
Abstract
Although Enterococcus faecalis is known as a commensal microorganism in the gastrointestinal tract, it is linked to various foodborne infections. In addition, biofilm formation in E. faecalis is associated with the infections by exacerbating inflammation. Hence, we demonstrated that bacteriocins produced by Pediococcus acidilactici exhibited antibiofilm and anti-inflammatory activities against E. faecalis. Bacteriocins of P. acidilactici K10 and HW01 strains significantly reduced biofilm formation by E. faecalis on surfaces of polystyrene (p < 0.005 and p < 0.01 at 24 h, respectively) and stainless steel (p < 0.005 and p < 0.01 at 72 h, respectively), while both bacteriocins did not effectively reduce the growth of E. faecalis planktonic cells. Moreover, extracellular polymeric substances (EPSs) produced by E. faecalis were substantially decreased in the presence of P. acidilactici bacteriocin (p < 0.005), suggesting that E. faecalis biofilm formation was reduced by decreasing the production of EPSs, but not by killing bacteria. The bacteriocin of P. acidilactici also reduced the adhesion of E. faecalis to human intestinal epithelial cells (p < 0.005). Furthermore, both bacteriocins significantly inhibited E. faecalis-induced interleukin-8 production in human intestinal epithelial cells (p < 0.01 for K10 bacteriocin and p < 0.005 for HW01 bacteriocin). These results suggest that the bacteriocin of P. acidilactici can eradicate E. faecalis biofilms and inhibit the E. faecalis-induced inflammatory response in intestinal epithelial cells.
Introduction
Enterococcus faecalis is a major foodborne pathogen and frequently isolated from contaminated foods (Giraffa, 2002). Although E. faecalis is a part of the normal flora in the intestine, it can cause various types of infections, most notably bacteremia, endocarditis, and urinary tract infections, and has emerged as an important nosocomial pathogen (Fisher and Phillips, 2009).
Biofilm formation by enterococci is considered as an important factor contributing to their pathogenesis (Creti et al., 2004). Several foodborne pathogens such as Escherichia coli can form biofilms (Sharma and Anand, 2002). A biofilm is a complex microbial ecosystem, formed by single or multiple species, which is associated with an extracellular matrix of polysaccharides (Costerton et al., 1995). It is also considered to be a major cause of food contamination, compromising food safety and quality (Srey et al., 2013). Since cells in a biofilm are highly resistant to heat, desiccation, acidic conditions, and antibiotics (Davies, 2003), they are up to 1000 times more resistant than planktonic cells to antimicrobial agents, such as antibiotics (Penesyan et al., 2015), which makes biofilm-associated infections difficult to treat.
Several chemical sanitizers, including sodium hypochlorite, are prevalently used for the control of biofilms (Cabeca et al., 2012). Nonchemical agents, including enzymes and phages, have been also suggested to eradicate biofilms (Srey et al., 2013). Moreover, bacteriocins of lactic acid bacteria seem to control biofilms (Okuda et al., 2013; Smith et al., 2016).
The bacteriocin of E. faecalis inhibits Candida albicans (Graham et al., 2017). However, the inhibitory effect of bacteriocin on E. faecalis biofilms has not yet been thoroughly investigated. Thus, in this study, we demonstrated that bacteriocins of Pediococcus acidilactici reduce E. faecalis biofilm formation. Since the biofilm is closely associated with adhesion and inflammation (Di Domenico et al., 2018), the effect of the P. acidilactici bacteriocin on the adhesion and inflammatory response of E. faecalis in human intestinal epithelial cells was also examined. P. acidilactici bacteriocin effectively reduced E. faecalis biofilm formation as well as adhesion to intestinal epithelial cells. Additionally, P. acidilactici bacteriocin also inhibited the inflammatory response in intestinal epithelial cells.
Materials and Methods
Bacterial strains and culture conditions
P. acidilactici K10 and HW01 strains were previously characterized as bacteriocin-producing strains (Kwon et al., 2002; Ahn et al., 2017) and cultured in de Man, Rogosa, and Sharpe (MRS) medium (Neogen, Lansing, MI) at 37°C and 30°C, respectively. E. faecalis KCTC 3511, which was isolated from a urine specimen, was purchased from Korean Collection for Type Cultures (Jeongeup, Korea) and cultured in brain-heart infusion (BHI) medium (BD Biosciences, Franklin Lakes, NJ) at 37°C.
Preparation of P. acidilactici bacteriocin
Culture supernatants were obtained and filtered through a 0.2-μm filter (Advantec Toyo Kaisha, Tokyo, Japan). Ammonium sulfate (Samchun Chemical, Seoul, Korea) was then added to reach 70% saturation and the supernatants were thoroughly stirred at 4°C overnight. Following centrifugation, precipitates containing the bacteriocin were dialyzed against distilled water using a dialysis tubing (molecular weight cutoff 1200; Sigma-Aldrich, St. Louis, MO) at 4°C for 24 h. The concentration of the bacteriocin was determined using a bicinchoninic acid protein assay (Thermo Fisher Scientific, Rockford, IL).
