
Abstract
Listeria monocytogenes is a foodborne pathogen of global relevance that causes outbreaks and sporadic cases of listeriosis, acquired through the consumption of contaminated products, including milk or meat products and ready-to-eat meat products subjected to intensive handling. The objective of the present study was to classify L. monocytogenes isolated from various food-related sources in the Federal District of Brazil and surrounding areas to sequence internalin A (inlA) genes from these isolates and assess their adhesion and invasion capacity using Caco-2 cells. In addition, 15 were classified as group I, 3 as group II, and 7 classified as group IV. Premature stop codons (PMSCs) at the nucleotide position 976 (GAA→TAA) of the inlA gene were identified in 5 of the 25 isolates. Adhesion and invasion tests in Caco-2 cells showed that all the isolates were capable of adhesion and cellular invasion, with isolates containing PMSCs exhibiting on average higher invasion capacity than those without PMSCs (p = 0.041) and a median of adhesion very distinctive from those without stop codons. These results are the first report of PMSCs in the inlA gene of L. monocytogenes from the Federal District of Brazil and Brazil.
Introduction
L
L. monocytogenes is a motile, facultative, anaerobic, nonspore forming Gram-positive bacillus (Rocourt and Buchrieser, 2007). The genus Listeria includes 25 species, and L. monocytogenes is considered to be pathogenic toward both humans and animals (Chiara et al., 2015; Orsi and Wiedmann, 2016). Thirteen serotypes of this pathogen have been identified based on the presence of surface antigens, and serotypes 4b, 1/2b, and 1/2a are known to be responsible for more than 95% of listeriosis cases in humans (Graves et al., 1999).
Many bacteria are capable of invasion and survival in a wide variety of nonphagocytic cells (Cossart, 2002). In fact, the ability of bacteria to cross the intestinal, cerebral, and placental barriers determines the clinical aspects of infection. The presence of a class of proteins called internalins, mainly of types A (internalin A [InlA]) and B (InlB), in L. monocytogenes promotes the invasion of nonspecific tissue cells (Tsai et al., 2006; Orsi et al., 2007; Chen et al., 2009; Van Stelten et al., 2010). Studies have shown that L. monocytogenes can recognize different receptors, including transmembrane glycoproteins (such as E-cadherin), complement receptor (gC1qR), hepatocyte growth factor receptor (c-Met), and extracellular matrix components (such as proteoglycans). The InlA protein has been shown to specifically interact with E-cadherin on the surface of human intestinal epithelial cells (Gaillard et al., 1987; Ortega et al., 2017).
Listeria-induced disease conditions depend on the colonization of cells in the small intestine by these pathogens and their spread to other organs (Schubert and Heinz, 2003). Listeria cell invasion and virulence can be affected by the presence of nonsense mutations (leading to premature stop codons [PMSCs]) in the internalin genes that cause dysregulated expression of the internalin protein (Van Stelten et al., 2010; Chen et al., 2011; Kyoui et al., 2014). This dysregulation results in attenuated virulence of the pathogen and decreased invasion of epithelial cells in the small intestine (Nightingale et al., 2005; Van Stelten and Nightingale, 2008; Ward et al., 2010). Screening of L. monocytogenes isolated from human clinical cases, ready-to-eat (RTE) foods, and food processing environments has revealed mutations in inlA leading to PMSCs. Strains containing these PMSCs produce a truncated InlA protein and are generally less invasive than those without a PMSC (Nightingale et al., 2005; Cruz et al., 2014). Investigations conducted in the United States have detected L. monocytogenes strains with stop codons in 35–45% of food samples (Kyoui et al., 2014). Twenty-one types of PMSC mutations in the inlA gene have been described earlier (Gelbíčová et al., 2015). In Brazil, a few studies have investigated the prevalence of this foodborne pathogen (Lemes-Marques et al., 2007), specifically in the Federal District and Midwest regions.
Given the above findings and the fact that few studies on the molecular epidemiology and virulence factors in L. monocytogenes have been undertaken in Brazil, the present study aimed to characterize L. monocytogenes isolates from animal origin industrially processed food products in the Federal District of Brazil and surrounding areas by polymerase chain reaction (PCR)-based serotyping, phylogenetic analysis, and inlA sequencing. In addition, the invasion and/or cell adhesion efficiency of these isolates were also assessed.
Materials and Methods
Origin of the L. monocytogenes strains
Twenty-five L. monocytogenes isolates from different sources, including cuts of beef, hot dog sausages, refrigerated chicken carcasses, and a mechanically processed poultry sample (obtained from various commercial establishments), as well as isolates from poultry slaughterhouse drain swabs, were evaluated in this study. The strains were isolated during 2013–2018, and all experiments were carried out in the Laboratory of Food Microbiology (LAMAL) of the Faculty of Agronomy and Veterinary Medicine at the University of Brasília (FAV/UnB) (Table 1).
