
Abstract
Shiga toxin-producing Escherichia coli (STEC) are major foodborne pathogens and seven serogroups, O26, O45, O103, O111, O121, O145, and O157, that account for the majority of the STEC-associated illness in humans. Similar to cattle, swine also harbor STEC and shed them in the feces and can be a source of human STEC infections. Information on the prevalence of STEC in swine feces is limited. Therefore, our objective was to utilize polymerase chain reaction (PCR) assays to determine prevalence of major virulence genes and serogroups of STEC. Fecal samples (n = 598), collected from finisher pigs within 3 weeks before marketing in 10 pig flows located in 8 states, were included in the study. Samples enriched in E. coli broth were subjected to a real-time PCR assay targeting three virulence genes, Shiga toxin 1 (stx1), Shiga toxin 2 (stx2), and intimin (eae), which encode for Shiga toxins 1 and 2, and intimin, respectively. A novel PCR assay was designed and validated to detect serogroups, O8, O20, O59, O86, O91, O100, O120, and O174, previously reported to be commonly present in swine feces. In addition, enriched fecal samples positive for Shiga toxin genes were subjected to a multiplex PCR assay targeting O26, O45, O103, O104, O111, O121, O145, and O157 serogroups implicated in human clinical infections. Of the 598 fecal samples tested by real-time PCR, 25.9%, 65.1%, and 67% were positive for stx1, stx2, and eae, respectively. The novel eight-plex PCR assay indicated the predominant prevalence of O8 (88.6%), O86 (35.5%), O174 (24.1%), O100 (20.2%), and O91 (15.6%) serogroups. Among the seven serogroups relevant to human infections, three serogroups, O121 (17.6%), O157 (14%), and O26 (11%) were predominant. PCR-based detection indicated high prevalence of Shiga toxin genes and serogroups that are known to carry Shiga toxin genes, including serogroups commonly prevalent in cattle feces and implicated in human infections and in edema disease in swine.
Introduction
Shiga toxin-producing Escherichia coli (STEC) reside in the hindgut of food animals and are shed in the feces, which can be a source of contamination of food resulting in foodborne illnesses in humans (Gyles, 2007). STEC infections can range from mild to bloody diarrhea (hemorrhagic colitis) to renal failure (hemolytic uremic syndrome) and even death (Karmali, 1989; Karmali et al., 2010). Shiga toxins (Stx), which are secreted proteins and cytotoxic because of inhibition of protein synthesis (Tesh and O'Brien, 1991), are major virulence factors. There are two types of Shiga toxins, Stx1 and Stx2, encoded by Shiga toxin 1 (stx1) and Shiga toxin 2 (stx2) genes, respectively, which are carried on a prophage (Unkmeir and Schmidt, 2000). Stx2 is more cytotoxic than Stx1, and within each, there are several subtypes (O'Brien et al., 1992). There are three subtypes of stx1: stx1a, stx1c, and stx1d, and seven subtypes of stx2: stx2a, stx2b, stx2c, stx2d, stx2e, stx2f, and stx2g (Scheutz et al., 2012). Another major virulence factor is intimin, a protein encoded by eae, which mediates the attachment of E. coli to host epithelial cells (Kaper et al., 2004). STEC O157:H7 is the most common serotype associated with human infections (Smith et al., 2014; Paletta et al., 2019). However, strains and serotypes belonging to six other serogroups (O26, O45, O103, O111, O121, and O145), commonly referred to as non-O157, are recognized in increased frequency to cause human infections (Brooks et al., 2001; Scallan et al., 2011; Gould et al., 2013). Ruminants are a primary reservoir of STEC, however, STEC do not cause disease in cattle. In contrast, STEC serogroups that produce a subtype of Shiga toxin 2, Stx2e, are pathogenic to pigs and cause a postweaning disease called edema disease (Aarestrup et al., 1997; Frydendahl, 2002).
Swine have been shown to harbor STEC in their gut and shed them in the feces at a similar rate as ruminants in studies reported (Fratamico et al., 2004, 2008; Cha et al., 2018). Although clinically relevant serogroups (O157 and O26) have been reported, a majority of the isolates were of serogroups O8, O59, O71, O86, O100, O163, O174, and O184, with O59 being the most predominant (Cha et al., 2018). The reports on prevalence of serogroups, other than those clinically relevant, were based on random isolation and identification of E. coli because serogroup-specific immunomagnetic beads for isolation, selective media for phenotypic identification based on colony color have not been developed, and serogroup confirmation of putative colonies by multiplex polymerase chain reaction (mPCR) has not been developed and validated. Therefore, information on prevalence of STEC serogroups that are frequently isolated in swine feces is lacking.