Biofilm formation assay
Previous studies have shown that inhibitory concentrations vary depending on the strains of lactic acid bacteria and purification procedure of bacteriocins. In particular, it is likely that the relatively high concentrations of crude bacteriocins exhibited antibacterial activity (Biswas et al., 2017; Wang et al., 2018). According to previous studies, different concentrations (0.5, 1, 2, and 4 mg/mL) of P. acidilactici K10 and HW01 bacteriocins (K10 bacteriocin and HW01 bacteriocin, respectively) were added to a microtiter plate. A same volume of E. faecalis culture suspension (1 × 107 colony-forming units [CFUs]/mL) was added and incubated at 37°C for 24 h to allow biofilm formation. After incubation, the microtiter plate was washed with phosphate-buffered saline (PBS) and the biofilm cells were stained with 0.1% crystal violet for 30 min. The microtiter plate was washed with PBS to remove excess stain, and crystal violet was dissolved in 0.1% acetic acid and 95% ethanol. The optical density (OD) was measured at 595 nm using a microplate reader (Allsheng, Hangzhou, China). An E. faecalis biofilm without bacteriocins was used as the positive control. To determine whether the bacteriocin disrupts the preformed E. faecalis biofilm, E. faecalis culture suspension was transferred to a microtiter plate and incubated at 37°C for 24 h. Afterward, the same volume of the P. acidilactici bacteriocin (1 mg/mL) was added and incubated for an additional 24 h. Disruption of the E. faecalis biofilm was measured as described above. The E. faecalis biofilm without bacteriocins served as the positive control.
Confocal laser scanning microscopy
E. faecalis was grown on confocal dishes (SPL Life Sciences, Pocheon, Korea) to allow biofilm formation with or without the P. acidilactici bacteriocin (1 mg/mL) at 37°C for 24 h, and the E. faecalis biofilm was stained using a bacterial viability kit (Thermo Fisher Scientific). Biofilm formation was visualized using a confocal laser scanning microscope (Eclipse Ti-E, Nikon, Tokyo, Japan).
Viability and growth kinetics of E. faecalis planktonic cells
The XTT (2,3-bis-[2-methoxy-4-nitro-5-sulfophenyl]-2H-tetrazolium-5-carboxanilide) reduction assay kit was used according to the manufacturer's instructions (Biotium, Fremont, CA). E. faecalis culture (200 μL) with or without the P. acidilactici bacteriocin (1 mg/mL) was mixed with XTT solution (100 μL) and incubated at 37°C for 4 h. Absorbance was measured at OD492. The background absorbance values at OD630 were subtracted from absorbance values at OD492 to normalize absorbance. E. faecalis was cultured with or without the P. acidilactici bacteriocin (1 mg/mL) at 37°C for 1, 3, 6, 12, and 24 h. At each time point, absorbance was measured at OD595. Additionally, the bacterial culture from the 24-h incubation was serially diluted and plated on BHI agar for enumeration. All E. faecalis cultures without bacteriocins were used as positive controls.
Production of extracellular polymeric substances by E. faecalis
Production of extracellular polymeric substances (EPSs) was performed as described by Ambalam et al. (2012) with minor modifications. Briefly, after the culture of E. faecalis with or without bacteriocins (1 mg/mL) for 24 h, bacterial pellets were diluted in 1% tryptone and 0.5% NaCl to obtain the turbidity of 1.0 at OD600. Thereafter, 500 μL of bacterial suspension and 500 μL of Congo red (80 μg/mL) were mixed and incubated at 37°C for 2 h with vigorous agitation in the dark. The amount of Congo red remaining in the supernatants was measured at OD492 and compared with the Congo red standard curve that was obtained from Congo red absorbance at OD492 versus concentration. Production of EPSs by E. faecalis without bacteriocins was used as the positive control.
Antibiofilm activity of the P. acidilactici bacteriocin on stainless steel
Biofilm assay on stainless steel was performed as previously described (Kim et al., 2019). Briefly, stainless steel coupons (SSCs) (Type 304, 1 × 1 cm, no. 4 finish) were sonicated at 60°C for 20 min in 15% (v/v) phosphoric acid and 15% (v/v) alkaline detergent solutions, washed by soaking in distilled water, and then boiled in distilled water for 10 min. After SSCs were autoclaved and dried at 60°C, they were placed in a 24-well culture plate and E. faecalis (1 × 107 CFUs/mL) was added to the culture plate with or without the P. acidilactici bacteriocin (1 mg/mL). Following incubation at 37°C for 72 h, SSCs were gently rinsed with PBS and extensively scraped to detach biofilm cells, which were then enumerated on BHI agar. The E. faecalis biofilm without bacteriocins was used as the positive control.