Sources of the Listeria monocytogenes Strains Isolated, Lineage, and Detection of Premature Stop Codons in the inlA Gene of Listeria monocytogenes at the Laboratory of Food Microbiology Faculty of Agronomy and Veterinary Medicine at the University of Brasília During 2013–2018
inlA, internalin A.
For isolation, 25 ± 1 g of each sample was weighed in sterile homogenization bags. Two hundred twenty-five milliliters of UVM broth (Acumedia®) was added, and the samples were incubated at 30°C ± 2°C for 24 h. Then, 0.1 mL of the sample was transferred from the UVM broth to a 10 mL aliquot of Fraser broth (Acumedia), which have been incubated at the same conditions (30°C ± 2°C) for 24 h. Finally, 0.1 mL of broth containing esculin hydrolysis was inoculated on Modified Oxford (MOX) agar medium (Difco®) supplemented with colistin and moxalactam and incubated at 30°C± 2°C for 24 h.
After incubation, colonies suspected of Listeria spp. based on their color and morphology were selected for identification. Multiplex PCR was used with primers for prfA (274pb; LIP1 and LPI2a) and prs (370pb; PRS1 and PRS2), as described by Kérouanton et al. (2010), to confirm the presence of L. monocytogenes. PCR was performed in a total volume of 50 μL, containing 0.4 μM of each primer (Invitrogen®), 200 μM dNTPs (Invitrogen), 0.2 mM MgCl2 (Invitrogen), and 2.5 U of Taq DNA polymerase (Invitrogen) in 1 × buffer (Invitrogen). PCR was carried out in a MyCycler thermal cycler system (Bio-Rad®) using the following conditions: denaturation for 3 min at 94°C, followed by 30 amplification cycles of annealing for 45 s at 53°C, elongation for 1 min 15 s at 72°C, denaturation for 40 s at 94°C, and a final extension step for 7 min at 72°C. PCR products were separated on a 2% agarose gel, stained with ethidium bromide (0.5 μg/mL), and visualized in an ImageQuant transilluminator (GE Healthcare Life Sciences).
Lineage of the isolated L. monocytogenes strains
The L. monocytogenes strains were serotyped using the PCR technique described by Doumith et al. (2004), in which amplification of specific fragments allows the classification of the strains into groups I, II, III, and IV. Group I comprises of serotypes 1/2a and 3a (amplification of Imo0737); group II comprises serotypes 1/2c and 3c (amplification of Imo0737 and Imo1118); group III comprises serotypes 1/2b, 3b, and 7 (amplification of ORF2819); and group IV comprises serotypes 4b, 4d, and 4e (amplification of ORF2819 and ORF2110).
Analysis of the inlA gene sequence
The method described by Poyart et al. (1996) was used to analyze the sequence of the 2400-bp inlA gene and detect stop codons. PCR was performed to amplify two regions of the inlA gene, nucleotides 192–1349 and 1472–2232 (Poyart et al., 1996). Primers O1 (5′-GAATGTAACAGACACG-3′—forward) and O2 (5′-ATCGCTAATAGTAGC-3′—reverse) were used to amplify a 1157-bp fragment, and primers O3 (5′-AGCCACTTAAGGCAAT-3′—forward) and O4 (5′-AGTTGATGTGTTAGA-3′—reverse) were used to amplify a 760-bp fragment. Each 50 μL reaction mixture contained 0.5 μM of the forward and reverse primers (Eurofins Genomics®), 200 μM dNTPs (Invitrogen), and 2.5 U of Platinum™ Taq DNA polymerase high fidelity (Invitrogen) in a 1 × enzyme buffer solution. The cycling conditions were 5 min at 95°C, followed by 40 cycles of 1 min at 55°C, 90 s at 72°C, and 1 min at 94°C, and a final extension step for 7 min at 72°C. The PCR products were separated on a 2% agarose gel, stained with ethidium bromide (0.5 μg/mL), and visualized on an ImageQuant transilluminator. The amplified inlA gene fragments were purified using the PureLink Kit (Invitrogen), quantified with the High DNA Mass Ladder (Invitrogen), and sequenced with an ABI 3500 (Applied Biosystems®) sequencing setup. The obtained nucleotide sequences were compared to those available in the National Center for Biotechnology Information (NCBI) database using the Basic Local Alignment Search Tool (BLAST) program, using inlA L. monocytogenes EGD-e (NCBI: NC_003210.1) as reference. A phylogenetic tree was generated by the maximum likelihood method based on an alignment of the partial nucleotide sequences of the inlA gene from the 25 L. monocytogenes isolates.