There is evidence that swine could be a source of STEC infections in humans. Survey studies done with retail pork meat have shown prevalence of STEC (Doyle and Schoeni, 1987; Read et al., 1990; Samadpour et al., 1994; Brooks et al., 2001). In addition, pork products have been linked to O157 and non-O157 STEC outbreaks (CDC, 1995; Paton et al., 1996; Williams et al., 2000; MacDonald et al., 2004; Conedera et al., 2007; Trotz-Williams et al., 2012; Honish et al., 2017). The primary objectives of our study were to use real-time PCR to determine the prevalence of the three major virulence genes associated with STEC and design and validate a novel eight-plex PCR targeting serogroups O8, O20, O59, O86, O91, O100, O120, and O174 to assess the fecal prevalence of STEC serogroups frequently reported to be present in swine feces. In addition, a mPCR assay was used to identify O26, O45, O103, O111, O121, O145, and O157, which are serogroups most frequently involved in human STEC infections.
Materials and Methods
The protocol for this study was approved by the Kansas State University Institutional Animal Care and Use Committee (IACUC#4197).
Study design and sample collection
A multisite observational and cross-sectional field study was designed and conducted by collecting fecal samples from 10 different pig flows. A flow of pigs is defined as a group of swine farms with pigs raised in age-segregated groups after weaning until they are ready for market and have similar health status, feeding, and management procedures. The selected pig flows were in Iowa, Minnesota, South Dakota, Nebraska, North Carolina, Oklahoma, Kansas, and Ohio. Each pig flow was sampled once. Six different finishing sites (minimum site size = 1000 pigs) from each pig flow were selected and fecal samples from 10 finisher pigs were collected randomly from each site close to marketing the pigs, the earliest being 3 weeks from market. In South Dakota and Iowa, a total of 12 finishing sites from 2 different pig flows were sampled. In most cases, graduate students collected the samples. In one instance (North Carolina), farm personnel collected and shipped the samples. The sample size estimation was based on multisite sampling involving 10 swine operations to detect a positive within each site. The intraclass correlation coefficient, which represents the part of total observations variance due to the between-site variability, was also considered for sample size estimation (Moerbeek et al., 2003; Dohoo et al., 2010). Assuming an estimated prevalence of 5% across sites, our sample size of 60 fecal samples per pig flow will enable us to detect STEC prevalence with a 95% confidence interval. Each fecal sample was collected using a sterile glove directly from the rectum. Fecal samples were transported in coolers with ice to the Kansas State University Pre Harvest Food Safety Laboratory.
Escherichia coli broth enrichment
Approximately 1 g of fecal sample was suspended into 9 mL of Escherichia coli (EC) broth (Difco, ThermoFisher, Waltham, MA) and mixed thoroughly by vortexing for 1 min and the sample suspension was incubated at 40°C for 6 h (Paddock et al., 2012). After incubation, 1 mL of the suspension was pipetted into a 2 mL centrifuge tube, boiled for 10 min in a water bath, centrifuged at 9400 g for 5 min and the DNA in the supernatant was purified by GeneClean Turbo Kit (MP Biomedicals, Solon, OH).
Real-time PCR assay to detect Shiga toxin and intimin genes
The extracted and purified DNA was used to run a triplex real-time PCR assay to detect stx1, stx2, and eae genes. The primers and probes for stx1 and stx2 were from Jacob et al. (2012). The assay conditions were as described by Noll et al. (2015a). In brief, the working concentrations of all primers in a primer mix were 10 pM/μL and of probes were 1.0 pM/μL for stx1, 10.0 pM/μL for stx2, and 10.0 pM/μL for eae. The reaction consisted of 1 μL of primer mix and each probe, 10 μL of Bio-Rad iQ Multiplex Powermix, 4 μL of sterile PCR grade water, and 1 μL of DNA template (total reaction volume = 20 μL). The assay running conditions consisted of 95°C for 10 min, followed by 45 cycles of 95°C for 15 s, 56°C for 20 s, and 72°C for 40 s.
Conventional PCR assay to detect O8, O20, O59, O86, O91, O100, O120, and O174 serogroups
Based on previous studies, eight serogroups, O8, O20, O59, O86, O91, O100, O120, and O174 were considered as the more common STEC in swine feces (Frydendahl, 2002; Fratamico et al., 2004, 2008; Cha et al., 2018). An eight-plex PCR assay to detect the predominant serogroups was designed and validated (described below). The extracted and purified DNA from 598 fecal samples was subjected to the 8-plex PCR assay to determine the prevalence of the 8 serogroups.
Design of primers
The wzx gene, which encodes for the flippase required for O-polysaccharide export, was used to design primers for serogroups O86, O91, O120, O174, O20, and O100. The wbs gene was used to design serogroup O59. The primers for orf469 gene in O8 were from Li et al. (2010). Primers were designed to match all available sequences for the respective serogroups and to amplify the targets with distinct amplicon sizes (Table 1).
Gene Targets, Primer Sequences, and Amplicon Sizes in the Multiplex Polymerase Chain Reaction Assay Developed to Detect Top-8 Serogroups of Shiga Toxin-Producing Escherichia coli in Swine Feces
E. coli strains
Strains belonging to the eight serogroups were used for the initial assay development, validation, and to determine detection sensitivity. Strains O86 and O120 were provided by E. coli Reference Center, Pennsylvania State University, University Park, PA; strains 5380 (O59), 6129 (O174), 5481 (O20), and 6395 (O100) were from the FDA STEC culture collection; and strains 2089-2 (O8) and 4162 (O91) were from our collection.