Inhibition of adhesion of E. faecalis to Caco-2 cells
A human intestinal cell line, Caco-2 cells (American Type Culture Collection, Manassas, VA), was cultured in complete Dulbecco's modified Eagle's medium (DMEM; Welgene, Gyeongsan, Korea) supplemented with 10% heat-inactivated fetal bovine serum (Gibco, Burlington, ON, Canada), 100 unit/mL penicillin, and 100 μg/mL streptomycin (HyClone, Logan, UT) in a humidified incubator at 37°C in 5% CO2. Postconfluent Caco-2 cells were rinsed with PBS and E. faecalis (1 × 108 CFUs/mL) suspended in antibiotic-free DMEM was treated with or without the P. acidilactici bacteriocin (1 mg/mL) at 37°C for 1 h under gentle agitation. Caco-2 cells were lysed by adding 0.2% Triton X-100 (Sigma-Aldrich) for 10 min. After vigorous pipetting, Caco-2 cell suspensions were serially diluted and E. faecalis adhesion was quantified by plating on BHI agar. E. faecalis attachment without bacteriocins was used as the positive control.
Inhibition of E. faecalis-induced interleukin-8 production in Caco-2 cells
Postconfluent Caco-2 cells were further incubated for 18 h in serum-free DMEM, and subsequently, cells were treated with E. faecalis (1 × 108 CFUs/mL) with or without the K10 or HW01 bacteriocin (1 mg/mL) at 37°C for 24 h. Cell culture supernatants were collected, and interleukin (IL)-8 production was determined using a commercial IL-8 enzyme-linked immunosorbent assay kit (R&D Systems, Minneapolis, MN). Production of IL-8 in Caco-2 cells without or with E. faecalis treatment was used as the negative or positive control, respectively.
Statistical analysis
All data presented in this study are expressed as mean ± standard deviation of triplicate samples obtained from at least three independent replicates. The statistical significance was determined in comparison with the appropriate controls by an unpaired two-tailed t-test using GraphPad Prism 5 (GraphPad Software, Inc., La Jolla, CA). The level of statistical significance was defined as a p-value of *<0.05, ** <0.01, and *** <0.005.
Results and Discussion
P. acidilactici bacteriocin reduces biofilm formation by E. faecalis
In this study, our findings showed the inhibitory effect of P. acidilactici bacteriocin on the E. faecalis biofilm. To examine whether the P. acidilactici bacteriocin reduces E. faecalis biofilm formation, quantification of the biofilm was performed. As shown in Table 1, K10 and HW01 bacteriocins effectively reduced biofilm formation by E. faecalis. The effects of both bacteriocins on kinetics of biofilm formation by E. faecalis were examined. Although both bacteriocins failed to reduce biofilm formation by E. faecalis at 1 h, they exerted a significant inhibitory action on biofilm formation at 3 h (p < 0.005) that was further observed at 6, 12, and 24 h (Fig. 1A, B). Figure 1C shows that K10 and HW01 bacteriocins did not eradicate the preformed biofilm of E. faecalis, suggesting that the P. acidilactici bacteriocin interferes with biofilm formation by E. faecalis, but does not disrupt the E. faecalis biofilm. The microscopic analysis also confirmed that the P. acidilactici bacteriocin dramatically reduced biofilm formation by E. faecalis (Fig. 2). E. faecalis without the bacteriocin treatment displayed an evenly distributed and densely populated biofilm layer, while sparsely distributed biofilms were observed in the presence of the P. acidilactici bacteriocin, as evidenced by confocal laser scanning microscopy. These results indicate that P. acidilactici bacteriocin substantially decreased biofilm formation by E. faecalis. Some E. faecalis strains isolated from foods are known to be readily able to form biofilms (Marinho et al., 2013; Anderson et al., 2015). It has been shown that chemical and nonchemical compounds inhibit E. faecalis biofilms. Sodium hypochlorite reduced biofilm formation by E. faecalis (Rosen et al., 2016). Zilm et al. (2017) also showed that D-amino acids such as