Caco2 cell culture
Caco2 (human colon adenocarcinoma) epithelial cells were kindly provided by Professor Elaine Cristina Pereira De Martinis and are from the collection housed in the Faculty of Pharmaceutical Sciences of the University of São Paulo (USP). The cells were cultured as described by Winkelströter and De Martinis (2015). Briefly, the cells were cultured in Dulbecco's modified Eagle's medium (DMEM) (Sigma-Aldrich®) high glucose, supplemented with 10% fetal bovine serum (Gibco®) and 100 U/mL each of penicillin and streptomycin (Sigma-Aldrich), and maintained at 37°C in a 5% CO2 incubator (Q316C; Quimis®) for 24 h until semiconfluent. Then, the cells were trypsinized (Trypsin; Sigma-Aldrich) and suspended in DMEM at a density of 2.5 × 105 cells/mL. A 1-mL aliquot of the cell suspension was transferred to each well of a 24-well polypropylene plate (Kasvi®) and incubated at 37°C in a 5% CO2 incubator (Quimis) for 24 h. The cells that were found adhered to the plates were used for L. monocytogenes adhesion and invasion assays.
Three-dimensional cell culture
To produce spheroids containing eukaryotic (Caco-2) three-dimensional (3D) cells, Cytodex® (>175 μm spherical particle size in 0.9% NaCl solution; Sigma) was used as microcarrier beads. A commercial bioreactor (Synthecon) was used according to the work of Radtke and Herbst-Kralovetz (2012). The bioreactor was incubated with rotation adjusted for 20 rpm, and the culture was incubated for further 96 h at 37°C with 5% CO2 in atmosphere. Following that, every other day, ca. 80% of the culture medium DMEM was replaced and incubation continued for up to 21 days. Cells were harvested at selected times, using a 1000 μL cut pipette tip (to prevent shear and protect the 3D structure).
Adhesion assays
The cell adhesion assays were performed as described by Moroni et al. (2006), with modifications. Briefly, L. monocytogenes strains were inoculated in brain–heart infusion broth (BHI; Difco) and incubated for 24 h at 37°C in a bacteriological incubator (Quimis). Then, serial dilutions of the bacterial suspension were prepared in sterile 1 × phosphate buffered saline (PBS) and plated on BHI (Difco) containing 1.5% bacteriological agar (Acumedia). Subsequently, the colonies were counted to determine the concentration (expressed in colony-forming unit [CFU]/mL). Caco-2 cells adherent to polypropylene plates (Kasvi) were washed thrice with 1 × PBS and infected with 100 μL of a bacterial suspension with a number density of 1 × 107 CFU/mL. Then, 300 μL of DMEM supplemented with 10% fetal bovine serum without antibiotics was added to each well. The plates were incubated in a 5% CO2 incubator for 2 h at 37°C. After incubation, the plates were again washed thrice with 1 × PBS to remove nonadherent bacteria. The cells were then treated with a lysis solution, consisting of 0.1% Triton X-100 (Sigma-Aldrich) in 1 × PBS, for 10 min at 37°C. The total viable bacteria in the cell lysates were determined by quantification of serial dilutions on BHI plates containing 1.5% bacteriological agar (Acumedia), and the results were expressed in CFU/mL. All experiments were performed in triplicate. The percentage of L. monocytogenes adherent to Caco2 cells was calculated multiplying the number of adherent cells by 100 and dividing by the number of cells added at the beginning of the experiment.
Cell invasion assay
The cell invasion assays were performed as described by Moroni et al. (2006), with modifications. L. monocytogenes strains were inoculated in BHI (Difco) and subsequently incubated for 24 h at 37°C in a bacteriological incubator (Quimis). Then, serial dilutions of this suspension were prepared in sterile 1 × PBS, which were replated on BHI (Difco) containing 1.5% of bacteriological agar (Acumedia), and the results were expressed in CFU/mL. Caco-2 cells adherent to polypropylene plates (Kasvi) were washed thrice with 1 × PBS and infected with 100 μL of a bacterial suspension containing 1 × 107 CFU/mL. Then, 300 μL of DMEM (Sigma-Aldrich) supplemented with 10% fetal bovine serum (Gibco) without antibiotics was added to each well. The plates were incubated in a 5% CO2 incubator (Quimis) for 2 h at 37°C. After incubation, the plates were again washed thrice with 1 × PBS to remove nonadherent bacterial cells. Then, 250 μL of sterile water containing 1 mg/mL gentamycin (Sigma-Aldrich) was added to each well and incubated for 1 h at 37°C in a 5% CO2 incubator (Quimis). After incubation, the plates were washed thrice with sterile 1 × PBS, mixed with 250 μL of cell lysis buffer consisting of 0.1% Triton X-100 (Sigma-Aldrich) in 1 × PBS, and incubated for 10 min at 37°C. The total number of viable bacterial cells in the cell lysate was determined by counting the serial dilutions seeded on BHI plates (Difco plus 1.5% bacteriological agar; Acumedia), and the results were expressed in CFU/mL. All experiments were performed in triplicate. The percentage of L. monocytogenes cells engulfed by Caco2 cells was calculated through the number of internalized bacterial cells recovered by multiplying by 100 and dividing by the number of cells adhered.