Template DNA preparation
Bacterial cultures, stored in CryoCare beads (Key Scientific Products, Stamford, TX) at −80°C, were streaked on blood agar plates (BAP; Remel, Lenexa, KS) and incubated overnight at 37°C. One or two colonies of each strain were suspended in 1 mL of distilled water and boiled for 10 min. After a quick centrifugation, 2 μL of the supernatant was used as the DNA template. GeneClean DNA Extraction Kit (MP Biomedicals) was used for DNA purification for EC broth-enriched fecal samples.
Assay conditions
All primer stocks were prepared in 1 × TE buffer (Integrated DNA Technologies, Inc., Coralville, IA) at concentrations of 100 pM/μL. Equal volumes and concentrations of the 8-primer pairs (16 primers) were mixed together. One microliter of the primer mix was used in a 20 μL PCR reaction resulting in final primer concentrations of 0.31 μM for each primer in the reaction. Each reaction also contained 10 μL of Bio-Rad iQ Multiplex Powermix (without additional supplement), 2 μL of boiled bacterial cell preparation or DNA extracted from fecal, spiked fecal, or enriched fecal samples, and 7 μL nuclease-free water. The PCR amplification program included a 5 min denaturation at 94°C, followed by 25 (for pure culture) or 35 cycles (for all others) of 94°C for 30 s, 69°C for 30 s, and 68°C for 75 s. The PCR products were run on QIAxcel Advanced (Qiagen, Germantown, MD) and data were analyzed on QIAxcel ScreenGel software version 1.0.2.
Assay sensitivity with pure cultures and in swine fecal samples spiked with pure cultures
Single colonies of the eight strains from BAP were cultured overnight in Luria Bertani (LB) broth (Becton Dickinson, Sparks, MD) individually, and 100 μL was inoculated into 10 mL LB broth and incubated at 37°C for 3 to 5 h until they reached an absorbance of 0.5 at 600 nm (∼108 CFU/mL). Each culture was serially diluted 10-fold, and dilutions of 10−5, 10−6, and 10−7 were spread-plated on MacConkey agar to determine bacterial cell concentrations (CFU/mL). To determine the sensitivity of the assay with pure cultures, the undiluted cultures of each strain were mixed in equal volumes and six 10-fold dilutions (10−1 to 10−6) were made, and 1 mL of each dilution was boiled for 10 min, centrifuged, and 2 μL of the supernatant was used in the PCR assay. To determine the sensitivity of the assay in fecal sample, 0.1 mL of undiluted culture of each strain was added individually to 9.2 mL EC broth suspended with a fecal sample determined to be PCR negative for the eight serogroups. The fecal suspension spiked with pure cultures were thoroughly mixed and six 10-fold dilutions (10−1 to 10−6) were made. Fecal suspension without the bacterial inoculum served as the negative control. The different dilutions of inoculated fecal sample suspensions and the uninoculated negative control were incubated for 6 h at 40°C. DNA was extracted from inoculated and negative control fecal samples before and after enrichment. The experiments to determine assay sensitivity with pure cultures and with fecal samples spiked with pure cultures were done twice.
Conventional PCR assay to detect O26, O45, O103, O104, O111, O121, O145, and O157 serogroups
Enriched fecal samples that were positive for stx1 or stx2 gene were then subjected to conventional mPCR assay to detect eight serogroups (O26, O45, O103, O104, O111, O121, O145, and O157) and three virulence genes (stx1, stx2, and eae) (Bai et al., 2012; Shridhar et al., 2016).
Statistical analysis
The statistical analysis was performed using STATA v 16.0 (Stata Corp, College Station, TX). Data were considered multilevel in nature with site nested within each pig flow. Bivariate descriptive statistics of prevalence of stx1, stx2, eae, and serogroup-specific genes were assessed by pig flow prior. The cumulative prevalence along with their 95% confidence intervals were calculated at the sample level across all 10 pig flows in 8 states as the proportion of samples that were positive for each virulence gene and serogroup divided by the total number of samples tested (n = 598).
Results
Prevalence of Shiga toxin and intimin genes in feces
Based on the real-time PCR assay, 70.1% (419/598, 95% CI: 66–74) of swine fecal samples were positive and 29.9% (179/598) were negative for stx1 or stx2 gene (Fig. 1). Only 20.9% (125/598, 95% CI: 17.6–24.2) of fecal samples were positive for both Shiga toxin genes. Among the pig flows tested, the prevalence of stx1 or stx2 ranged from 30% to 98.3% (Table 2). The overall prevalence of eae was 66.6% (398/598, 95% CI: 63–70) (Fig. 1) and ranged from 33.3% to 95% among the pig flows sampled (Table 2).

Prevalence of three major virulence genes (mean and 95% confidence intervals) that code for Shiga toxin 1, Shiga toxin 2, and intimin of Shiga toxin-producing Escherichia coli determined by a real-time PCR assay in swine feces (n = 598). eae, intimin; PCR, polymerase chain reaction; stx1, Shiga toxin 1; stx2, Shiga toxin 2.
