 1). A total of 421 freshwater fish and 212 seafood samples were collected, and the samples were placed in sterile sealed plastic bags, transported to the laboratory in a cold box below 4°C, and analyzed immediately.
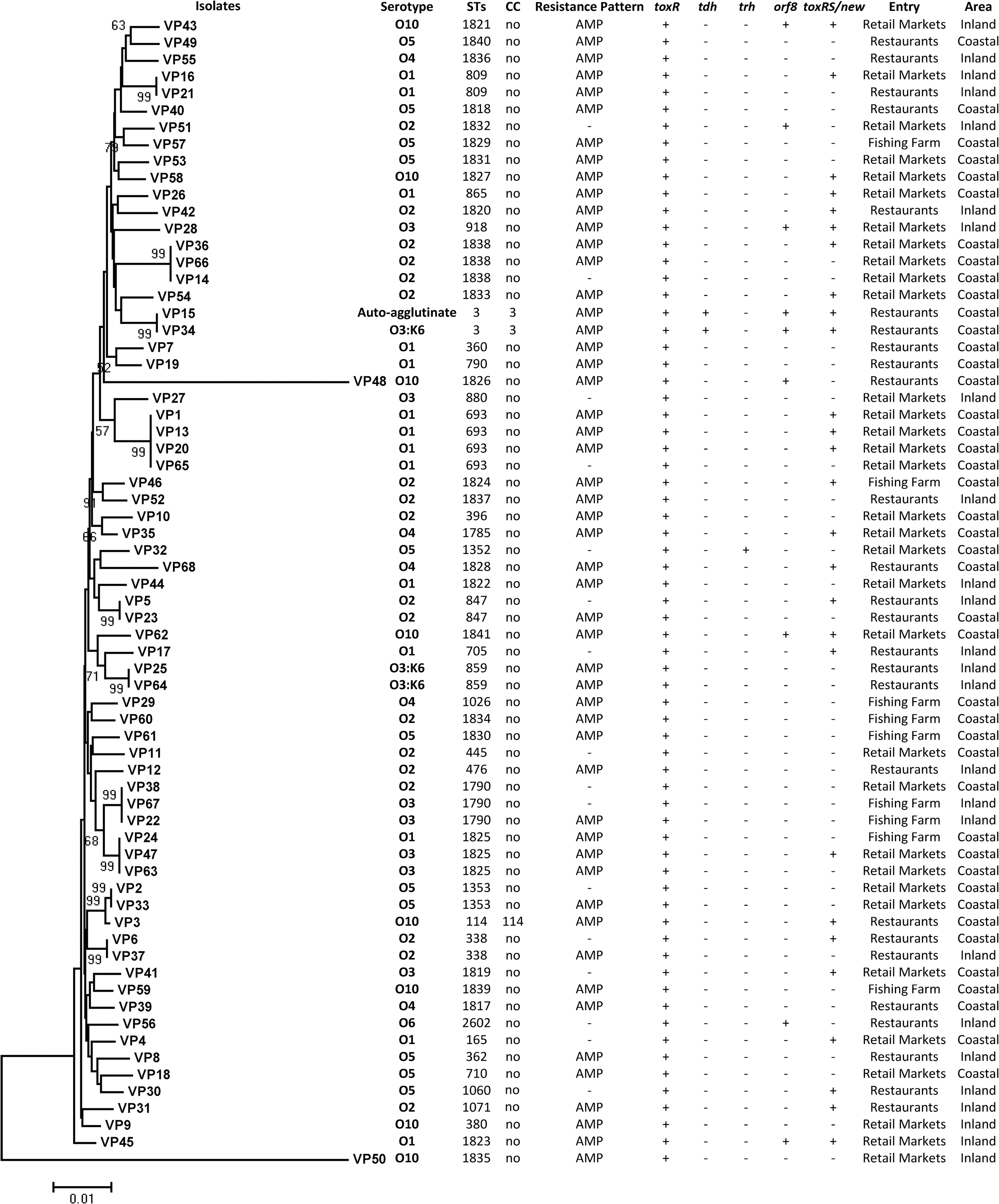
Characteristics of Vibrio parahaemolyticus strains isolated from freshwater fish in Zhejiang Province, China. The neighbor-joining phylogeny tree is constructed by using seven housekeeping genes according to the PubMLST database. The number of Bootstrap replications was 1000. AMP, ampicillin; CC, clonal complex; STs, sequence types.