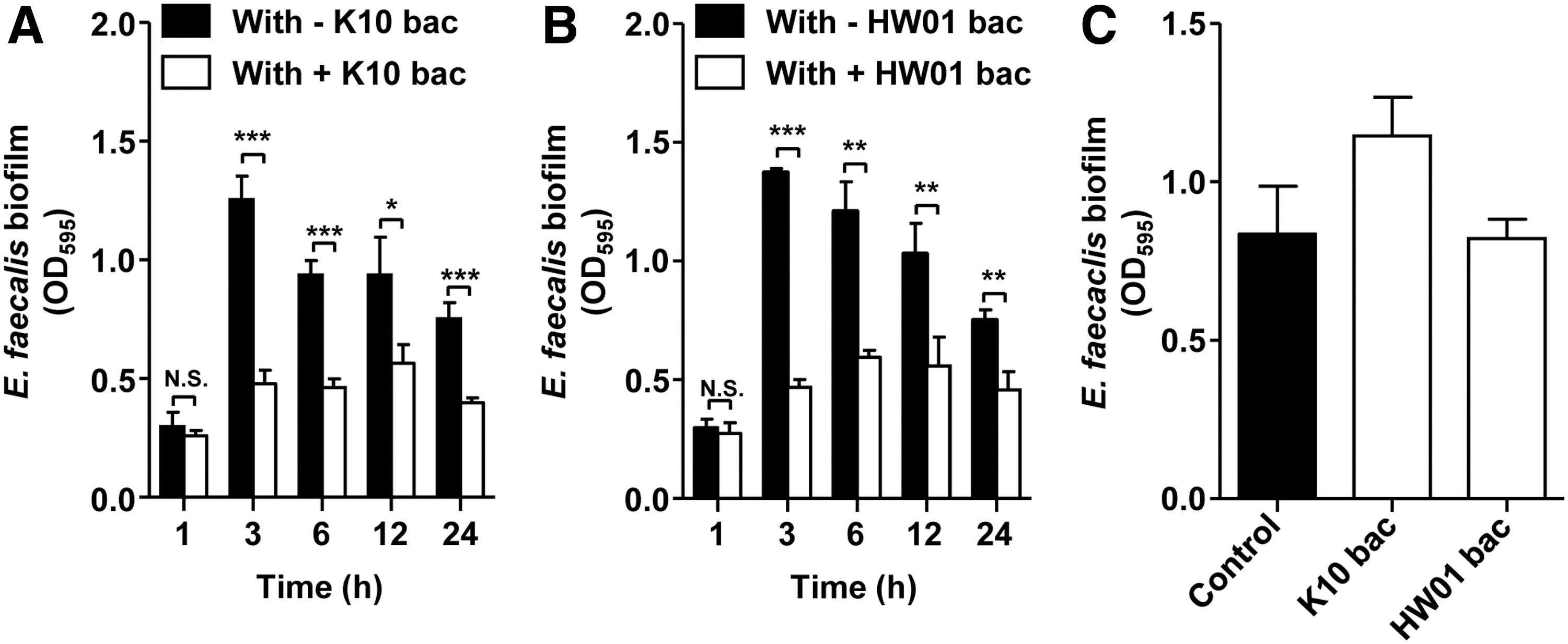
Inhibitory effects of K10 and HW01 bacteriocins on biofilm formation. Enterococcus faecalis was cocultured with either K10

Microscopic analysis of inhibition of biofilm formation by Enterococcus faecalis in the presence of K10 and HW01 bacteriocins. E. faecalis was cultured with either K10 or HW01 bacteriocin for 24 h, and biofilm formation was analyzed by confocal laser scanning microscopy. Images taken from one of three similar results are shown.
Antibiofilm Effect of K10 and HW01 Bacteriocins Against Enterococcus faecalis
Results are shown as mean with standard deviation by measuring turbidity at 595 nm.
p-Values are determined by comparison with the control (without treatment).
OD, optical density.
P. acidilactici bacteriocin does not affect the viability of E. faecalis
To examine the mode of action of the P. acidilactici bacteriocin against biofilm formation by E. faecalis, effects of the P. acidilactici bacteriocin on the viability of E. faecalis planktonic cells, XTT reduction, growth kinetics of E. faecalis planktonic cells, and production of EPSs were examined. As shown in Figure 3A, CFUs of E. faecalis planktonic cells after a 24-h incubation were comparable with or without the P. acidilactici bacteriocin, implying that the reduced biofilm formation was not attributed to E. faecalis killing by the P. acidilactici bacteriocin. To further confirm the viability of E. faecalis planktonic cells with or without bacteriocin at 24 h, the XTT assay was performed as a measure of metabolic activity to estimate viable cells. Figure 3B confirms that no XTT reduction occurred with or without K10 and HW01 bacteriocins, suggesting that the P. acidilactici bacteriocin does not reduce E. faecalis biofilms by directly killing bacteria. Although the growth rate of E. faecalis in the presence of the P. acidilactici bacteriocin was reduced at 3 to 12 h of incubation, it was not significantly reduced by the P. acidilactici bacteriocin at 24 h of incubation (Fig. 3C). These results indicate that the P. acidilactici bacteriocin does not decrease biofilm formation by E. faecalis by killing bacteria. However, EPSs were reduced in the presence of both K10 and HW01 bacteriocins (Fig. 3D). The EPS is attached to the outer surface of bacteria and is considered as one of the major components during biofilm formation (Bala Subramanian et al., 2010). It promotes initial attachment of the cell to a surface as well as formation and maturation of the biofilm structure (Flemming and Wingender, 2010). Furthermore, sodium hypochlorite considerably inhibits EPS production by Alteromonas sp. and Pseudomonas sp. as well as the adhesion to glass (Rathi and Satheesh, 2012). Biofilm formation and EPS production of Pseudomonas aeruginosa PA14 were also decreased by ginger extract (Kim and Park, 2013). Therefore, production of EPSs is essential for biofilm formation. According to the findings in this study, bacteriocins can decrease the EPS production of E. faecalis and thereby reduce E. faecalis biofilm formation. Since EPSs play an important role in cell aggregation, resulting in biofilm formation (Bar-Zeev et al., 2012), a decrease in EPS production by P. acidilactici may interfere with cell aggregation, leading to reduced E. faecalis biofilms. It has been reported that antimicrobial peptides, such as bacteriocins, can degrade EPSs produced by Streptococcus mutans, leading to the inhibition of biofilm formation (Ansari et al., 2017). Although more research is needed to understand the precise mechanism of action, the P. acidilactici bacteriocin either degrades the production of EPSs or directly interferes with production of EPSs.