Prevalence of Genes Encoding Major Virulence Factors, Shiga Toxin 1 (stx1), Shiga Toxin 2 (stx2), and Intimin (eae), of Shiga Toxin-Producing Escherichia coli in Swine Feces (n = 598) Based on a Triplex Real-Time Polymerase Chain Reaction Assay
eae, intimin; stx1, Shiga toxin 1; stx2, Shiga toxin 2.
mPCR assay to detect the eight common swine STEC serogroups
Specificity of the assay with pure cultures
The specificity of each primer pairs was tested individually using the mixture of the eight E. coli serogroups, O8, O20, O59, O86, O91, O100, O120, and O174. The primers only amplified the corresponding serogroup. After optimizing PCR conditions (stated in the Materials and Methods section), eight distinct bands were obtained. The size of the amplicons (kbp) were 562 (O86), 448 (O8), 388 (O59), 352 (O120), 317 (O174), 277 (O91), 238 (O20), and 193 (O100) (Fig. 2).
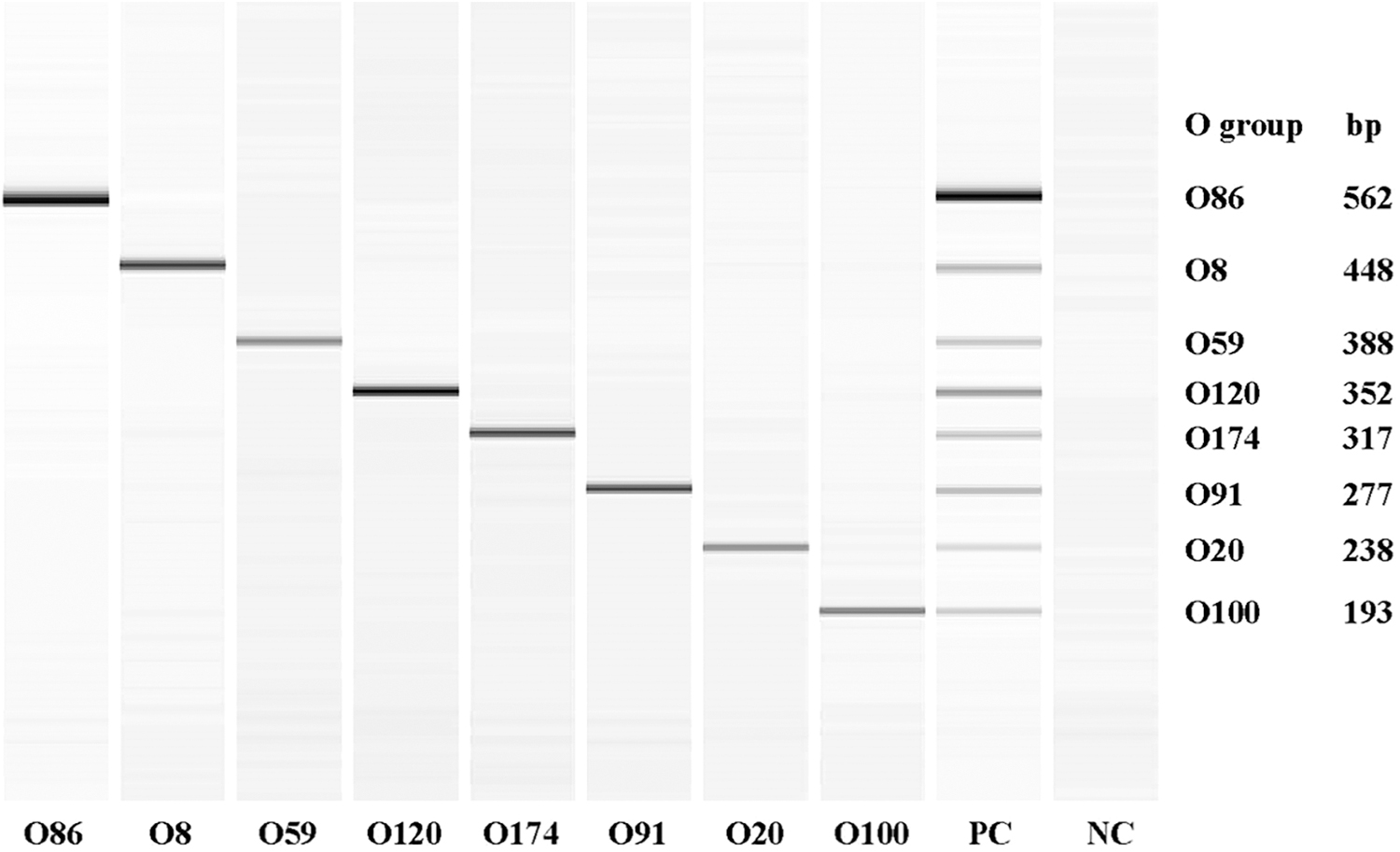
QIAxcel image of the amplicons of serogroup-specific genes of eight individual and pooled Shiga toxin-producing Escherichia coli amplified by eight-plex PCR assay. NC, negative control; PC, positive control (mixture of all eight serogroups); PCR, polymerase chain reaction.