V. parahaemolyticus strain isolation and serotyping
The V. parahaemolyticus strain was isolated according to the Bacteriological Analytical Manual of Vibrio (Kaysner and Depaola, 2004; GB4789.7, 2008). Briefly, each fish was rinsed with tap water and gently shook off excess water. A part of the meat and skin on the back of a fish was cut by sterile scissors. Twenty-five grams of the fish meat and skin samples was placed in 225 mL of sterile 3.5% NaCl, solution and homogenized in blenders. Homogenates were incubated at 37°C for 16–18 h. After incubation, a loopful of the top 1 cm was streaked onto thiosulfate–citrate–bile salts–sucrose (TCBS) agar plates and incubated at 37°C for 18–24 h. Presumptive colonies (green or blue green in color, 2–3 mm in diameter) were streaked onto Chromogenic Vibrio medium and incubated at 37°C for 24 h. The mauve colonies were selected for the further V. parahaemolyticus identification by oxidase activity test, Gram staining, 3.5% NaCl triple sugar iron tests, and API 20E diagnostic strips (BioMérieux Company). O-antigen was determined with 11 O (lipopolysaccharide) antisera (Denka Seiken Ltd., Tokyo, Japan), and isolates positive for O3 serotype were further tested by K6 (capsule) antisera (Denka Seiken Ltd.).
Determination of virulence genes and pandemic markers
The presence of tdh, trh, toxR, orf8, and toxRS/new genes was determined using the published primers and cycling parameters (Gil et al., 2007). Briefly, polymerase chain reaction (PCR) was performed in a 25 μL reaction mixture containing 12.5 μL Premix Taq DNA polymerase (TaKaRa, Tokyo, Japan), 4 μL of each primer mix (5 μM), 2 μL of DNA template, and 6.5 μL of ddH2O. After a 5 min predenaturation at 94°C, the PCR mixtures were subjected to 35 cycles of amplification at 94°C for 1 min, 55°C for 1 min, 72°C for 1 min, and a final extension of 72°C for 7 min. Then, electrophoresis of the PCR products was conducted with a 1.5% agarose gel. V. parahaemolyticus ATCC 33847 (tdh +) and ATCC 17802 (trh +) were used as positive controls, respectively. The pathogenic group was defined as tdh + and/or trh +, and all other isolates were assigned to the nonpathogenic group. The isolates in the pandemic group were defined as tdh +, trh −, toxRS/new +, and orf8 +/−, and all other isolates were assigned to the nonpandemic group.
Antibiotic susceptibility testing
The antibiotic resistances of all V. parahaemolyticus isolates were tested by broth microdilution minimum inhibitory concentration (MIC) method according to the guidelines of the Clinical and Laboratory Standards Institute (CLSI, 2017). The following antimicrobial compounds were assessed: cefoxitin (FOX, 0.5–32 mg/L), azithromycin (AZI, 0.12–16 mg/L), chloramphenicol (CHL, 2–32 mg/L), tetracycline (TET, 4–32 mg/L), amoxicillin/clavulanic acid 2:1 ratio (AUG, 1/0.5–32/16 mg/L), trimethoprim/sulfamethoxazole (SXT, 0.12/2.38–4/76), ciprofloxacin (CIP, 0.015–4 mg/L), ampicillin (1–32 mg/L), and gentamicin (GEN, 0.25–16 mg/L). Escherichia coli ATCC 25922 was included in the test for quality control. The MIC results were analyzed based on the breakpoints for Vibrio species advised in CLSI document M45 (CLSI, 2017).
Multilocus sequence typing
MLST was performed as previously described (Rahman et al., 2017). Briefly, seven housekeeping genes (dnaE, gyrB, recA, dtdS, pntA, pyrC, and tnaA) from all isolates were amplified by PCR following the protocol in the V. parahaemolyticus MLST database (
Statistical analysis
Data were entered to SPSS v. 18.0 software (SPSS, Inc.) for analysis. Test of significance was done using chi-square test. A p-value of <0.05 was considered statistically significant.
Results
Prevalence analysis
Sixty-eight V. parahaemolyticus isolates were identified out of the 421 freshwater fish samples from April to December 2014 in Zhejiang province. The detection rate of V. parahaemolyticus was significantly different (p < 0.05) among the fishing farms, the retail markets and the restaurants (Table 1). Eighty-nine V. parahaemolyticus isolates were identified out of 212 seafood samples.
