
Abstract
Although serotyping is the most important method of identification of taxonomy in Salmonella, conventional serotype determination with a complete set of antisera is time consuming and laborious. Recently, rapid serotyping procedures with polymerase chain reaction (PCR) have been developed. In this study, we established a novel PCR-based rapid serotyping method that employs a unique target gene. Alignment study of Salmonella-specific gene (Salmonella enterotoxin [stn]) revealed a correlation between the stn gene sequence and the serotype of the organism. In 750 bp of stn gene, 55 nucleotides indicated single nucleotide polymorphism (SNP)-like polymorphism, and the correlation between the SNP-like polymorphism and the serotype of the organism suggests that SNP-like sequences in stn gene can serve as an index for serotyping. To develop a rapid serotyping method based on the SNP-like polymorphism, we selected serotype-associated 12 SNP-like sites in the stn gene and established a method based on high-resolution melting (HRM) and PCR, which identifies nucleotides at SNP-like sites within 1.5 h. This newly established rapid serotyping procedure (stn-HRM) could identify nine serotypes, including the frequently isolated serovar Enteritidis. These nine serotypes cover 64.3% of cases of Salmonella, as reported by the World Health Organization/Global Foodborne Infection Network (WHO/GFN) Country Databank from 2001 to 2010. In this study, we employed a unique target gene, stn, which is completely independent of the genes that were targeted in previously reported rapid serotyping procedures. Therefore, the results obtained by our newly developed stn-HRM procedure are independent of the results obtained by other procedures. Besides, stn-HRM can ensure accurate identification of the bacterial species as stn is a Salmonella-specific gene. It is expected that the combination of newly constructed stn-HRM and previously reported procedures could further improve the credibility of Salmonella isolate serotyping.
Introduction
Within the genus Salmonella, more than 2500 serotypes (or serovars) can be differentiated using the Kauffmann–White scheme (Grimont and Weill, 2008). The serotype of genus Salmonella is defined based on the agglutination of organism with antisera against variants of the somatic (O) and flagellar (H) antigens. Although subtyping procedures have been developed for genus Salmonella (Wattiau et al., 2011), serotyping is still widely accepted as a principal differentiation scheme of its subtypes. In a surveillance program conducted by World Health Organization/Global Foodborne Infection Network (WHO/GFN), Salmonella isolation data were associated with their serotypes to control and prevent foodborne and other Salmonella-related infections from farm to table (
Recently, several molecular-based subtyping methods using conventional polymerase chain reaction (PCR), real-time PCR, pulsed-field gel electrophoresis, and multilocus sequencing typing have been developed for Salmonella serotyping (Sukhnanand et al., 2005; Akiba et al., 2011; Rajtak et al., 2011; Zeinzinger et al., 2012; Li et al., 2014; Majchrzak et al., 2014; Masek et al., 2014; Shi et al., 2015). Among these methods, conventional PCR-based subtyping procedures are cost effective and easy to conduct; however, these procedures allow the identification of only a single or a few serotypes simultaneously (Akiba et al., 2011; Li et al., 2014; Majchrzak et al., 2014). In contrast, a recently reported real-time PCR-based procedure allows the identification of more than 30 serotypes within a couple of hours (Zeinzinger et al., 2012). High resolution melting (HRM) is a relatively new and cost-effective PCR-based method, which can be used for single nucleotide polymorphism (SNP) typing (Zhou et al., 2004). The HRM technique has been recently applied to the rapid detection or subtyping of several types of bacteria, including Salmonella species (Yang et al., 2009; Jeng et al., 2012; Zeinzinger et al., 2012). The principle of HRM analysis for SNP is the generation of discriminable melting curve profiles due to sequence variations at SNP sites. Among the several types of HRM, the procedure with an unlabeled oligonucleotide probe was known as one of the most sensitive and cost effective (Zhou et al., 2004).