Sensitivity of the assay with pure cultures and fecal samples spiked with pure cultures
The concentrations of eight strains grown individually ranged from 2.2 to 4.5 × 108 CFU (colony-forming unit)/mL (average of two experiments). The minimum concentration of the pooled culture that amplified all eight genes was 2.3 × 103 CFU/mL. In fecal samples spiked with different concentrations of the pooled cultures of the eight serogroups, the sensitivity of detection was 1.8 × 104 CFU/g before enrichment and 1.8 × 102 CFU/g after enrichment for 6 h in EC broth.
Prevalence of O8, O20, O59, O86, O91, O100, O120, and O174 STEC serogroups in feces
A total of 548 (91.6%) of the 598 fecal samples tested were positive for 1 or more of the 8 serogroups tested by the novel mPCR assay. The prevalence of the common STEC serogroups detected in swine fecal samples is shown in Figure 3. Serogroup O8 was the most predominant with a prevalence of 88.6% (530/598, 95% CI:86–91.1), followed by O86 at 35.5% (212/598, 95% CI: 31.6–39.2), O174 at 24.1% (144/598, 95% CI: 20.6–27.5), O100 at 20.2% (121/598, 95% CI: 17–23.4), and O91 at 15.6% (93/598, 95% CI: 12.6–18.5). All 10 pig flows from 8 states had fecal samples positive for serogroups O8 and O86 (Table 3). The prevalence of O20, O120, and O59 were less than 5% of the samples tested. A majority of the serogroup-positive 548 samples contained 1 to 3 serogroups (89.1%), and none was positive for all 8 serogroups (Table 4).
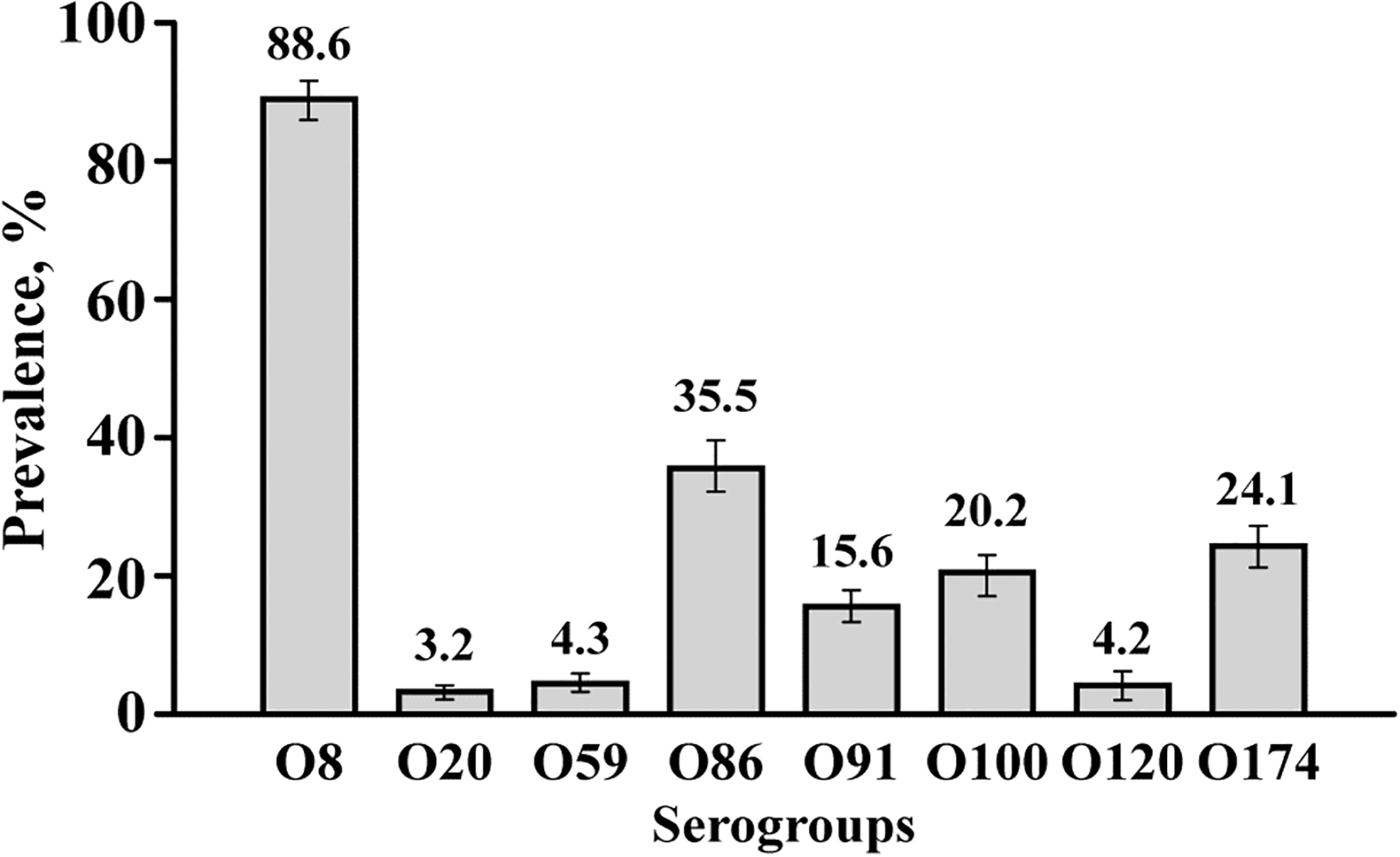
Prevalence of the common serogroups (O8, O20, O59, O86, O91, O120, and O174) of Shiga toxin-producing Escherichia coli (mean and 95% confidence intervals) determined by eight-plex PCR assay in swine feces (n = 598). PCR, polymerase chain reaction.
