
Abstract
Salmonella enterica serovar Typhimurium is a pathogen harbored by livestock and shed in their feces, which serves as an acquisition source for adult house flies. This study used a green fluorescent protein (GFP) expressing strain of Salmonella Typhimurium to assess its acquisition by and survival within house flies, and transmission from and between flies in the presence or absence of cantaloupe. Female house flies were exposed to manure inoculated with either sterile phosphate-buffered saline or GFP-Salmonella Typhimurium for 12 h, then used in four experiments each performed over 24 h. Experiment 1 assessed the survival of GFP-Salmonella Typhimurium within inoculated flies. Experiment 2 determined transmission of GFP-Salmonella Typhimurium from inoculated flies to cantaloupe. Experiment 3 assessed fly acquisition of GFP-Salmonella Typhimurium from inoculated cantaloupe. Experiment 4 evaluated transmission of GFP-Salmonella Typhimurium between inoculated flies and uninoculated flies in the presence and absence of cantaloupe. GFP-Salmonella Typhimurium survived in inoculated flies but bacterial abundance decreased between 0 and 6 h without cantaloupe present and between 0 and 6 h and 6 and 24 h with cantaloupe present. Uninoculated flies acquired GFP-Salmonella Typhimurium from inoculated cantaloupe and bacterial abundance increased in cantaloupe and flies from 6 to 24 h. More uninoculated flies exposed to inoculated flies acquired GFP-Salmonella Typhimurium when cantaloupe was present than when absent. We infer that the presence of a shared food source facilitated the transfer of GFP-Salmonella Typhimurium from inoculated to uninoculated flies. Our study demonstrated that house flies acquired, harbored, and excreted viable GFP-Salmonella Typhimurium and transferred bacteria to food and each other. Understanding the dynamics of bacterial acquisition and transmission of bacteria between flies and food helps in assessing the risk flies pose to food safety and human health.
House flies are synanthropic nuisance pests that pose a risk to human and animal health (Moon, 2019). Adult flies associate with animal excrement containing abundant microbial communities and human food items, raising public health and food safety concerns (Graczyk et al., 2001; Gupta et al., 2012; Nayduch and Burrus, 2017). Flies harbor bacteria externally on their body surface and internally after direct ingestion or by self-grooming (Jacques et al., 2017; Nayduch and Burrus, 2017). Flies excrete ingested bacteria by regurgitation or defecation (Nayduch et al., 2018). House flies can fly distances as far as 30 km (Baldacchino et al., 2017), emphasizing their role as long-range bridges between sanitary and unsanitary sites.
Salmonella enterica serovar Typhimurium is a bacterial pathogen that causes mild to severe gastrointestinal illness in humans (Andino and Hanning, 2015). This pathogen colonizes the alimentary canal of livestock and is shed in their feces (Himathongkham et al., 1999; Holt et al., 2007). We previously demonstrated that Salmonella Typhimurium persisted in the house fly digestive tract for at least 24 h postingestion (Chifanzwa and Nayduch, 2018) and was acquired from inoculated cattle manure (Thomson et al., 2017). Furthermore, viable Salmonella Typhimurium was excreted by house flies, implicating them as potential vectors (Nayduch et al., 2018).
The gregarious nature of house flies and behaviors such as allo-coprophagy and co-feeding provide opportunities for transmitting pathogens from flies to human food items and among flies (West, 1951; Hanski, 1987). Several studies have described the potential of house flies to contaminate dairy and meat products with foodborne pathogens (Nayduch et al., 2002; De Jesús et al., 2004; Macovei et al., 2008; Pace et al., 2017), but very few have focused on their potential to contaminate fruits, which are attractive to flies because of their high sugar content (Greenberg, 1959; Moon, 2019). Because Salmonella Typhimurium can survive and grow on fruits such as cantaloupe (Chimbombi, 2010; Huang et al., 2015; Bennett et al., 2018), the presence of fruit may facilitate bacterial acquisition and transmission between house flies.