Salmonella enterotoxin (stn) is the gene characteristically observed in Salmonella species (Chopra et al., 1987). While its function in Salmonella pathogenesis has not been elucidated in detail (Watson et al., 1998; Wallis et al., 1999; Nakano et al., 2012), the PCR primers specific for this gene are generally used for rapid and specific screening of Salmonella species. This is because a 750 bp sequence of stn gene is highly conserved in the Salmonella species (Chopra et al., 1994; Makino et al., 1999; Moore and Feist, 2007). In this study, we analyzed stn gene sequences derived from 391 Salmonella isolates encompassing 51 serotypes and identified specific SNP-like sequences, which can be used as an index for serotyping. To develop a rapid serotyping method based on the SNP-like sequences in the stn gene, we employed HRM for the typing of these SNP-like sites.
Materials and Methods
Bacterial strains
A total of 149 isolates of Salmonella encompassing 24 serotypes were used for stn gene sequencing. These Salmonella strains were isolated from patients with diarrhea or meat samples in the Kingdom of Thailand. They were serotyped using commercially available O and H antisera developed for serotyping of Salmonella (Denka Seiken, Tokyo, Japan) according to the manufacturer's instructions. For validation analysis, 45 Salmonella strains isolated in Japan were independently prepared and serotyped with the commercially available antisera as above.
Analysis of stn gene sequence
PCR primers designed to hybridize to 5′-flanking region (5′-TCGTGCAGTGGCTTACATCATC-3′) or 3′-flanking region (5′-CAAGGGAGTGGCGTAATACGC-3′) of the stn gene (accession no. L16014.1) of Salmonella Typhimurium standard strain LT2 was used to obtain the PCR product containing whole stn gene. The template was prepared according to the following procedure. The isolated colony of Salmonella cultured on Luria-Bertani (LB) agar was suspended in 100 μL of distilled water. The suspension was heated at 100°C for 5 min, and centrifuged at 20,000 × g for 5 min to obtain the supernatant. The supernatant containing the genomic DNA of Salmonella isolates was subjected to PCR. The amplification reaction was performed using the PrimeSTAR Max DNA Polymerase Kit (TaKaRa Bio, Inc., Shiga, Japan) with 30 cycles of optimized conditions (98°C, 10 s; 55°C, 15 s; 72°C, 60 s). The size of the amplified PCR fragment was analyzed by agarose gel electrophoresis using DNA Molecular Marker XIV (Roche Diagnostics, Penzberg, Germany) as a molecular weight marker. The PCR products were purified using the illustra GFX PCR DNA and Gel Band Purification Kit (GE Healthcare, Buckinghamshire, United Kingdom), and sequenced at Hokkaido System Science Co., Ltd. (Hokkaido, Japan) with the ABI PRISM 3130 Genetic Analyzer (Applied Biosystems, Foster City, CA) using the BigDye Terminator v3.1 Cycle Sequencing Kit. All PCR products were prepared and sequenced in triplicate. The obtained nucleotide sequence data are available in the DDBJ/EMBL/GenBank databases under the accession numbers LC553048–LC553196. In addition, 242 stn gene sequences were obtained from nucleotide BLAST searches (
HRM-curve PCR analysis
PCR primers and unlabeled oligonucleotide probes were individually designed to detect SNP-like sequences (Table 1), and the same PCR program was used for the detection of all SNP-like sequences. The template for HRM-PCR analysis was prepared according to the following procedure. The isolated colony of Salmonella cultured on LB agar was suspended into 100 μL of distilled water. The suspension was heated at 100°C for 5 min, and centrifuged at 20,000 × g for 5 min to obtain the supernatant. The supernatant containing the genomic DNA of Salmonella isolates was subjected to asymmetric HRM-PCR analysis with LightCycler 480 High-Resolution Melting Master (Roche Diagnostics) (Zhou et al., 2004). The PCR reaction and HRM analysis were performed on the LightCycler Nano system (Roche Diagnostics), and data were analyzed with LightCycler Nano Software v. 1.1 (Roche Diagnostics). Twenty microliters of reaction mixture consisted of 2.0 μL of extracted genomic DNA, 0.06 or 0.30 μM of primer (Table 1), 0.24 μM of unlabeled oligonucleotide probe, 1 × LightCycler 480 Master Mix, 1.0 μL of dimethylsulfoxide, and 3 mM MgCl2. Assays were carried out with the following touchdown PCR cycling and HRM conditions. The PCR was initiated with a 10 min hold at 95°C, followed by 39 cycles of 95°C for 10 s, a touchdown cycling step (decreasing 1.0°C/cycle) with annealing ranging from 64°C to 59°C for 15 s and 72°C for 10 s. After amplification, the samples were heated to 95°C for 1 min and then cooled to 40°C for 4 min to encourage heteroduplex formation. HRM curve data were obtained by melting over 40°C to 90°C with an integration time of 0.1 s. The Melting peaks plots and Difference plots of each unlabeled oligonucleotide probes were developed by Tm calling analysis and HRM analysis, respectively, with LightCycler Nano Software v. 1.1.