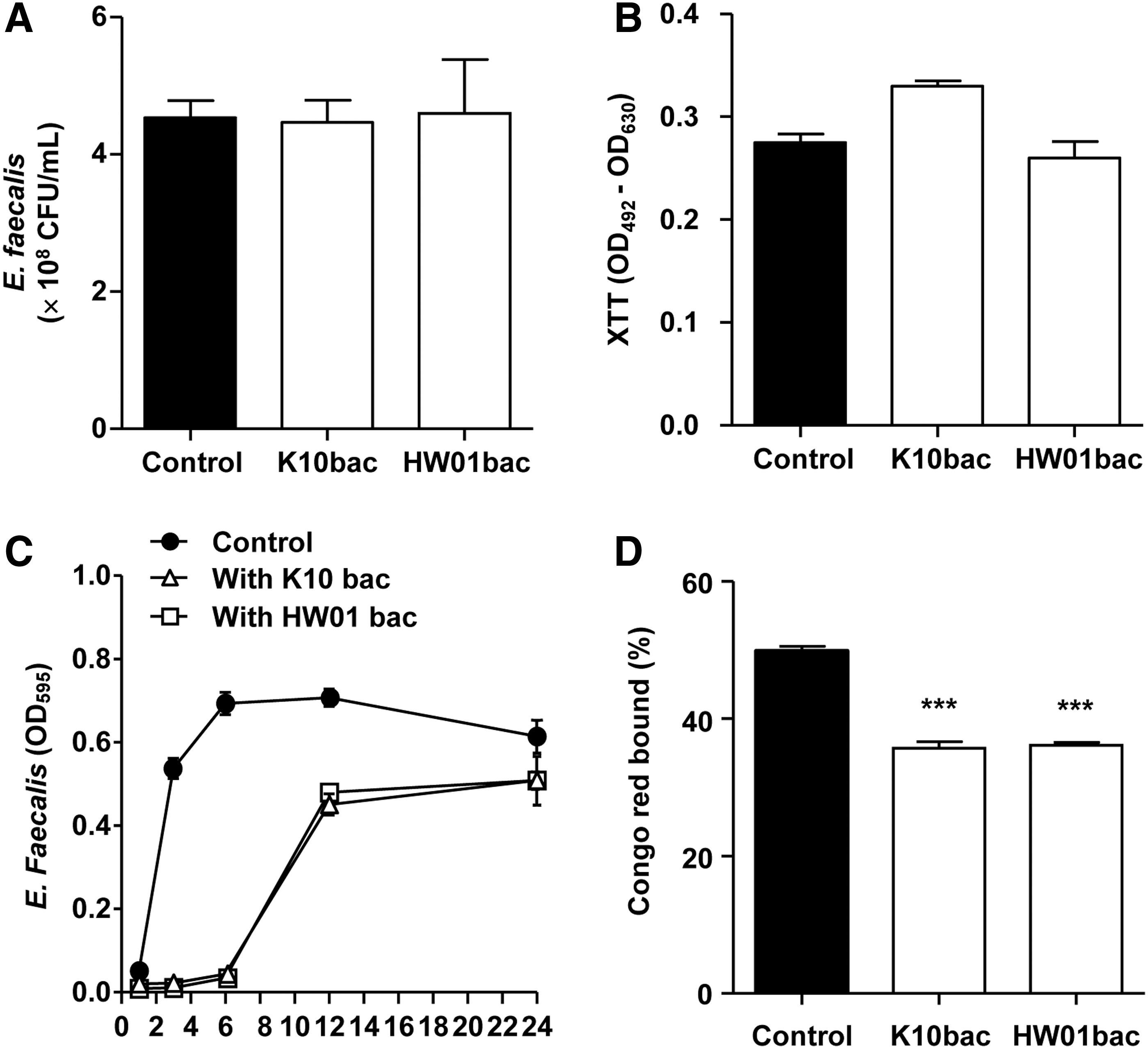
The viability of Enterococcus faecalis planktonic cells in the presence of K10 and HW01 bacteriocins.