Prevalence of Serogroups O8, O20, O59, O86, O91, O100, O120, and O174 of Shiga Toxin-producing Escherichia coli in Swine Feces (n = 598) Based on the Eight-Plex Polymerase Chain Reaction Assay
Number and Percentage of Samples Positive for One or More of the Eight Common Serogroups of Shiga Toxin-Producing Escherichia coli in Swine Fecal Samples (n = 598) Based on the Eight-Plex Polymerase Chain Reaction Assay
Prevalence of O26, O45, O103, O104, O111, O121, O145, and O157 serogroups in feces
A total of 225 (49.1%) fecal samples were positive for one or more top-7 and O104 serogroups by the mPCR assay. The prevalence of the eight serogroups detected in swine fecal samples is shown in Figure 4. Of the top-7 STEC, O121 (17.6%, 95% CI: 14.5–20.6) was the predominant serogroup, followed by O157 (14.1%, 95% CI: 11.3–16.8) and O26 (11%, 95% CI: 8.5–13.5). However, serogroup O121 was not detected in fecal samples from the pig flow in Oklahoma (Table 5). All 10 pig flows from 8 states had fecal samples positive for serogroups O26 and O157 (Table 5). Only two fecal samples, one from Iowa and another from South Dakota, were positive for serogroup O111. The overall prevalence of serogroup O104 was 4.8% (29/598, 95% CI: 3.1–6.6) and ranged from 0% to 20% (Table 5). Although the fecal samples used for mPCR assay were all positive for either stx1 or stx2 by real-time PCR, a small proportion of fecal samples were negative for stx1 or stx2 by the conventional PCR assay. For example, the proportion of fecal samples positive for stx1 and stx2 were 70.1% and 66.1%, by real-time and conventional PCR, respectively. Fecal samples negative for O26, O45, O103, O104, O111, O121, and O157 serogroups, but positive for either stx1 or stx2, were categorized as samples positive for other serogroups of STEC. The overall prevalence of the serogroups, other than the targeted eight, was 31.3% (187/598) and ranged from 1.7% to 71.7% (Table 5). A majority of the mPCR-positive samples (n = 225) had one (64.0%) or two (25.3%) STEC serogroups and none had more than five serogroups in a sample (Table 6).
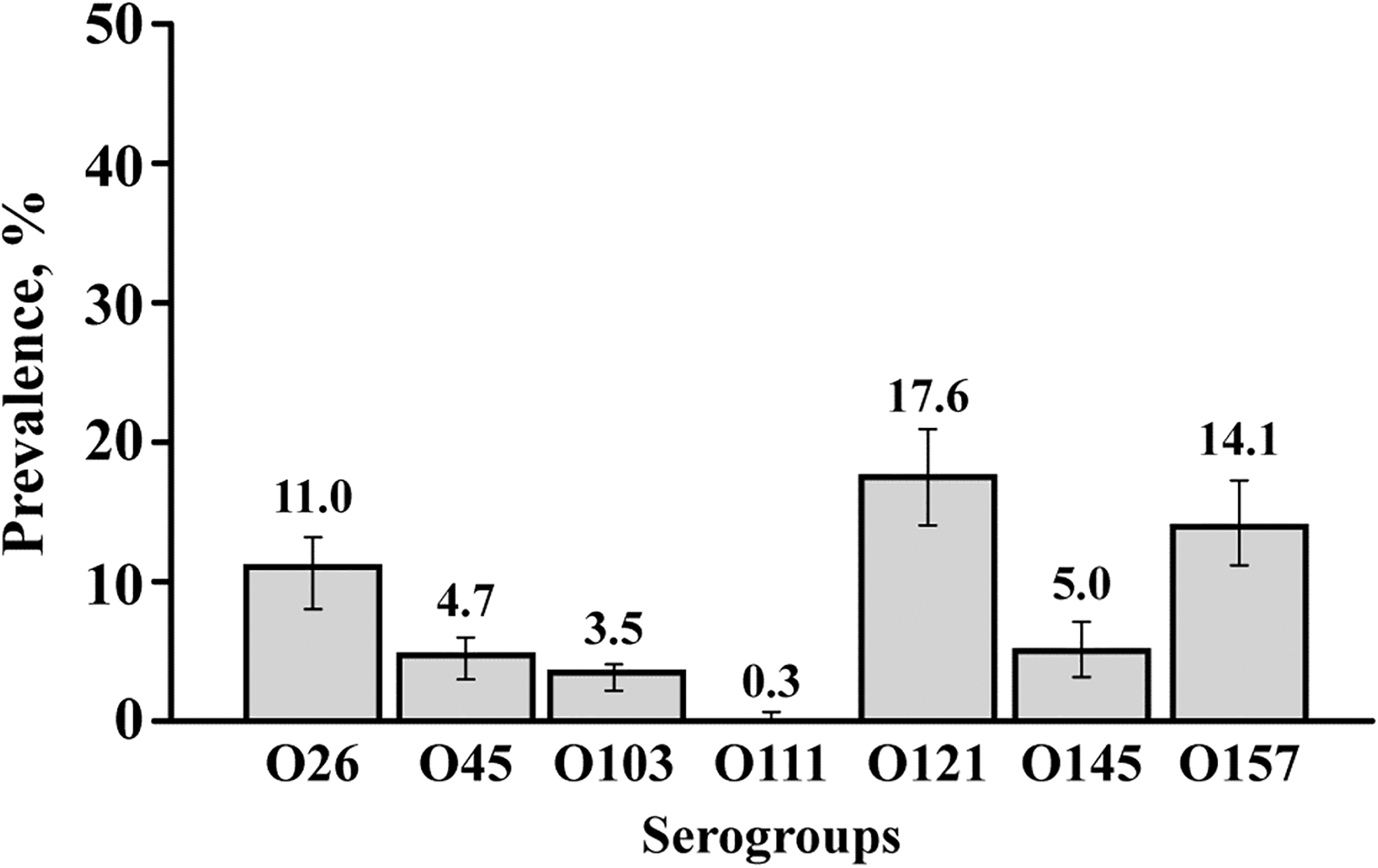
Prevalence of the seven serogroups (O26, O45, O103, O111, O121, O145, and O157) of Shiga toxin-producing Escherichia coli (mean and 95% confidence intervals) determined by multiplex PCR assay in swine feces (n = 598). PCR, polymerase chain reaction.
Prevalence of Serogroups O26, O45, O103, O104, O111, O121, O145, and O157 and Three Major Virulence Genes, stx1, stx2, and eae of Shiga Toxin-Producing Escherichia coli in Swine Feces (n = 598) Based on a Multiplex Polymerase Chain Reaction Assay
Include samples that were positive for stx1 or stx2, but negative for the eight targeted serogroups.
eae, intimin; stx1, Shiga toxin 1; stx2, Shiga toxin 2.
Number and Percentage of Samples Positive for One or More of Top-7 Shiga Toxin-Producing Escherichia coli Serogroups in Swine Fecal Samples (n = 598) Based on Multiplex Polymerase Chain Reaction Assay