Statistical analyses
Statistical analyses were performed with the STATA 12® program (StataCorp, 2011). Descriptive statistics of the mean rates of adhesion and invasion by the L. monocytogenes isolates with and without stop codons in inlA was calculated. To compare isolates with and without PMSCs, these variables were initially tested for normality with the Shapiro–Wilk test. For the nonparametric data, the Kruskal–Wallis test was used. A statistically significant difference was defined as p < 0.05.
Results and Discussion
Strain lineage of L. monocytogenes isolates
Of the 25 L. monocytogenes isolates, 15 were classified as group I (15/25—60%), 3 as group II (3/25—12%), and 7 classified as group IV (7/25—28%). The sources of the samples from where the bacteria were isolated, along with their respective classification, are shown in Table 1.
Thirteen L. monocytogenes serotypes have been earlier reported in literature (Norton et al., 2001; Gray et al., 2004; Fugett et al., 2007), and the lineage reported in the present study has been detected in more than 90% of listeriosis cases and outbreaks (Jawetz et al., 1998; Cabrita et al., 2004). According to Lianou et al. (2006), serotypes 1/2a and 1/2b are more frequently involved in sporadic cases of listeriosis, whereas serotype 4b is mostly associated with outbreaks. Serotype 1/2c, although less frequently reported, is commonly associated with foods of animal origin (Lambertz et al., 2013; Vallim et al., 2015; Palma et al., 2016) and environments associated with these foods (Nalério et al., 2009; Lambertz et al., 2013). Regarding the genetic lineages described by Norton et al. (2001), Gray et al. (2004), and Fugett et al. (2007), serotype 1/2a and 4b are classified in lineage I, which is often reported in cases of human listeriosis (Hofer et al., 2006; Lemes-Marques et al., 2007).
The lineage detected in this study was also reported by Kramarenko et al. (2013) in different foods of animal origin in Estonia and by Wu et al. (2015) in RTE foods of animal origin in China. The same serotypes were described by Doumith et al. (2004) in India and by Braga et al. (2017) in frozen food products available in retail markets in Montevideo, Uruguay.
Partial analysis of inlA gene sequences
The results of the inlA gene sequence analysis are shown in Table 1. The partial sequencing of the inlA gene revealed the presence of PMSCs at position 976 (GAA→TAA) in 5 of the 25 (20%) L. monocytogenes isolates, including 3 isolates from bovine meat samples, 1 identified in a chicken carcass isolate, and 1 isolated from mechanically processed poultry.
There were five L. monocytogenes isolates detected in this study with PMSCs at the nucleotide position 976 of the inlA gene. These results are identical to the findings of Gelbíčová et al. (2015), who detected a PMSC in the same position in a L. monocytogenes serotype 1/2c isolate from a clinical sample in the Czech Republic. Analysis of inlA sequences from the different L. monocytogenes isolates showed that the presence of PMSCs in positions close to that identified in the present study does not interfere with the invasion ability (Nightingale et al., 2005; Chen et al., 2011). Mutations that lead to PMSCs in the inlA gene have been reported in isolates from food, but rarely in clinical isolates (Van Stelten et al., 2010). When clinical isolates were analyzed, 96% were found to contain the intact inlA gene, compared to 65% of food isolates (Jacquet et al., 2004). The presence of stop codons in regions close to that identified in our study (position 976) has been previously described in human clinical isolates and food isolates in other countries (Nightingale et al., 2005; Felício et al., 2007; Ragon et al., 2008; Van Stelten et al., 2010; Cantinelli et al., 2013; Kovacevic et al., 2013). However, there are no reports on the presence of PMSCs in the inlA gene in Brazilian samples. Therefore, the present study is the first to evaluate this in the country and the region. The results obtained in this study indicate that these isolates represent a potential pathogenic risk in relation to the observed position of the identified PMSCs.