P. acidilactici bacteriocin strongly reduces biofilm formation by E. faecalis on stainless steel
To our knowledge, reduction of E. faecalis biofilms on stainless steel by bacteriocins was first reported in the current study. Biofilm formation by E. faecalis on the SSC was significantly reduced by ∼90% in the presence of the K10 bacteriocin (p < 0.005) (Fig. 4A). The HW01 bacteriocin also reduced biofilm formation, although less effectively (∼40% reduction) than the K10 bacteriocin (Fig. 4B). Crude bacteriocin-like molecules extracted from various Lactobacillus fermentum strains showed different levels of antimicrobial activities against several pathogenic bacteria (Heredia-Castro et al., 2015), assuming that the antimicrobial activities may be strain specific. Our results also demonstrated that although K10 and HW01 bacteriocins were produced by P. acidilactici, both bacteriocins exhibited different levels of antibiofilm activities on the surfaces of SSCs. In the previous work using SSCs, several types of sanitization measures reduced E. faecalis biofilms (da Silva Fernandes et al., 2015), and chemical sanitation effectively removed biofilm formation of the mixed species of Enterococcus faecium, E. faecalis, and Bacillus cereus after being simultaneously grown (da Silva Fernandes et al., 2017). Negreiros et al. (2016) described that the essential oil of Baccharis psiadioides dramatically decreased the complex and dense E. faecalis biofilm on the SSC.
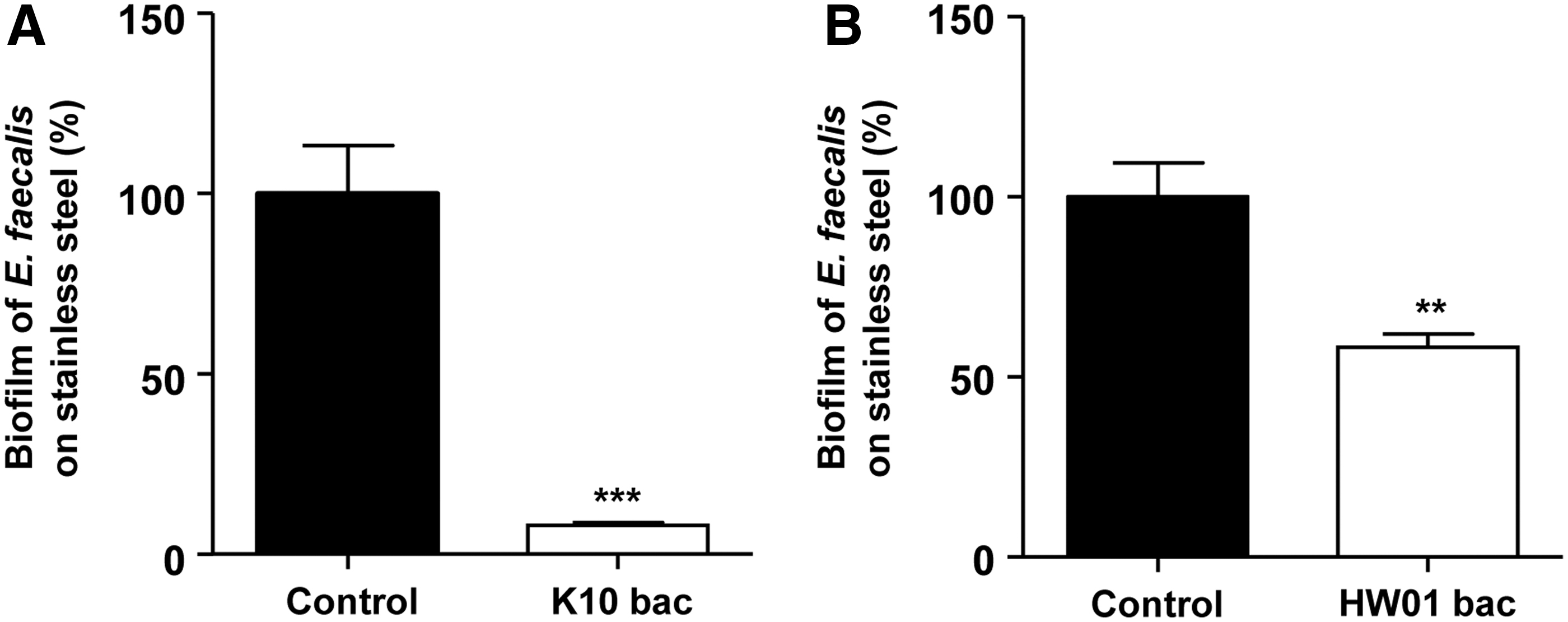
Antibiofilm activities of K10 and HW01 bacteriocins against Enterococcus faecalis on stainless steel. E. faecalis was incubated with or without K10
P. acidilactici bacteriocin reduces attachment of E. faecalis to intestinal epithelial cells
Our observations showed that the P. acidilactici bacteriocin efficiently inhibits the adhesion of E. faecalis to intestinal epithelial cells in vitro. As shown in Figure 5, the attachment of E. faecalis to Caco-2 cells was significantly inhibited by ∼90% and 60% in the presence of HW01 and K10 bacteriocins, respectively. The attachment to intestinal epithelial cells of pathogenic bacteria is associated with development of inflammation (Okumura and Takeda, 2017). In addition, bacterial attachment is an essential step for development of biofilm formation (Feng et al., 2015). E. faecalis is one of several microorganisms that easily attach to and form biofilms in the intestine (Barnes et al., 2017). Although inhibition of E. faecalis adhesion to intestinal epithelial cells has not been extensively studied, it has been demonstrated that lactic acid bacteria effectively inhibited the attachment of E. faecalis to Caco-2 cells (Todoriki et al., 2001; Singh et al., 2017; Fhoula et al., 2018). Additionally, bacteriocin-producing Bifidobacterium strains decreased the activation of adhesion and invasion of Listeria monocytogenes in Caco-2 and HT-29 cells (Moroni et al., 2006). However, it is not yet known if bacteriocins alone inhibit the adhesion of foodborne pathogenic bacteria to intestinal epithelial cells.