The Detection Rate of Vibrio parahaemolyticus from Freshwater Fish in Zhejiang Province, China
Test of significance was done using chi-square test. A p-value of <0.05 was considered statistically significant.
Vp, V. parahaemolyticus.
Serotyping
Eight serotypes were identified among the 68 isolates, dominated by serotype O2 (18 isolates), O1 (14 isolates), and O5 (11 isolates). The results of the O-antigen serotyping for all 68 isolates are shown in Figure 1. Three O3:K6 isolates were found.
Detection of virulence genes
All the isolates was positive for toxR and tlh, which were in good agreements with the previous biochemical identification experiments (Cai and Zhang, 2018). Two isolates were positive for tdh gene; 1 was positive for trh gene; 4 were positive for orf8 gene; and 27 were positive for toxRS/new gene (Fig. 1).
Antibiotic profiles
The antibiotic profiles of the 68 isolates with respect to 10 antibiotics are shown in Supplementary Table S1. Most (52 isolates, 76.5%) of the isolates were resistant to ampicillin (Fig. 1). None of the V. parahaemolyticus isolates was resistant to cefoxitin, azithromycin, chloramphenicol, tetracycline, amoxicillin/clavulanic acid, trimethoprim/sulfamethoxazole, ciprofloxacin, and gentamicin. Around 88.9% (8/9) of isolates from fishing farm, 72.7% (24/33) from retail markets, and 76.9% (20/26) from restaurants show ampicillin resistance.
MLST analysis
MLST profiles of the V. parahaemolyticus isolates are presented in Supplementary Table S1. Among the 68 isolates, a total of 53 different STs were identified, including 28 novel STs (ST1817 to 1843, ST2602), indicating a high degree of genotypic diversity (Supplementary Table S1). Forty-three of the STs included single isolates, whereas the 10 STs contained 2–4 isolates. The most common STs were ST693 (5.9%, 4/68), ST1790 (4.4%, 3/68), ST1825 (4.4%, 3/68), and ST1838 (4.4%, 3/68). Among all the STs detected from the freshwater fish samples, only ST3, ST338, and ST476 have been reported in diarrhea patients before (Han et al., 2014, 2015). Two ST3 V. parahaemolyticus isolates belonged to the V. parahaemolyticus pandemic complex. Twenty-three alleles have not been previously described, including dnaE 365–367, gyrB 498, 499, recA 363–366, dtdS 432–435, pntA 247–249, pyrC 407–410, and tnaA 260–262.