Our study aimed to determine if green fluorescent protein (GFP)-Salmonella Typhimurium (1) persisted in house flies, (2) was transferred from flies to cantaloupe, (3) was acquired from cantaloupe by flies, and (4) was transferred among house flies in the absence and presence of a shared food source (cantaloupe). We hypothesized that GFP-Salmonella Typhimurium would survive in house flies and that flies could transfer the bacteria to cantaloupe. Furthermore, we hypothesized that cantaloupe would serve as a suitable substrate for GFP-Salmonella Typhimurium growth, that uninoculated flies would acquire GFP-Salmonella Typhimurium from contaminated cantaloupe, and that GFP-Salmonella Typhimurium would be transferred among flies but that the presence of cantaloupe would facilitate fly-to-fly transmission.
Materials and Methods
House flies
Flies in this study were reared on cattle manure since 2014 and originated from the parent colony at Kansas State University (KSU) (Thomson et al., 2017). Adult flies were provided sugar and water ad libitum. Egg powder was provided the day before an experiment to induce vitellogenesis and all flies used in the experiments were 5- to 7-d old gravid females.
GFP-Salmonella Typhimurium
A GFP expressing strain of Salmonella Typhimurium SR-11 conferring kanamycin and ampicillin resistance was used in this study (Chifanzwa and Nayduch, 2018). Bacterial cultures were incubated at 37°C in either tryptic soy broth (TSB) or agar (TSA) containing antibiotics (50 μg/mL w/v each of ampicillin sodium [Amp] and kanamycin sulfate [Kan]; Fisher Scientific, Atlanta, GA). A single GFP-Salmonella Typhimurium colony was added to 100 μL of TSB Amp/Kan and incubated for 19 h at 60 rpm to generate the seed culture. For each experiment, GFP-Salmonella Typhimurium inoculum was prepared by transferring 75 μL of seed culture into 15 mL TSB and incubating for 1 h at 37°C at 100 rpm. The inoculum was transferred to manure as described in the pre-experimental setup. For all enumerations, GFP-Salmonella Typhimurium was cultured on TSA Amp/Kan and plates were incubated at 37°C for 24 h.
Cantaloupe preparation
Precut cantaloupe was purchased from a local grocery store the day before each assay and kept at 4°C overnight. Cantaloupe was cut into 10 g pieces before use in Experiments 2–4, hereunder.
Pre-experimental setup: exposure of house flies to manure
Cattle manure was collected from the KSU dairy unit in November 2016. Manure (1.6 kg) was sterilized by autoclaving and individual 200 g aliquots were frozen for later use. For each assay, flies were exposed to 30 g of autoclaved manure that was inoculated with 500 μL of either sterile phosphate-buffered saline (PBS; for control flies) or 5.25 ± 0.72 × 105 colony-forming units (CFU) GFP-Salmonella Typhimurium resuspended in PBS (for Salmonella Typhimurium-inoculated flies) as described in Thomson et al. (2017). Flies acquired bacteria from manure in half-gallon plastic canisters containing a 60 × 15 mm petri dish (Fisherbrand) with inoculated manure and an additional 150 μL droplet of 10% sucrose on parafilm, which was provided as an alternate food source, consistent with our previous study (Thomson et al., 2017). Female flies (n = 25) were added to the canister and allowed to feed ad libitum for 12 h at room temperature (RT; 25–27°C).
Experiment 1: GFP-Salmonella Typhimurium survival in house flies
Female flies were exposed to manure inoculated with GFP-Salmonella Typhimurium, as described previously. After 12 h, 1 g manure and five flies were removed for processing and bacterial culture. To quantify GFP-Salmonella Typhimurium (CFU/g), 1 g manure was suspended in 19 mL of sterile PBS and homogenized by vortexing for 3 min and individual flies were homogenized in 1 mL sterile PBS. Serial dilutions of manure or fly homogenate were plated in duplicate on TSA Amp/Kan and incubated at 37°C for 24 h to enumerate GFP-Salmonella Typhimurium.
To determine the temporal survival of GFP-Salmonella Typhimurium in flies, 15 of the remaining flies were removed and individually placed in separate 32 oz polypropylene jars. Jars were secured with sealing wrap (with small air holes) and maintained at RT. At 6, 12, and 24 h, five jars were chilled and flies (n = 5) were removed, individually homogenized, and processed to culture and enumerate GFP-Salmonella Typhimurium. The number of excreta droplets (regurgitant or feces) in the jar was recorded and the internal jar surface was swabbed to collect the droplets and streak them on TSA Amp/Kan. The experiment was replicated three times.