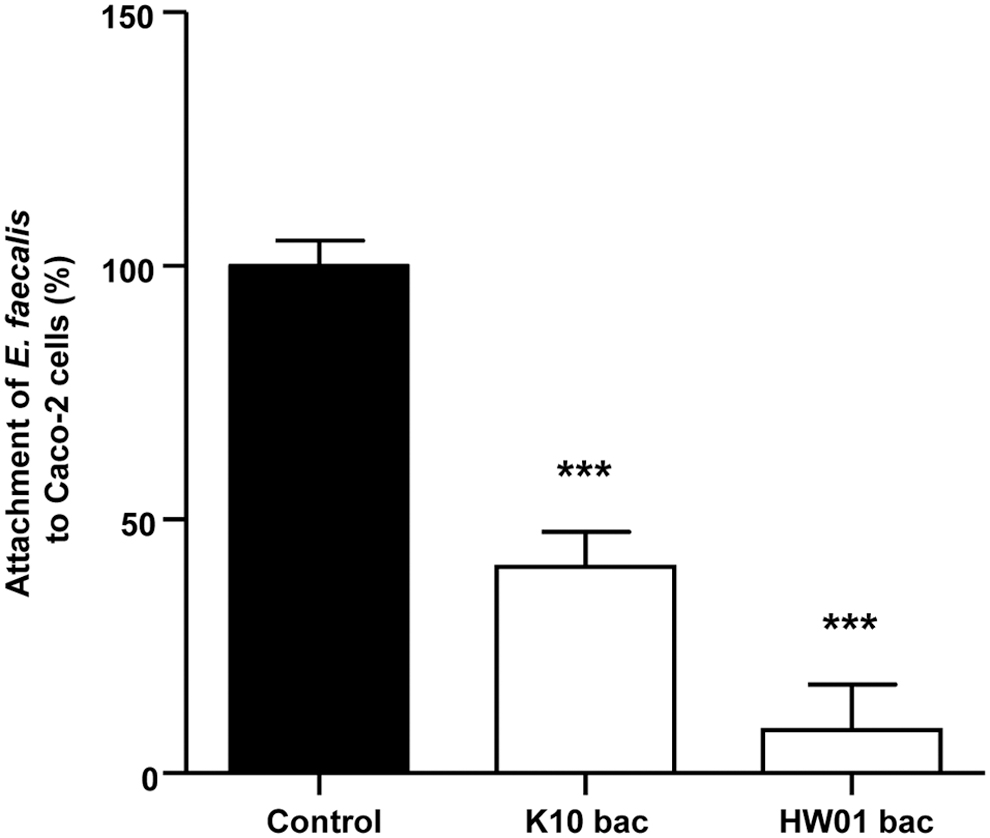
Inhibitory effects of K10 and HW01 bacteriocins on adhesion of Enterococcus faecalis to Caco-2 cells. Caco-2 cells were coincubated with E. faecalis in the presence or absence of either K10 or HW01 bacteriocin for 1 h. After incubation, the cells were washed, and attached E. faecalis cells were enumerated by plating on BHI agar. The adhesion of E. faecalis without bacteriocins was assigned 100% (control). Results are expressed as mean ± standard deviation from triplicate samples. Statistical significance compared with control (absence of bacteriocin) is indicated by ***p < 0.005. BHI, brain-heart infusion.
P. acidilactici bacteriocin inhibits IL-8 production in intestinal epithelial cells
To investigate whether the P. acidilactici bacteriocin inhibits IL-8 production in Caco-2 cells, live E. faecalis cells (1 × 108 CFUs/mL) were cultured with Caco-2 cells with or without the P. acidilactici bacteriocin for 24 h. As expected, E. faecalis significantly induced IL-8 production in Caco-2 cells (Fig. 6). However, K10 and HW01 bacteriocins dramatically inhibited E. faecalis-induced IL-8 production in Caco-2 cells (Fig. 6A and B, respectively). E. faecalis is involved in inflammatory responses in the intestine (Zhou et al., 2016). Strickertsson et al. (2013) reported that E. faecalis upregulated proinflammatory cytokines such as IL-8 and tumor necrosis factor-α in human gastric adenocarcinoma cells. Like the previous report, our findings showed that E. faecalis induced IL-8 production in Caco-2 cells. Although many lactobacilli have already been shown to reduce IL-8 production in intestinal epithelial cells (Ren et al., 2013; Lepine et al., 2018; Tuo et al., 2018), the current study also demonstrates the inhibitory activity of bacteriocins produced by Pediococcus on inflammatory responses in intestinal epithelial cells in vitro.