Looking at the relationship between STs and sampling area, we found that 16.2% (43/266) of samples from coastal areas were positive for V. parahaemolyticus. Among all the isolates, 22 STs were identified, including 2 ST3 which were positive for tdh, orf8, and toxRS/new. The 16.1% (25/155) of samples from inland areas were positive for V. parahaemolyticus, including 26 STs, and none of the isolates contained tdh or trh gene. ST338, ST847, and ST1790 were found from both coastal and inland areas (Fig. 2A).
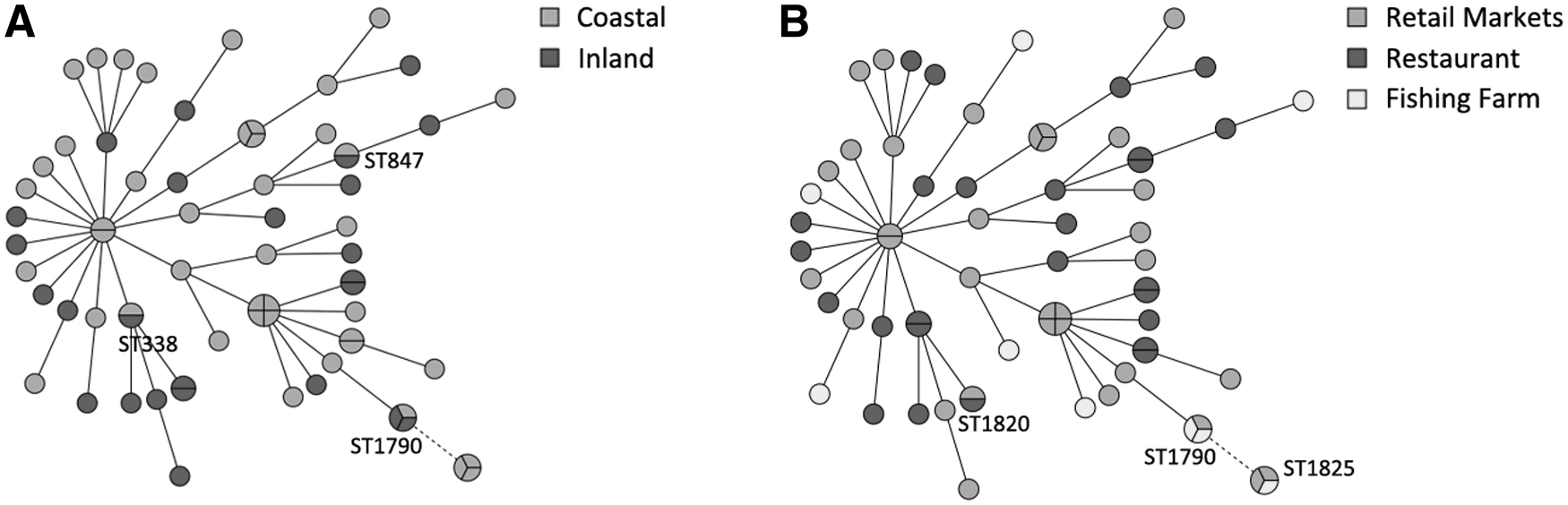
Minimum spanning tree analysis of freshwater fish Vibrio parahaemolyticus isolates from different kinds of area
Considering that the STs of V. parahaemolyticus isolates in freshwater fish might vary with different sampling sources, we made sense to analyze the relationship between STs as well as sampling entries and areas (Fig. 1). The detection rates of V. parahaemolyticus from fishing farms were low (7.5%, 9/120), and we could not find any tdh or trh genes in those isolates. A total of nine isolates from the fishing farm were genotyped into eight STs, displaying high genetic diversity. The 15.9% (33/208) of samples from the retail markets were positive for V. parahaemolyticus, including 26 STs, and none of the isolates contained the tdh or trh genes. The isolation rates of V. parahaemolyticus were 28% (26/93) in the restaurant samples. Among all the isolates, 22 STs were identified, including 2 ST3, which were positive for tdh, orf8, and toxRS/new. Overlapping of STs among retail markets, restaurants, and fishing farm sources was low, only ST1790, ST1820, and ST1825 as shown in Figure 2B.