Polymerase Chain Reaction Primers and Unlabeled Oligonucleotide Probes Used in stn High-Resolution Melting
Primer concentration in the stn-HRM analysis.
Nucleotides corresponding to SNP-like sites are underlined. “-P” indicates a 3′-phosphate modification used to block the 3′ end from extension during thermal cycling.
HRM, high-resolution melting; SNP, single nucleotide polymorphism; stn, Salmonella enterotoxin.
Results
Alignment study of stn gene sequences
PCR fragments of the stn gene from 149 Salmonella isolates encompassing 24 serotypes were amplified and sequenced. In addition, 242 stn gene sequences originating from 43 serotypes were obtained from nucleotide BLAST searches. The alignment of a total of 391 stn genes from 51 serotypes revealed that these stn sequences could be fallen into 49 patterns, as shown in Supplementary Figures S1 and S2, and Table 2. In the 750 bp of the stn gene, 695 nucleotides were completely conserved throughout all isolates. On the other hand, 55 nucleotides indicated SNP-like polymorphism (Table 2). Noteworthy, comparing with serotype information, the nucleotides used in these SNP-like sites were almost conserved within each serotype. For example, at position 85, all serovars of Enteritidis, Virchow, Agona, Anatum, Infantis, etc. have guanine while all the serovars of Choleraesuis, Heidelberg, etc. have adenine. In addition, we revealed that the correlation between the pattern of polymorphism in each SNP-like site and serotype vary at different sites. For example, the serovars Enteritidis and Typhimurium have the same nucleotide (guanine) at the position 85 but have different nucleotides at the position 118 (cytosine and adenine for serovars Enteritidis and Typhimurium, respectively). These data indicate the possibility that serotypes can be inferred according to the nucleotide usage at some SNP-like sites in the stn gene.
List of Single Nucleotide Polymorphism-Like Sites in the stn Gene
Accession numbers of stn gene or genome in representative isolates are indicated in parentheses.
Rapid identification of the nucleotides used in each SNP-like site with HRM analysis
To establish the method for estimating the serotype of Salmonella isolate without analysis of whole stn gene sequence, we selected 12 SNP-like sites listed in Table 3 and analyzed the correlation between nucleotide usage pattern at these sites and serotypes. As seen in Table 3, 20 sequence types (STs) marked with superscript letter “b” include single serotypes. These data indicate that these 20 serotypes can be possibly identified by the analysis of selected 12 SNP-like sites.
List of Nucleotides Utilized at Selected 12 Single Nucleotide Polymorphism-Like Sites and Corresponding Serotypes
Figure in parentheses indicates the number of sequences used for the alignment study.
ST with nucleotide usage patterns that appeared only in a single serotype.
stn-HRM analysis for serovar Anatum indicates a unique result due to SNP-like sequence observed only in position 706 of serovar Anatum (see Results for details).
ST, sequence type.