Phylogenetic analysis of the inlA sequences and serotypes identified in this study using Geneious version 7.1.5 (Fig. 1) allowed grouping of isolates with PMSCs. This analysis shows similarities between isolates 3, 26, 27, and 36, indicating a possible shared origin. The isolate 10, which also has a PMSC, was found belonging to a close but different group that contained isolates without PMSC, suggesting that this characteristic is not monophyletic, despite the small number of isolates and few discriminatory data analyzed.
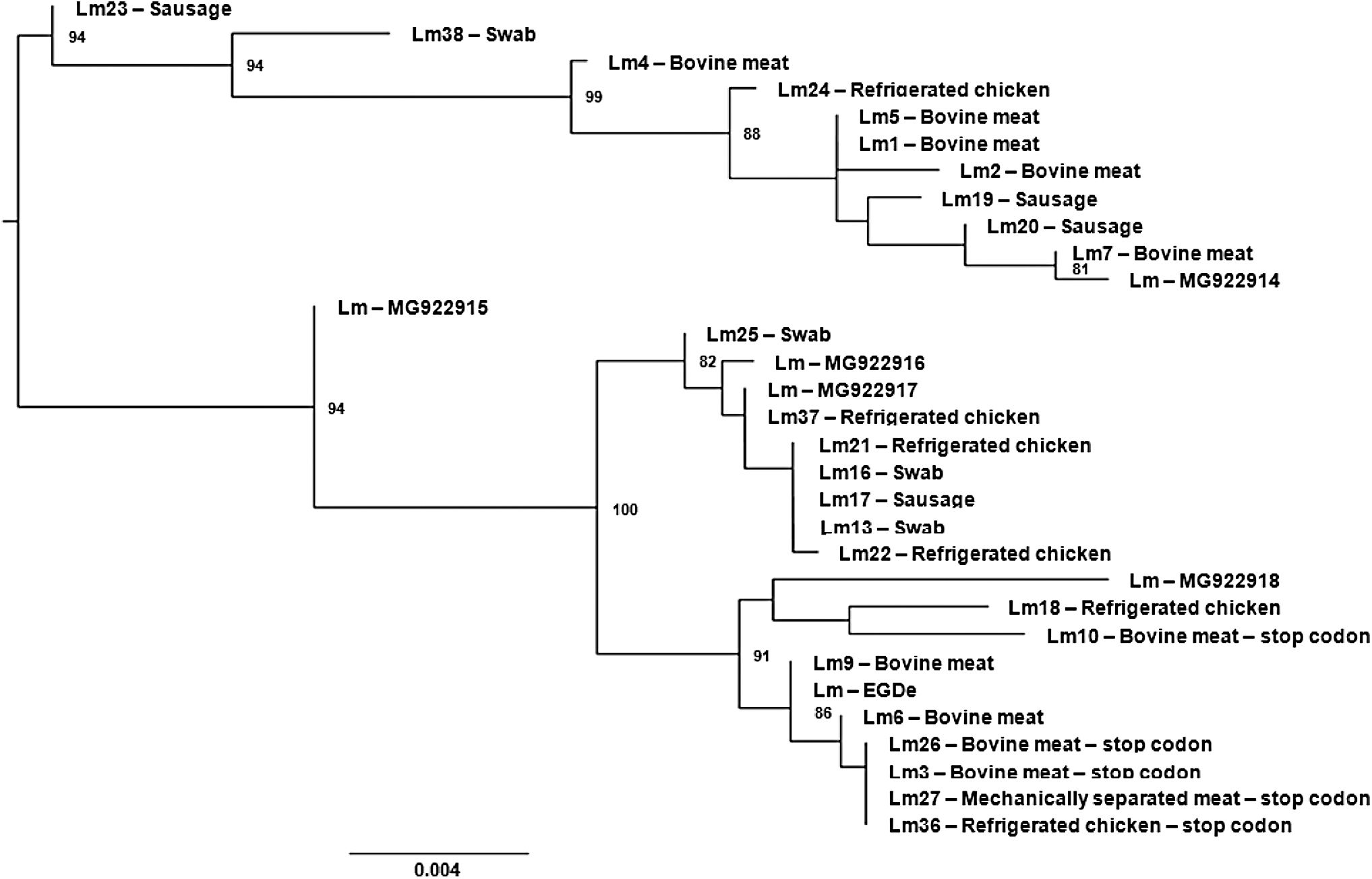
Phylogenetic analysis of the inlA genes from Listeria monocytogenes isolates. The phylogenetic tree was generated by the maximum likelihood method based on an alignment of the partial nucleotide sequences of the inlA gene from the 25 L. monocytogenes isolates obtained and from other 6 obtained from databanks. Branches with <70% bootstrap support were collapsed. inlA, internalin A.