Compared with V. parahaemolyticus from seafood collected in the same sampling sites, the diversity of strains from both sources was abundant (Fig. 3, Table 2, and Supplementary Table S2). Around 13.2% (9/68) freshwater fish isolates overlapped with seafood isolates. The overlapping STs included ST3, ST114, ST338, ST859, ST865, and ST880. The difference between the detection rate of the pathogenic group from freshwater fish and seafood isolates was not significant (p = 0.081) (Fig. 3B), as well as the pandemic group (p = 0.283) (Fig. 3C).
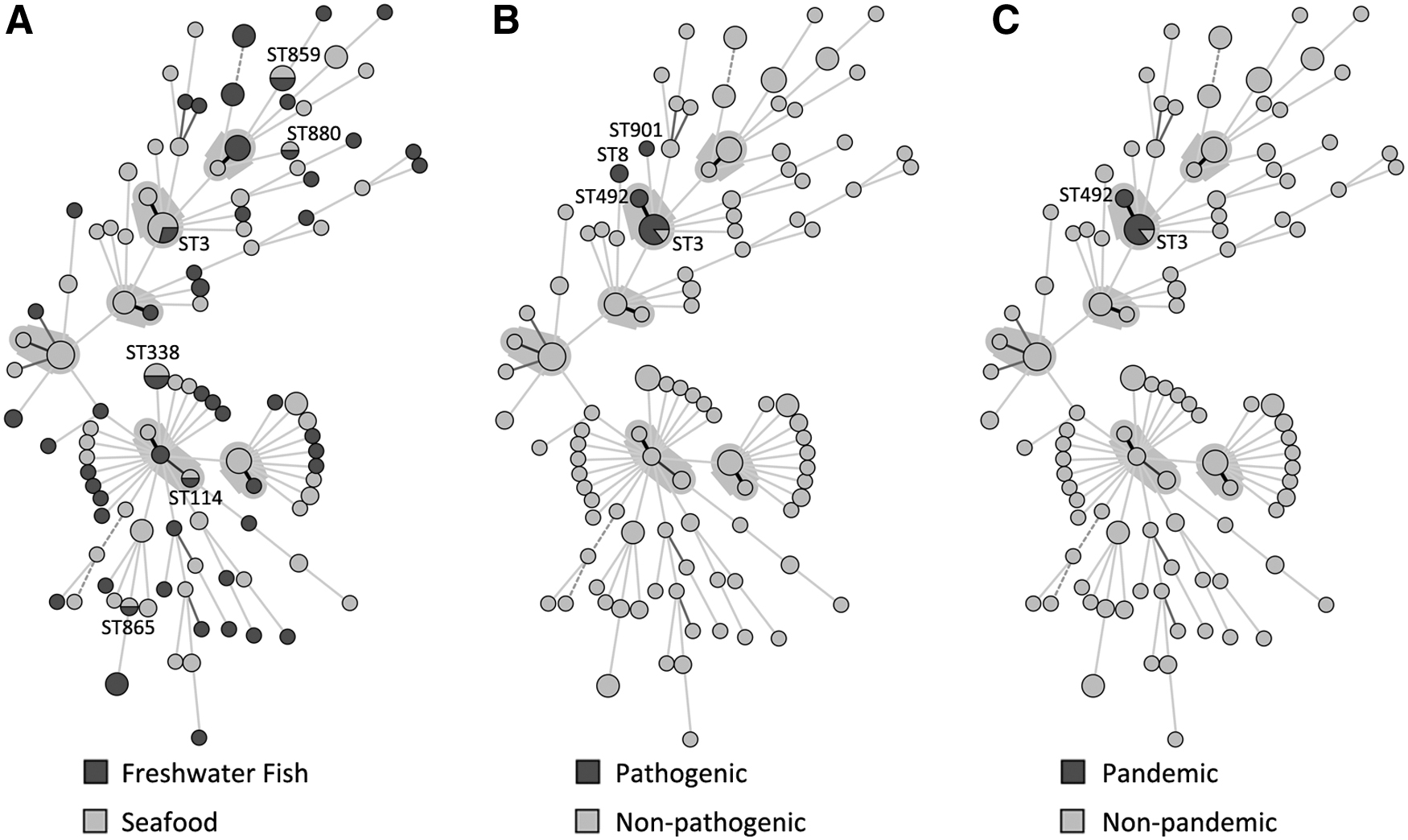
Phylogenetic analysis of Vibrio parahaemolyticus isolates in Zhejiang Province, China. The minimum spanning trees are based on MLST analysis of 68 freshwater fish and 89 seafood isolates. STs are represented by circles connected by branches proportional to the allelic distance, and the area of each circle corresponds to the number of isolates of a given ST in the PubMLST database. Colors represent freshwater fish/seafood strains
The Results of Vibrio parahaemolyticus Detection in Freshwater Fish and Seafood Samples for Vibrio parahaemolyticus, Pathogenic Vibrio parahaemolyticus, and Pandemic Vibrio parahaemolyticus in Zhejiang Province, China
Pathogenic Vp, tdh +, and/or trh + Vp isolates; Pandemic Vp, tdh +, trh −, toxRS/new +, and orf8 +/− Vp isolates.
The percentage of Vp positive.
Vp, V. parahaemolyticus.
Discussion
V. parahaemolyticus is one of the leading causes of foodborne outbreaks especially in coastal areas, and the fast and global spreading of V. parahaemolyticus has become a serious public health concern. However, sources and virulence repertoire of V. parahaemolyticus are still not completely understood. Dietary structures in China are considerably different compared with other parts of the world, and food sources are abundant in a way that results in great diversity. Several studies have been conducted on characteristics of V. parahaemolyticus in various aquatic products in China. These studies targeted on a lot of freshwater products, such as freshwater shrimp, crayfish, and Chinese mitten crab (He et al., 2016; Huang et al., 2016; Xie et al., 2016; Witherall et al., 2019). In this study, we characterized and identified molecular properties of V. parahaemolyticus from freshwater fish samples.