We attempted to develop a rapid determination method of nucleotides utilized in each SNP-like site. In this study, we employed the HRM technique with an unlabeled oligonucleotide probe. For this purpose, we developed 12 sets of PCR primers and unlabeled oligonucleotide probe fully matched with either variant (Table 1) and analyzed melting curve profiles of these probes against SNP-like sites (Fig. 1 and Supplementary Fig. S3). We used the same PCR condition for all 12 SNP-like sites, which facilitates the simultaneous analysis of the 12 SNP-like sites, enabling complete rapid serotyping within 1.5 h. We analyzed our 149 stocks of Salmonella isolates with the newly developed procedure (stn-HRM). Results obtained by stn-HRM were consistent with sequencing results, except for serovar Anatum. In the case of serovar Anatum (ST19), probe for position 712 produced a unique melting curve due to SNP-like sequence at position 706 that was uniquely observed in this serovar (Table 2). The resultant melting curve for position 712 for serovar Anatum was similar to the curve for thymine, even though the serovar Anatum has cytosine at this position. Due to this feature, we identified the serovar Anatum from other serotypes with stn-HRM (ST32 in Table 3). These results demonstrate the possibility that “identifiable” 20 STs (marked with superscript letter “b” in Table 3) together with serovar Anatum can be rapidly identified with the stn-HRM procedure. However, in this study, we did not include Salmonella isolates belonging to some STs; future study with the stn-HRM will be performed against a more comprehensive collection of serotypes.
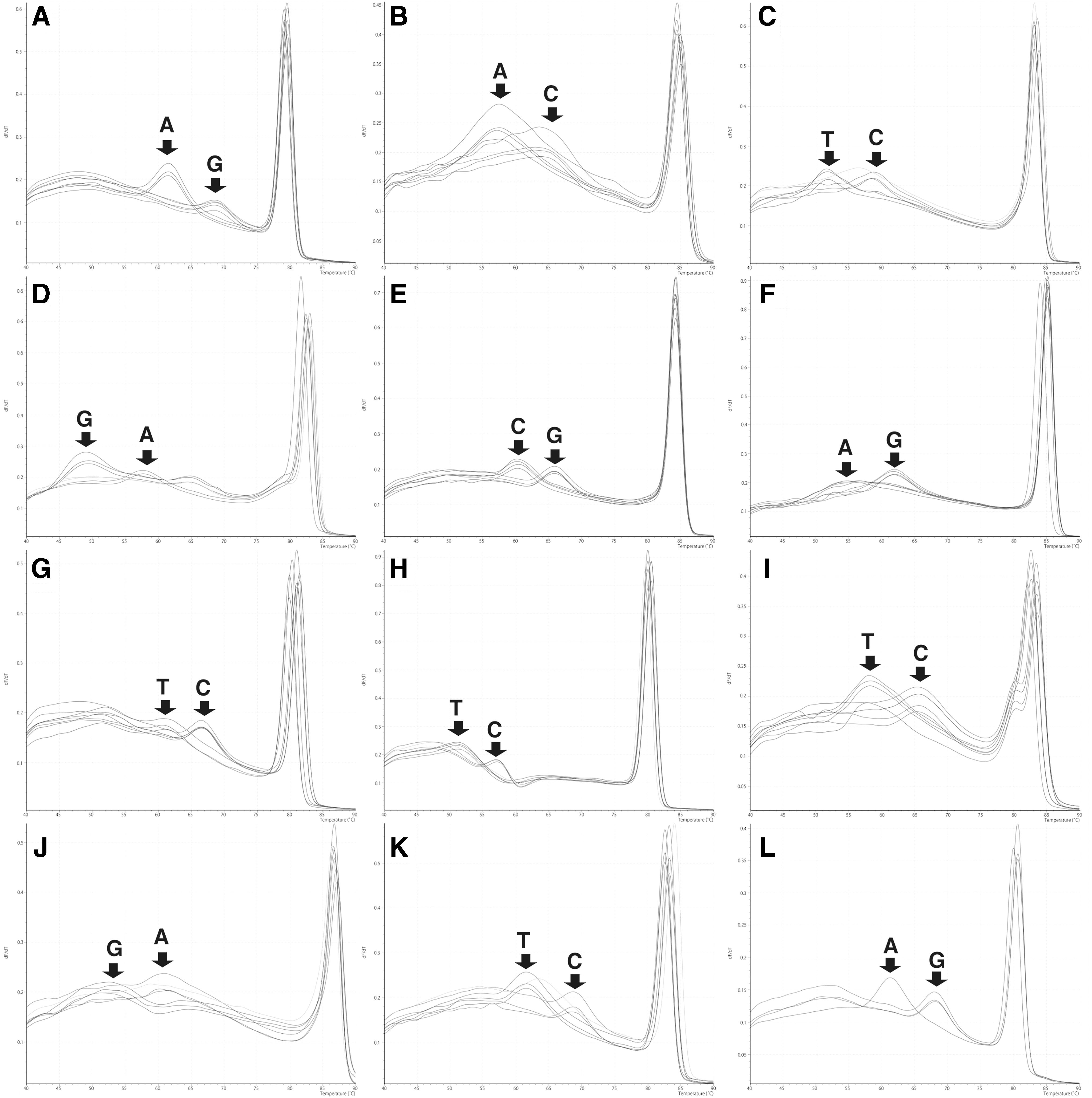
Representative melting peak plots obtained by stn-HRM analysis. Melting peak plots were developed by Tm calling analysis of melting curves obtained by stn-HRM. Arrows indicate positions of the peaks characteristically observed with each variant. The graphs indicate the results for SNP-like sites at
Validation analysis of stn-HRM
To assess the performance of the newly established stn-HRM procedure, we obtained the genomic DNA extract of Salmonella isolates independently collected in Japan, and analyzed them with the stn-HRM procedure. Based on the results of the HRM analysis, we determined ST for each Salmonella isolate (Table 4). On the other hand, serotypes of the organisms were determined by agglutination tests for the same isolates. Based on the stn-HRM results, we observed that 10 isolates belonged to “identifiable” STs (J1 