Discussion
In this study, we determined by PCR the prevalence in the feces of finishing pigs of three major virulence genes (stx1, stx2, and eae), eight serogroups (O8, O20, O59, O86, O91, O100, O120, and O174), frequently isolated from swine feces, and eight serogroups (O26, O45, O103, O104, O111, O121, O145, and O157), which include the top-7 serogroups involved in human clinical infections. The real-time PCR assay for the three major virulence genes indicated that a high proportion of fecal samples was positive for one or both Shiga toxin genes (70.1%), and stx2 was the predominant Shiga toxin gene, which are similar to prevalence of the virulence genes in cattle feces (Cernicchiaro et al., 2013; Dargatz et al., 2013). The prevalence of Shiga toxin genes in feces determined by conventional PCR were slightly lower (66.1% vs. 70.1%), which is likely because real-time is generally considered to be more sensitive than conventional PCR (Noll et al., 2015a).
Previous studies that have isolated STEC from clinically healthy pigs have reported occurrence of a number of serogroups other than the seven serogroups implicated in human STEC infections. Some of the serogroups detected in swine feces included O8, O9, O20, O26, O59, O71, O86, O100, O128, O163, O174, and O184 (Fratamico et al., 2004; Wells et al., 2012; Meng et al., 2014; Tseng et al., 2014a; Cha et al., 2018; Peng et al., 2019). Because immunomagnetic beads are not available for serogroups other than those clinically relevant to humans and selective media have not been designed for phenotypic identification, detections of the swine STEC serogroups are generally based on culture methods involving isolation of putative E. coli colonies from nonselective media and determining Shiga toxin genes by PCR and serogroups by serology. The ability to detect and isolate different STEC serogroups from fecal or meat are affected by the isolation protocols, particularly the type of enrichment and culture media utilized (Tillman et al., 2012; Verhaegen et al., 2015; Cha et al., 2018). Therefore, PCR-based method has the distinct advantage of detection without selectivity for one serogroup over the other. To estimate the prevalence of the frequently detected STEC serogroups (O8, O20, O59, O86, O91, O100, O120, and O174) in swine feces (Frydendahl, 2002; Fratamico et al., 2004, 2008; Cha et al., 2018; Peng et al., 2019), a novel mPCR assay was designed and validated. The primers in our study were designed to obtain eight amplicons that formed distinct bands. The designed primers were specific for the eight serogroups, based on specificity assays determined with pure cultures of a number of STEC. The specificity and sensitivity of the assay to detect the eight serogroups were also evaluated with fecal samples inoculated with the pooled mixture of the eight STEC. As expected, the sensitivity of the assay was improved with a 6-h enrichment step in EC broth.