Experiment 2: transmission of GFP-Salmonella Typhimurium from flies to cantaloupe
Twenty-five flies were exposed to manure inoculated with GFP-Salmonella Typhimurium and after 12 h manure (1 g) and flies (n = 10) were removed and processed for culture and bacterial enumeration. The remaining flies (n = 15) were transferred to individual 32 oz jars and maintained at RT. A 10-g piece of cantaloupe was placed in a small petri dish in each jar. At each time point (6, 12, and 24 h), flies (n = 5) and cantaloupe (n = 5) were removed for processing. Each cantaloupe piece was added to a Stomacher®400 Classic closure bag (177 × 305 mm, 80–400 mL; Seward Limited, Roman Way, United Kingdom) with an equivalent mL/g ratio of sterile PBS and liquefied in a Stomacher400 Circulator (Seward Limited, Roman Way, United Kingdom) at 260 rpm for 10 min. Flies, cantaloupe, and excreta droplets were processed and cultured as described in Experiment 1. The experiment was replicated three times.
Experiment 3: fly acquisition of GFP-Salmonella Typhimurium from inoculated cantaloupe
Two control canisters with n = 25 female flies in each were exposed to manure inoculated with PBS and after 12 h, flies (n = 5) and manure (1 g) were processed from both canisters as in Experiment 1, to confirm absence of GFP-Salmonella Typhimurium. Twenty-four of the remaining flies (equally selected from both canisters) were placed in six 32-oz polypropylene jars (n = 4 flies/jar) containing a 10 g piece of cantaloupe surface inoculated with a 2-μL droplet GFP-Salmonella Typhimurium (mean 165 ± 11 CFU). This amount of bacteria was selected because it fell within the range of GFP-Salmonella Typhimurium that flies excrete (Nayduch et al., 2018) and was determined by suspending 2 μL from the inoculum in 100 μL PBS and then culturing on TSA Amp/Kan. Six jars containing inoculated cantaloupe only (no flies) served as controls. At 6, 12, and 24 h postinoculation, two control and two treatment jars were processed as in Experiment 2 to culture GFP-Salmonella Typhimurium from flies, cantaloupe, and jar swabs of excreta. The experiment was replicated three times.
Experiment 4: transmission of GFP-Salmonella Typhimurium among flies with and without cantaloupe
Twenty-five female house flies were exposed to manure inoculated with GFP-Salmonella Typhimurium (one canister) and 50 females were exposed to manure inoculated with PBS (two control canisters; n = 25/each). After 12 h, manure (1 g) and flies (n = 10) were removed and processed from each canister to culture and enumerate GFP-Salmonella Typhimurium (or verify absence in controls). One GFP-Salmonella Typhimurium-exposed (ST+) fly and four PBS-exposed (ST−) flies were placed into each of six 32 oz polypropylene jars and maintained at RT. A 10-g piece of cantaloupe was placed on a petri dish in three of the six jars while the other three jars contained only the ST+ and ST− flies (no cantaloupe). After 24 h, flies, cantaloupe, and jar swabs were processed for GFP-Salmonella Typhimurium culture and enumeration, as described previously. The experiment was replicated three times.
Statistical analyses
A generalized linear mixed model (GLMM; Proc GLIMMIX in SAS) with a Gaussian distribution, identity link, and restricted pseudo-likelihood estimation was used to determine (1) if GFP-Salmonella Typhimurium abundance varied in flies (Experiment 1) or cantaloupe (Experiment 2) over time and (2) the effect of GFP-Salmonella Typhimurium concentration in cantaloupe on the abundance of GFP-Salmonella Typhimurium in flies over time (Experiment 3). The outcome was the log10-transformed CFU +1 (to account for 0 CFU in the model) of GFP-Salmonella Typhimurium in flies (Experiments 1, 2, and 3) or the log10 CFU +1/g of GFP-Salmonella Typhimurium in cantaloupe (Experiment 2). Independent variables were time, Salmonella Typhimurium concentration in cantaloupe (log10 CFU +1/g), and an interaction of both variables. Trial (replicate) was included as a random intercept (Experiments 1–3).