to J10), 26 isolates belonged to “unidentifiable” STs (J11 to J36), and 9 isolates did not belong to any ST listed in Table 3 (J37 to J45). Among the 10 isolates belonging to “identifiable” STs (J1 to J10), serotypes estimated based on stn-HRM were consistent with the results obtained by agglutination test (Table 4, Category A), except for isolate J4 and J9 (Table 4, Category B). Serotype estimated based on stn-HRM for isolate J4 and J9 were serovars Dublin (ST16) and Paratyphi A (ST22), respectively, although serotype determined by agglutination test were serovar Berta and Singapore, respectively. In the alignment study, we could not obtain the stn gene sequence of serovar Berta and Singapore. In addition, only one and two sequences were obtained for ST16 and ST22, respectively (Table 3). These data suggest the need for accumulation of more information about stn sequence, especially for the STs with little stn sequence information. Among the 26 isolates determined as “unidentifiable” STs (J11 to J36), the serotypes determined by agglutination test for isolates J11 to J19, J22 to J24, J27 to J30, and J35 to J36 are listed in Table 3. While serotypes of these isolates were unable to be determined based on stn-HRM because these STs contain more than two serotypes, correspondences between ST and serotype were consistent with the results indicated in Table 3 (Table 4, Category C). Besides, the results of the agglutination test were the same for isolates J41, J42, and J43; they shared the stn-HRM result. We demonstrate that these data confirm the correlation between the results obtained by the stn-HRM and agglutination test.
Results of Validation Analysis with Salmonella Isolates That Were Independently Isolated in Japan
STs determined by stn-HRM analysis.
Results of validation analysis are categorized as below: (Category A) Serotype could be successfully determined by stn-HRM; (Category B) Serotype determined by stn-HRM was incorrect; (Category C) Serotype could not be determined by stn-HRM because corresponding STs include more than two serotypes in Table 3; (Category D) Serotype could not be determined by stn-HRM because serotype determined with agglutination test was not available in the alignment study; (Category E) Serotype could not be determined by stn-HRM because nucleotide usage patterns determined by stn-HRM did not belong to any ST listed in Table 3 (see Results for details).
Not typed. Nucleotide usage patterns determined by stn-HRM did not belong to any ST listed in Table 3.