Adhesion and invasion of Caco-2 cells by L. monocytogenes isolates
The results of the adhesion and invasion assays carried out with the 25 L. monocytogenes isolates, both with and without PMSCs, in Caco-2 cells are detailed in Table 2 and Figure 2.
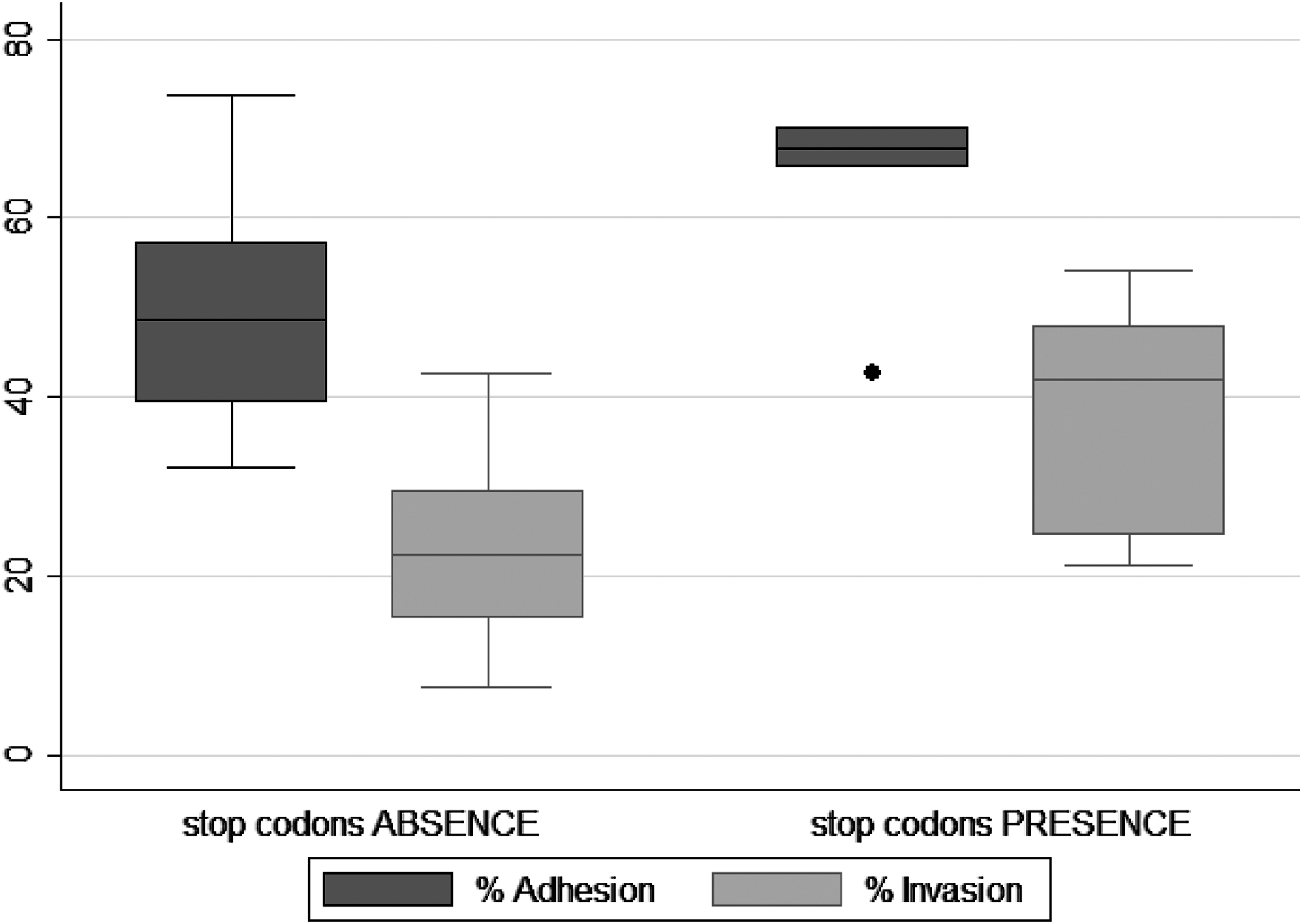
Adhesion and invasion capability of Listeria monocytogenes isolates (with and without premature stop codons) of diverse origins toward Caco-2 cells.