To compare raw proportions of flies or cantaloupe positive for GFP-Salmonella Typhimurium, a chi-square test was used (Proc FREQ and compprop macro in SAS version 9.4 [SAS Institute, Inc., Cary, NC]; Zar, 1999). GLMMs (using a binomial distribution, logit link, and restricted pseudo-likelihood estimation) were fitted to test the effect of time on the proportion of GFP-Salmonella Typhimurium in flies or cantaloupe. Time (categorical: 0, 6, and 24 h) was included as an independent variable, except for Experiment 4 where time was not a variable, and trial was a random intercept. A linear mixed-effects model with a negative binomial distribution, logit link, and restricted pseudo-likelihood estimation (Experiments 1 and 2) was fitted in SAS to determine the effect of time (fixed effect) on the number of excreta droplets recovered from the jars, where trial was a random intercept. The Tukey–Kramer procedure was used to adjust for multiple comparisons in all GLMMs and p-values (<0.05) were used to determine statistical significance.
Results
Experiment 1: GFP-Salmonella Typhimurium survival in house flies
Overall, there was a significant effect of time on the abundance of GFP-Salmonella Typhimurium in house flies (p < 0.001). GFP-Salmonella Typhimurium abundance significantly decreased in flies from 0 to 6 h postingestion (p < 0.001; Fig. 1a). Overall, time did not significantly affect the proportion of flies positive for GFP-Salmonella Typhimurium (p = 0.580; Fig. 1a); however, the proportion of positive flies at 0 h (15/15) was significantly higher than the proportion of positive flies at other collection times (p = 0.001). There was no significant difference in the number of excreta droplets in jars over time (p = 0.677; Table 1). GFP-Salmonella Typhimurium was recovered from excreta droplets in only 1 of 15 jars (at 12 h; Table 2).
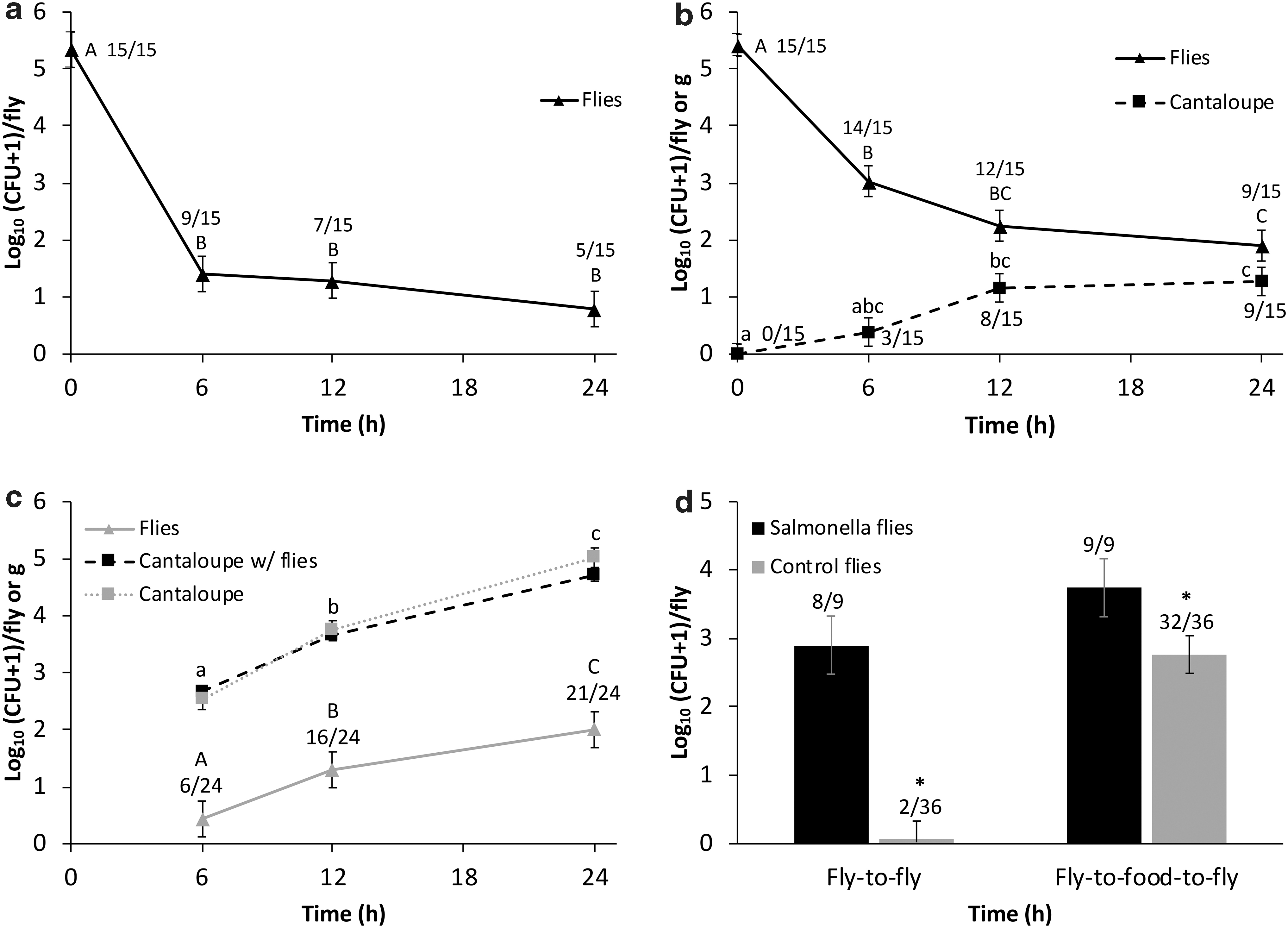
GFP-Salmonella enterica serovar Typhimurium abundance in flies and cantaloupe. Each panel displays results from an experiment:
The Mean Number of Fly Excreta Droplets Recovered from Jars
Mean ± standard error of the mean for n = 15 jars are given. Letters represent significant differences between time points within an experiment (a vs. b p = 0.012; b vs. c p = 0.001; a vs. c p < 0.001).
Proportion of Jars with Excreta Containing Green Fluorescent Protein-Salmonella enterica Serovar Typhimurium
See text for experimental procedures. Proportions represent the number of jars where GFP-Salmonella Typhimurium was cultured from swabs out of the total number of jars sampled at each time point.