Discussion
In the present study, we reveal a correlation between the stn gene sequence and the serotype of Salmonella species. In addition, based on the serotype-specific SNP-like typing, we identified the serotype-associated STs for some serovars. In the alignment study, we confirmed that 110 stn genes obtained from independent Salmonella isolates share the same ST in serovar Enteritidis (Table 3). While many stn gene sequences were obtained from frequently isolated serotypes, including serovar Enteritidis and Typhimurium, not enough information was obtained for some other serotypes. For example, only one sequence each was obtained for serovars Uganda (ST15), Wandsworth (ST17), Bareilly (ST21), Baildon (ST26), and Hvittingfoss (ST27). For serovars Javiana (ST6), Derby (ST7), Senftenberg (ST9 and 31), Infantis (ST20), and Paratyphi A (ST22), only two sequences were obtained, and for serovar Dublin (ST16), only three sequences were obtained (Table 3). Validation analysis suggested that more information is required to improve the accuracy of stn-HRM analysis especially for serotypes without enough stn sequences. In the alignment study, more than six sequences were obtained for nine STs (ST1, ST5, ST8, ST11, ST12, ST23, ST28, ST30, and ST32). These nine serotypes include frequently isolated serotypes and cover 64.3% of the cases reported to WHO/GFN Country Databank from 2001 to 2010 (Supplementary Table S1).
While stn-HRM could identify some frequently isolated serotypes such as serovar Enteritidis (ST1), Newport (ST11), Heidelberg (ST28), Hadar (ST8), and Agona (ST12), other frequent isolated serotypes such as serovar Typhimurium could not be identified. To improve the number of identifiable serotypes, we propose the application of additional analysis in combination with stn-HRM.
Case 1
Serotype combination can be discriminated by additional SNP-like sequence analysis.
Although serovars Virchow and Weltevreden share the same nucleotide usage pattern at selected 12 SNP-like sites (Table 3, ST10), they have a difference in nucleotide usage at position 571 (i.e., adenine is present at position 571 of serovar Virchow, and thymine is present at the corresponding position of serovar Weltevreden; Table 2). This difference suggests that in the case of a Salmonella isolate with the result of ST10, we could discriminate serovar Virchow from serovar Weltevreden based on additional analysis of nucleotide usage at position 571. In future studies, we intend to construct an HRM system for position 571.
Case 2
Serotype combination can be discriminated by O-antigen analysis.
The O-antigen analysis with an agglutination test is easy to conduct and requires only a few minutes to complete. In the case of ST3, serovars Montevideo, Panama, Minnesota, and Give share the same stn sequence not only in the 12 selected SNP-like sites but also in the whole stn gene (Table 3, ST3 and Supplementary Fig. S2, S5). These data indicate that these serotypes cannot be discriminated based on the stn gene sequence. On the other hand, these serotypes do not share the same O-antigen formula (i.e., O-antigen formula are 6,7,14 for serovar Montevideo, 1,9,12 for serovar Panama, 21 for serovar Minnesota, and 3 for serovar Give). This finding indicates that we can discriminate these serotypes by additional analysis of their O-antigen formula with agglutination test.
Case 3
Discrimination serovar Typhimurium from serovar 4,[5],12:i:-.
Based on the report by WHO, serovar Typhimurium is the second most frequently isolated serotype (
Using a combination of stn-HRM with simple additional analyses described above, some serotypes can be easily identified. If these simple additional analyses are employed as a supporting analysis for stn-HRM, the entire serotyping system can identify 83.7% of the cases reported to WHO/GFN Country Databank from 2001 to 2010.
Conclusions
Rapid serotyping procedures are considered a helpful tool to improve the credibility of the serotyping results by combining them with traditional methods (Shi et al., 2015). Most previously reported PCR-based procedures employed genes involved in O- or H-antigen synthesis as the target. In this study, we employed a unique target gene, stn, which is completely independent of the genes targeted in previous reports. Therefore, the results obtained by stn-HRM are independent of the results obtained from previously reported procedures. This feature gives usefulness to stn-HRM as a validation tool for serotyping. In addition, stn-HRM can ensure accurate identification of the bacterial species along with serotyping, because stn is a Salmonella-specific gene. It is expected that the combination of newly constructed stn-HRM and previously reported procedures could further improve the credibility of serotyping and species identification results.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported in part by Grants-in-Aid for Regional R&D Proposal-Based Program from Northern Advancement Center for Science & Technology of Hokkaido Japan, grant number: H21-T-3-7.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.