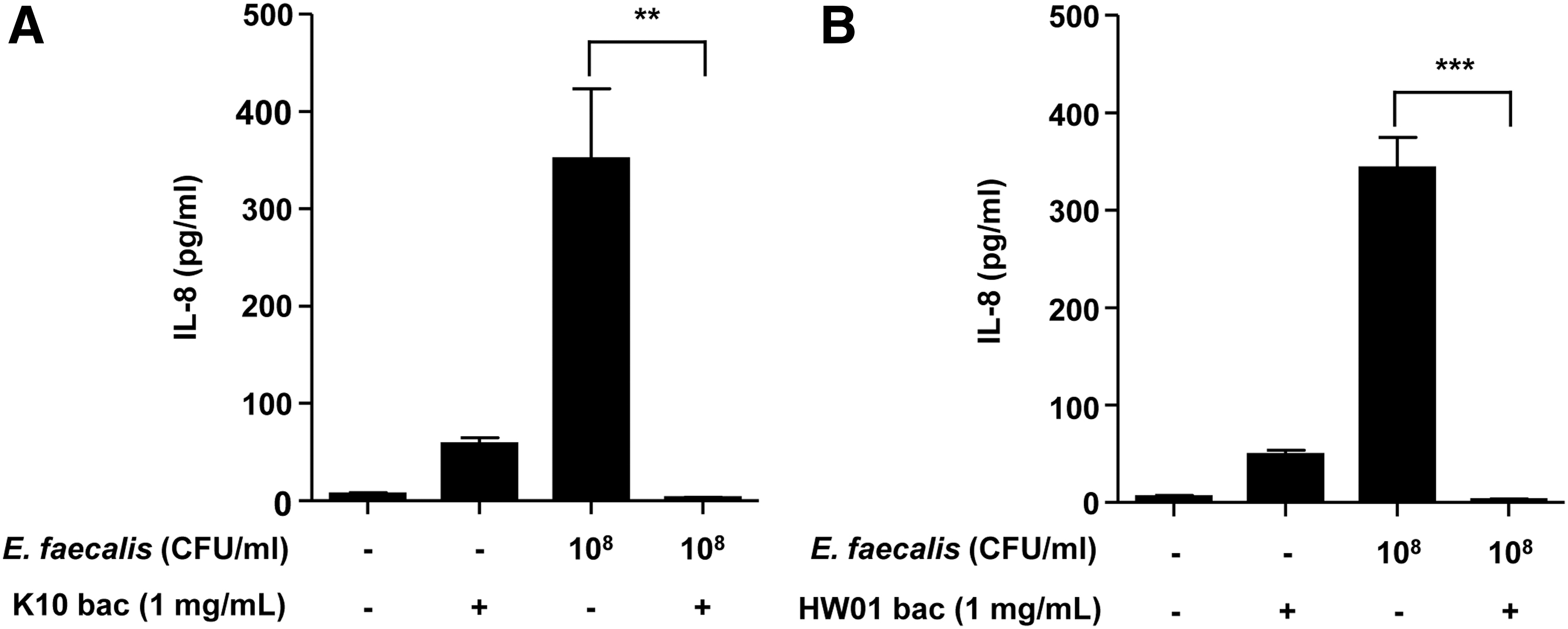
Inhibition of Enterococcus faecalis-induced IL-8 production in Caco-2 cells. Caco-2 cells were incubated with E. faecalis in the presence or absence of K10
In conclusion, based on inhibition of biofilms on SSCs and adhesion inhibition of E. faecalis to Caco-2 cells, the antimicrobial activities of both K10 and HW01 bacteriocins of P. acidilactici may be strain specific. Although molecular studies would be needed to establish the precise mechanism(s) of the reduced biofilm formation, it was demonstrated that biofilm formation is associated with EPS production (Kim and Park, 2013). This suggests that reduced E. faecalis biofilm formation also resulted from decreased EPS production by the P. acidilactici bacteriocin. In addition, this study showed that the P. acidilactici bacteriocin also inhibits the E. faecalis-induced inflammatory response in intestinal epithelial cells in vitro. Taken together, this study suggests that the P. acidilactici bacteriocin could be a promising antimicrobial agent for prevention of E. faecalis infection by reducing biofilm formation.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported by a grant from the National Research Foundation of Korea, which is funded by the Korean government (NRF-2020R1A2C1010010).