V. parahaemolyticus is a halophilic bacterium usually found in shallow coastal waters as well as the deepest parts of the ocean, and it is highly abundant in aquatic environments, including estuaries, marine coastal waters, and sediments. Reports on V. parahaemolyticus are rarely seen in freshwater or associated products. V. parahaemolyticus has been isolated from river sources and freshwater environs (Sarkar et al., 1985; Bockemühl et al., 1986; Pei et al., 2016). In this study, the salinity is <0.2% in the water from all the fishing farms. We found that ATCC 17802 can grow in 0.1–0.5% salinity and ATCC 33847 can grow in 0.5% salinity. It has been proved that viable but nonculturable V. parahaemolyticus was highly resistant to low salinity (0% NaCl) inactivation (Wong and Wang,
We have found that more V. parahaemolyticus isolates were detected from the freshwater fish samples in the retail markets and restaurants. Similarly, Lee et al. (2018) has also reported the detection of V. parahaemolyticus from wet markets or supermarkets in Malaysia. Freshwater fish and seafood are always sold in the same area and share fish-culturing water in retail markets in China, which may be the reason that there are more V. parahaemolyticus contaminates in freshwater fish from the retail markets. We assumed that V. parahaemolyticus contaminates in retail market freshwater fish might be coming from both fishing farm and retail market seafood. Our MLST typing results showed that 99% similarity of some V. parahaemolyticus isolates collected from fishing farm and retail market entry, that is, VP38-VP67-VP22, VP24-VP47-VP63, and 13.2% (9/68) freshwater fish isolates overlapped with seafood isolates in retail markets. Another thing worth mentioning is that a third of the isolates from fishing farm sources share the same STs with its counterpart from retail market. Thus, a part of the V. parahaemolyticus contaminates might have come from fishing farm sources and cross-contamination of V. parahaemolyticus from seafood in the closed area at the markets.
Not all strains of V. parahaemolyticus cause illness; on the contrary, pathogenic (tdh+ and/or trh+ ) strains represent a small percentage of the total V. parahaemolyticus present in the environment or seafood (Jiang et al., 2018; Tran et al., 2018; Vu et al., 2018). The tdh and trh genes, especially the tdh gene, are considered major virulence factors in V. parahaemolyticus. Lee et al. (2018) have found that 4 V. parahaemolyticus isolates from 240 freshwater fish samples were trh + and none was tdh + in Malaysia markets. Our results confirmed 2 tdh + isolates and 1 trh + isolate from 301 freshwater fish samples in restaurants and markets. The discovery of tdh+ pandemic complex may be a public health threat, and further confirms that freshwater fish is a reservoir of V. parahaemolyticus virulent strains.
MLST is conveniently used to study the genetic diversity of V. parahaemolyticus isolates and analyze the relationship between isolates from local areas and other parts of the globe. In the current studies, marine and associated products show great diversity in MLST diversity (Jiang et al., 2018; Lopatek et al., 2018). Additionally, the MLST phenotype diversity is rich in freshwater products, such as freshwater shrimp, crayfish, and Chinese mitten crab. This phenomenon is also found in isolates collected from freshwater fish in our study. Furthermore, V. parahaemolyticus isolates with high similarity but differ in the serotypes, pattern of genes, and antibiotic resistance (i.e., VP1, VP13, VP20, VP65; VP38, VP67, VP22; VP24, VP47, VP63). Earlier observations showed that extensive diversity in serotypes, pattern of genes, and antibiotic resistance occurred among isolates representing the same ST (Theethakaew et al., 2013). ST3 causes infections worldwide and persists as the dominant type in Asia and America (Baker-Austin et al., 2014). In addition to ST3, other clinical STs, including ST338 and ST476 (Han et al., 2014, 2015), also collected in freshwater fish, indicated that the danger of gastroenteritis is still present for the consumption of freshwater fish.
Antibiotics are the mainstay of treatment options to manage infections caused by V. parahaemolyticus, and antimicrobial resistance could be a serious problem for therapy directed against this pathogen. Thus, we assess the antimicrobial susceptibility of V. parahaemolyticus isolates from freshwater fish samples. Resistance to ampicillin is most commonly associated with V. parahaemolyticus isolates in previous studies (Elmahdi et al., 2016; Tan et al., 2017). Our study was in accordance with this trend.
Conclusions
Overall, our study explored the relationship between freshwater fish and V. parahaemolyticus. We showed that V. parahaemolyticus population in freshwater fish is genetically diverse. Among the 68 isolates, a total of 53 different STs were identified, including the presence of 28 novel STs. Furthermore, three pandemic complexes (tdh +, orf8 +, toxRS/new +) were identified, revealing that freshwater fish may work as a reservoir of pathogenic V. parahaemolyticus isolates.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Key Research and Development Program of China (2017YFC1601503), 13th Five-Year National Major Science and Technology Projects of China (2018ZX10714002), Medical and Health Science and Technology Project of Zhejiang (2018RC027, 2020KY524, WKJ-ZJ-1917), the Natural Science Foundation of Zhejiang Province (LY20H190001), and the National Natural Science Foundation of China (81801978).
Supplementary Material
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.