Adhesion and Invasion Capability of Listeria monocytogenes Isolates (With and Without Premature Stop Codons) of Diverse Origins Toward Caco-2 Cells
Adhesion and invasion assay were performed on 3D Caco-2 cells. For the tests in 3D model, some samples were chosen in which two did not have stop codons and three samples did. The results are shown in Table 3.
Adhesion and Invasion Capability of Listeria monocytogenes Isolates (With and Without Premature Stop Codons) of Diverse Origins Toward Three-Dimensional Caco-2 Cells
All 25 isolates were able to adhere to and invade Caco-2 cells, with percent adhesion values in the range 32.2–73.7% and invasion values of 7.6–54.15%. In particular, the five isolates that contained PMSCs had percent adhesion and invasion mean rates of 63.26% (standard deviation [SD] 11.6%; median 67.7%) and 37.95% (SD 14.53%; median 42%), respectively. Isolates without PMSCs had percent adhesion and invasion mean rates of 49.7% (SD 11.9%; median 48.7%) and 23.65% (SD 10.64%; median 22.4%), respectively.
The listerias of this work had a high rate of adhesion and invasion when tests were carried out in a monolayer system. When the tests were performed with 3D cells, there was less invasion. This is expected, since 3D cells mimic what occurs in vivo (Peterson and Artis, 2014; Drummond et al., 2016).
The L. monocytogenes isolates that contained PMSCs also exhibited the ability to adhere to and invade Caco-2 cells. Therefore, the presence of a stop codon at nucleotide position 976 does not appear to hinder the adhesion and invasion ability of the isolates, which is corroborated by results for isolates with PMSCs previously reported by Jacquet et al. (2004), who observed the same findings for a C → T transition at position 565, and Van Stelten and Nightingale (2008), who observed similar results for a C → T transition at position 229 and a G → T mutation at position 508. Van Stelten et al. (2010) also detected a T → A mutation at nucleotide position 758, which was also found to not prevent the invasion and adhesion of L. monocytogenes in Caco-2 cells, but demonstrated attenuated virulence. The vast majority of clinical isolates have complete inlA, contrary to what happens in food isolates; a growing number of evidences suggest no direct correlation between either inlA sequence or InlA integrity and the capability to invade cells or to cause infection (Su et al., 2019).
The statistical analysis showed variability in the adhesion and invasion capacity of the isolates in Caco-2 cells. The results of the adhesion and invasion assays were subjected to the Kruskal–Wallis analysis to assess the statistical differences between means of groups with and without stop codons. Analysis of the adhesion rates did not show, with 95% confidence (p = 0.057), difference between the two groups (with and without PMSCs). However, there was a relevant difference between the medians of the two types of isolates, demonstrating a tendency toward higher adhesion capacity in those samples with stop codon. Analysis of the invasion rate confirmed, with 95% confidence (p = 0.041), that the isolates containing a PMSC had on average a higher rate of invasion than those without a PMSC. Nevertheless, both groups of isolates were able to adhere to and invade Caco-2 cells.
These results did not allow us to unambiguously conclude that L. monocytogenes isolates with a PMSC at position 976 have greater adhesion or invasion capacity compared to strains without this mutation, since this characteristic was only found in five of the analyzed isolates, but allow us to discuss this significant finding at these samples. Winkelströter and De Martinis (2013), da Silva et al. (2017), and Xie et al. (2018) have reported that other mechanisms, in addition to that associated with the inlA protein, could be involved in bacterial adhesion and invasion. The increase in invasion in mutant InlA strains can be explained by the presence of other structures that also favor invasion. Su et al. (2019) state that internalin profiles were not associated with L. monocytogenes ability to invade human Caco-2 cells. Possibly there is the action of a set of proteins involved. There are several internalins identified in the genome, being encoded by different genes, as an example inlD which is related to invasion activity (Seveau et al., 2007).
Therefore, the results of this research corroborate with the literature that InlA has an important, although not exclusive, role in determining the virulence of L. monocytogenes strains (Van Stelten et al., 2016; Dellafiora et al., 2020). Among other factors related to invasion is listeriolysin O (pore-forming toxin), which is known as an important cooperative determinant to increase the efficiency of host cell invasion (Phelps et al., 2018). Listeriolysin O, in addition to interrupting the phagosome to allow bacterial proliferation in the cytosol, also perforates the plasma membrane, facilitating the internalization of L. monocytogenes in epithelial cells. Thus, listeriolysin O also plays a key role in bacterial invasion of epithelial cells. InlA and LLO are the most critical invasion factors, although they act differently and depending on the type of cell (Phelps et al., 2018). These same authors showed that the primary function of the InlA-E-cadherin interaction is to anchor the bacterium on the host's surface, but this interaction can also control the efficiency of bacterial internalization in some, but not all cell lines. Thus, several factors could interfere in the adhesion and invasion of Listeria.