GFP, green fluorescent protein; N/A, not applicable.
Experiment 2: transmission of GFP-Salmonella Typhimurium from flies to cantaloupe
There was a significant effect of time on GFP-Salmonella Typhimurium abundance in house flies (p < 0.001), which decreased from 0 to 6 h (p < 0.001) and 0 to 24 h (Fig. 1b; p = 0.022). There was no significant difference between bacterial abundance at 12 and 24 h (p = 0.798). There was no overall effect of time on the proportion of GFP-Salmonella Typhimurium-positive flies (Fig. 1b, p = 0.7845). However, there was an effect of time on GFP-Salmonella Typhimurium concentration in cantaloupe (p < 0.001), which significantly increased from 0 to 12 h (Fig. 1b; p = 0.002). The proportion of cantaloupe positive for GFP-Salmonella Typhimurium did not significantly differ over time (Fig. 1b; p = 0.245). The mean number of excreta droplets in jars changed over time (p < 0.001; Table 1), with an increase from 6 to 12 h (p = 0.012), 12 to 24 h (p = 0.001), and 6 to 24 h (p < 0.001). GFP-Salmonella Typhimurium was recovered from excreta droplets in 5 of 15 jars at both 6 and 12 h and 3 of 15 jars at 24 h (Table 2).
Experiment 3: acquisition of GFP-Salmonella Typhimurium by flies from inoculated cantaloupe
Overall, time affected the abundance of GFP-Salmonella Typhimurium in house flies (p < 0.001), which increased from 6 to 12 h (p = 0.004; Fig. 1c), from 12 to 24 h (p = 0.024), and from 6 to 24 h (p < 0.001). There was a significant effect of time on the proportion of flies positive for GFP-Salmonella Typhimurium (p = 0.011; Fig. 1c), which increased from 6 of 24 flies at 6 h to 21 of 24 flies at 24 h (p = 0.011). Time also significantly affected the concentration of GFP-Salmonella Typhimurium in cantaloupe (p < 0.001), but there was no effect of fly presence (p = 0.465). GFP-Salmonella Typhimurium concentration in control cantaloupe increased from 6 to 12 h (p < 0.001; Fig. 1c) and from 6 to 24 h (p < 0.001). Similarly, GFP-Salmonella Typhimurium concentration in cantaloupe with flies present increased from 6 to 12 h (p < 0.001) and from 6 to 24 h (p < 0.001). Furthermore, the interaction of time and the bacterial concentration in cantaloupe positively affected the abundance of GFP-Salmonella Typhimurium in flies (p = 0.001; Fig. 1c). GFP-Salmonella Typhimurium was recovered from excreta droplets collected from 0 of 4 jars at 6 h, 3 of 4 jars at 12 h, and 4 of 4 jars at 24 h (Table 2). The number of excreta droplets was not recorded because jars contained more than one fly.