Of the eight serogroups, previously reported to common in swine feces, the serogroups with highest prevalence was O8 (88.6%), O86 (35.5%), and O174 (24.1%). This agrees with previous studies (Fratamico et al., 2004, 2008; Kaufmann et al., 2006; Cha et al., 2018) that have concluded that serogroup O8 was the most common serogroup in swine feces. Although clinically healthy pigs carry STEC and shed them in the feces, strains belonging to certain serogroups, particularly O8, O138, O139, O141, and O147, are more frequently associated with the edema disease (Fairbrother and Gyles, 2012; Cha et al., 2018).
The 7 STEC serogroups, clinically relevant to human infections, were present in all 10 pig flows in 8 states at varying proportions and the prevalence of serogroup O121 was the highest (17.6%). The predominance of O121 agrees with previous studies on STEC isolations from swine feces in the United States (Fratamico et al., 2004, 2008; Cha et al., 2018). However, STEC O121 is not the dominant serogroup in swine in other countries in Europe, Asia, or South America (Kaufmann et al., 2006; Beutin et al., 2008; Baranzoni et al., 2016; Colello et al., 2016). The high prevalence of O121 among the non-O157 in swine feces in the United States is in contrast with the prevalence in cattle, in which O26 or O103 are the dominant serogroups (Kalchayanad et al., 2013; Cernicchiaro et al., 2014; Dewsbury et al., 2015; Noll et al., 2015b, 2018; Cull et al., 2017; Valilis et al., 2018). Interestingly, none of the reported STEC infections linked to swine or pork products has implicated serogroup O121, although the serogroup has been isolated from samples collected from pig carcasses (Colello et al., 2016). E. coli O157 was the second most prevalent top-7 serogroup in this study. Although in the initial 2000 NAHMS study, no O157 was isolated from swine feces (Fratamico et al., 2004), studies that specifically targeted O157 with immunomagnetic beads and plating on selective medium have identified O157 (Schierack et al., 2006; Feder et al., 2007; Vu-Khac et al., 2007). The presence of O157:H7 serotype in swine feces has been reported from other countries (Nakazawa and Akkiba, 1999; Rios et al., 1999; Johnsen et al., 2001; Botteldoorn et al., 2003). The dominance of O157 (11.5%) observed in this study is in contrast with previous studies of low prevalence (Frydendahl, 2002; Bonardi et al., 2003; Feder et al., 2007; Tseng et al., 2014b; Colello et al., 2016). Pork products linked to outbreaks have more often been associated with O157 (Williams et al., 2000; MacDonald et al., 2004; Conedera et al., 2007; Trotz-Williams et al., 2012) than non-O157serogroups (Paton et al., 1996).
The mPCR assay, designed and validated to detect the top-7 serogroups and three virulence genes in cattle feces, also included serogroup O104 (Shridhar et al., 2016) because serotype O104:H4, a hybrid pathotype of STEC and enteroaggregative E. coli, was involved in a major foodborne outbreak in Germany in 2011 (Bielaszewska et al., 2006). Although cattle were shown to harbor serogroup O104, with H7 being the major serotype, none of the isolates possessed traits characteristic of the enteroaggregative E. coli (Paddock et al., 2013; Shridhar et al., 2016). The present study indicates that pigs also harbor serogroup O104 and whether the isolates possess any of enteroaggregative traits will require isolation and molecular characterization.
Conclusions
PCR-based detection indicated high prevalence of Shiga toxin genes and serogroups that are known to carry Shiga toxin genes, including serogroups commonly prevalent in cattle feces and implicated in human infections and serogroups implicated in edema disease in swine. Although cattle are a major reservoir of STEC, our study shows swine harbor a number of serogroups of STEC. No culture method has been developed for selective detection and isolation of the predominant STEC serogroups because of lack of serogroup-specific immunomagnetic separation beads and selective media. Therefore, PCR assay is the only available method to estimate the prevalence. However, the limitations of the PCR method is that it detects serogroup and not the serotype, and more importantly, Shiga toxin genes present may or may not be associated with the serogroups detected in the same sample. We are unaware of any published report on the fecal prevalence of the serogroups of E. coli that are known to carry Shiga toxin genes in swine, including serogroups implicated in edema disease in swine. Therefore, a culture method of detection and isolation is needed to identify the serogroups that carry stx genes and subsequent subtyping of stx genes to assess their public health importance and potential to cause human infections and edema disease in swine.
Footnotes
Acknowledgments
The publication is contribution number 20-181-J from the Kansas Agricultural Experiment Station, Manhattan, KS. Authors thank Mal Hoover for her assistance with the images.
Disclosure Statement
No competing financial interests exist.
Funding information
This material is based upon the work that was partially funded by the National Pork Board grant #18–057.