The adhesion process of L. monocytogenes facilitates cell invasion by activating a signaling cascade in the host cell; this stage involves several bacterial surface adhesion factors (Camejo et al., 2011). For each adherence stage to occur, it depends on several bacterial factors that act synchronously and are being controlled by a specific set of regulatory genes (Camejo et al., 2011). The orchestrated interaction involves the participation of virulence factors specifically regulated at the transcriptional, post-transcriptional, and post-translational (Camejo et al., 2011). The main virulence factors present in adherence are Lap, Ami, DltA, FbpA, InlJ, CtaP, LapB, ActA, RecA, and InlF. The virulence factors present in the invasion are InlA, InlB, Vip, Auto, P60, Lgt, GTCA, LpeA, MprF, LLO, Flagella, and ActA (Camejo et al., 2011). Thus, in this way, several virulence factors may be involved at the same stage of the cycle, with some operating at the same stage and others having a more restricted action.
Dellafiora et al. (2020) argue that despite the prevalence of full-length InlA sequences in clinical isolates, a growing body of evidence suggests that there is no direct correlation between the inlA sequence or InlA integrity and the ability to invade cells or cause infection. Experimental data pointed out that a strong InlA-Ecad interaction may not be essential to determine the cell invasion capacity of L. monocytogenes (Dellafiora et al., 2020).
Analyses of missense mutations, using computational models, demonstrate possible effects of its mutations on the capability to interact with Ecad and that a strong interaction between InlA and Ecad is important, but perhaps not essential in the capacity for cell invasion (Dellafiora et al., 2020). Dellafiora et al. (2020) call attention to missense mutations, even though they are largely ignored; they need to be investigated in relation to their effects on cell invasion processes. The strength of the interaction may not be directly related to the capacity for cellular invasion, reflecting the nonexclusive role of inlA in determining the virulence of L. monocytogenes (Dellafiora et al., 2020).
InlA binding to E-cadherin is sufficient for Listeria invasion; however, modulation of endocytosis by InlB accelerates this process, and although dispensable for attachment, it synergistically promotes invasion of multicellular junctions (MCJs) through activation of c-Met kinase signaling (Pentecost, 2010). In addition, caveolin endocytosis is essential for the internalization of listeria, as the bacterial transfer of cells in the cell is severely impaired when the cells are depleted in caveolin-1 (Dhanda et al., 2020). The mechanisms of invasion of listeria are not yet fully understood; it is known that M host cells and InlA are involved; however, L. monocytogenes can cross the intestinal barrier in its absence (Drolia and Bhunia, 2019). The interaction of the Listeria adhesion protein (LAP) with the host cell receptor (heat shock protein 60) breaks the epithelial barrier, promoting bacterial translocation (Drolia et al., 2018; Drolia and Bhunia, 2019).
Studies with a greater number of L. monocytogenes isolates need to be performed to determine whether a PMSC has any effect on the invasion of Caco-2 cells and to evaluate the possibility of other mechanisms of bacterial adhesion and invasion. Nevertheless, cell adhesion and invasion ability of the L. monocytogenes isolates estimated in this study allowed us to suggest that they could represent a potential public health risk. In addition, our study represents the first report of PMSCs present in the inlA gene in Brazil in general and the Federal District of Brazil in particular.
Conclusions
L. monocytogenes isolates with lineage I, II, and IV were detected in different sources, such as commercial beef cuts, hot dog sausages, refrigerated poultry carcasses, mechanically processed poultry, and swabs from floor drains of a poultry slaughterhouse. Sequencing of the inlA gene of these isolates revealed the presence of a PMSC in five isolates. The percent mean of invasion capacity of isolates with a PMSC in inlA was significantly higher compared with isolates without the mutation. These results highlight the public health risk associated with these L. monocytogenes isolates of diverse origin and may serve as a basis for future studies evaluating other factors involved in the virulence of this bacterial pathogen.
Footnotes
Acknowledgments
The authors thank the teams in the Laboratories of Food Microbiology, Veterinary Medical Microbiology, and Gene Therapy Technologies at the University of Brasília, as well as the Pharmaceutical Sciences Laboratory at the USP, Ribeirão Preto, Brazil.
Disclosure Statement
No conflicts of interest to be declared.
Funding Information
This work was supported by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), Brazil (Finance Code 001) and the Research Dean and Postgraduate program of the University of Brasília. Financing agencies played a role in granting scholarships and purchasing chemicals.