Experiment 4: transmission of GFP-Salmonella Typhimurium among flies with and without cantaloupe
At 24 h, there was no significant effect of treatment (cantaloupe presence or absence) on the abundance of GFP-Salmonella Typhimurium in ST+ flies (p = 0.1845; Fig. 1d). However, treatment significantly affected the abundance of GFP-Salmonella Typhimurium in ST− flies (p < 0.001; Fig. 1d). There was no significant difference in the proportion of ST+ flies positive for GFP-Salmonella Typhimurium in the presence or absence of cantaloupe (p = 0.978; Fig. 1d). However, there was a significant difference in the proportion of ST− flies positive for GFP-Salmonella Typhimurium (p = 0.001). Only 2 of 36 ST− flies were positive for GFP-Salmonella Typhimurium without cantaloupe compared with 32 of 36 when cantaloupe was present (p = 0.001; Fig. 1d). No GFP-Salmonella Typhimurium was recovered from excreta swabs from the jars without cantaloupe, whereas excreta from seven of nine jars with cantaloupe were culture positive for GFP-Salmonella Typhimurium (Table 2). The number of excreta droplets was not recorded because there was more than one fly in each jar.
Discussion
We investigated the acquisition and transmission of GFP-Salmonella Typhimurium between house flies and cantaloupe in four experiments. In Experiment 1, the abundance of GFP-Salmonella Typhimurium decreased in flies over time but time did not affect the proportion of positive flies, most likely because of a small sample size. The decrease in bacterial abundance 0–6 h may be attributed to bacterial digestion because no excreta collected from the jars were positive for GFP-Salmonella Typhimurium and there was no change in the number of excreta droplets over time. In contrast to our results, GFP-Salmonella Typhimurium abundance increased within flies 6 h after ingesting inoculated culture broth (Chifanzwa and Nayduch, 2018) and the ingested “dose” of bacteria affected survival. In our study, the amount ingested by each fly was difficult to assess owing to the ad libitum access to manure. Previous studies (McGaughey and Nayduch, 2009; Nayduch et al., 2013; Fleming et al., 2014) have shown excretion of viable bacteria from flies who ingested bacteria in culture media, which may promote proliferation and survival. These studies collectively demonstrate that the acquisition substrate may determine the fate of bacteria in flies.
Experiment 2 assessed the transfer of GFP-Salmonella Typhimurium from flies to cantaloupe. There was a decrease in bacterial abundance in flies and the proportion of flies positive for bacteria over time. The proportion of GFP-Salmonella Typhimurium-positive cantaloupe pieces increased over time indicating that the flies successfully transferred bacteria to cantaloupe. The increase of GFP-Salmonella Typhimurium concentration on cantaloupe over 24 h may be attributable to repeated inoculations from flies and/or growth of bacteria on the cantaloupe. Because GFP-Salmonella Typhimurium concentration in cantaloupe without flies did not differ from that with flies, flies likely did not significantly contribute to the increase in bacterial abundance. Of interest, the abundance of GFP-Salmonella Typhimurium in flies did not decrease as quickly between 0 and 6 h when cantaloupe was available as a food source (Experiment 2) than when flies had no cantaloupe (Experiment 1), indicating that the presence of cantaloupe enhances bacterial survival in the fly and potentially bolsters vector potential.
Experiment 3 assessed the acquisition of GFP-Salmonella Typhimurium from inoculated cantaloupe by flies. There was an increase in abundance of GFP-Salmonella Typhimurium in both flies and cantaloupe over time, which could be attributed to bacterial proliferation on the cantaloupe and recurrent ingestion by flies. The proliferation of GFP-Salmonella Typhimurium on the cantaloupe may produce volatiles that are attractive to flies (Lam et al., 2007), increasing their interest and contact.
To determine whether food facilitated bacterial transmission among flies, Experiment 4 examined the transfer of GFP-Salmonella Typhimurium between one ST+ fly and 4 ST− flies in the presence and absence of cantaloupe. Cantaloupe presence did not affect GFP-Salmonella Typhimurium abundance in ST+ flies or the proportion of ST+ flies positive for bacteria. Of interest, the abundance of GFP-Salmonella Typhimurium in ST+ flies was greater than the abundance of GFP-Salmonella Typhimurium in flies in Experiment 1 (without cantaloupe). The presence of the ST− flies in Experiment 4 may have affected the ST+ fly behavior impacting their digestion or excretion of GFP-Salmonella Typhimurium. House flies spend most of their time resting or regurgitating when they are alone (Barber and Starnes, 1949), but when other flies are present these behaviors may change. Because regurgitation or “bubbling” plays an important role in preparing the meal for digestion (Stoffolano and Haselton, 2013), the gregarious interactions with other flies may have interrupted this behavior and the subsequent digestion of GFP-Salmonella Typhimurium.
Experiment 4 determined that food facilitated the transfer of bacteria from ST+ flies to ST− flies. Both abundance of GFP-Salmonella Typhimurium in ST− flies and proportion of ST− flies positive for GFP-Salmonella Typhimurium were greater when cantaloupe was present than when it was absent. ST+ flies likely contaminated cantaloupe with GFP-Salmonella Typhimurium, which then served as a source of bacteria for ST− flies. When cantaloupe was absent, only 2 of 32 ST− flies were positive for GFP-Salmonella Typhimurium after 24 h compared with 32 of 36 ST− flies acquiring bacteria with cantaloupe present. Of interest, the 4 ST− flies that were negative for GFP-Salmonella Typhimurium at 24 h originated from the same container and although the ST+ fly was positive for GFP-Salmonella Typhimurium, the cantaloupe was not. This observation lends support to the hypothesis that contaminated cantaloupe facilitated GFP-Salmonella Typhimurium transmission between flies.
When cantaloupe is present, fly excreta also may serve as an indirect source of GFP-Salmonella Typhimurium for flies. No change in the number of excreta droplets over time was observed in jars without cantaloupe (Experiment 1), and more excreta were recovered when cantaloupe was present (Experiment 2). In all experiments, more jars were culture positive for GFP-Salmonella Typhimurium when cantaloupe was present than when absent, suggesting that cantaloupe facilitated excretion of viable bacteria. For example, more jars were positive for GFP-Salmonella Typhimurium in Experiment 2 compared with Experiment 1. Similarly, in Experiment 3, all four jars containing cantaloupe inoculated with GFP-Salmonella Typhimurium were positive at 24 h, and in Experiment 4, zero of nine jars without cantaloupe were positive for GFP-Salmonella Typhimurium compared with seven of nine jars with cantaloupe.
This study explored the survival of Salmonella Typhimurium in house flies and the potential for flies to acquire this pathogen from manure or fruit, transmit bacteria to fruit and transfer bacteria to each other through fruit. The presence of cantaloupe facilitated both bacterial excretion from flies and bacterial transfer among flies. Our results emphasize the importance of house fly management in food safety programs implemented at markets, dining facilities, food manufacturing plants, and other environments where human food is openly presented or handled. We demonstrated that the presence of flies and food creates a potential public health issue, where flies both inoculate food with pathogens and create a source of bacteria for other flies. In practical sense, when flies have unrestricted access to both sources of bacteria (e.g., garbage bins) and uncovered food (e.g., buffets, picnics, or salad bars) they can amplify the risk of foodborne pathogen contamination especially when other flies are present. To fully assess their role in food safety, fly transmission of other bacterial pathogens to a variety of foods should be explored.
Footnotes
Acknowledgments
The authors thank A. Cooper for helpful feedback on this article. Methods and results presented here are also available in the PhD dissertation of the first author (
Disclosure Statement
No competing financial interests exist.
Funding Information
This project was funded by USDA-ARS National Program 104, project numbers 3020-32000-007-00D and 3020-32000-018-00D.