
Abstract
Shiga-toxin producing Escherichia coli (STEC) are zoonotic foodborne pathogens that are capable of causing serious human illness. Ovine ruminants are recognized as an important source of STEC and a notable contributor to contamination within the food industry. This review examined the prevalence of STEC in the ovine food production chain from farm-to-fork, reporting carriage in sheep herds, during abattoir processing, and in raw and ready-to-eat meats and meat products. Factors affecting the prevalence of STEC, including seasonality and animal age, were also examined. A relative prevalence can be obtained by calculating the mean prevalence observed over multiple surveys, weighted by sample number. A relative mean prevalence was obtained for STEC O157 and all STEC serogroups at multiple points along the ovine production chain by using suitable published surveys. A relative mean prevalence (and range) for STEC O157 was calculated: for feces 4.4% (0.2–28.1%), fleece 7.6% (0.8–12.8%), carcass 2.1% (0.2–9.8%), and raw ovine meat 1.9% (0.2–6.3%). For all STEC independent of serotype, a relative mean prevalence was calculated: for feces 33.3% (0.9–90.0%), carcass 58.7% (2.0–81.6%), and raw ovine meat 15.4% (2.7–35.5%). The prevalence of STEC in ovine fleece was reported in only one earlier survey, which recorded a prevalence of 86.2%. Animal age was reported to affect shedding in many surveys, with younger animals typically reported as having a higher prevalence of the pathogen. The prevalence of STEC decreases significantly along the ovine production chain after the application of postharvest interventions. Ovine products pose a small risk of potential STEC contamination to the food supply chain.
Introduction
Shiga-
This review is focused on the carriage and prevalence of STEC (an E. coli that carries the stx gene) in ovine ruminants, indiscriminate of age, breed, and sex. The prevalence of STEC O157 and non-O157 is discussed. STEC O157 when referenced throughout the text is with respect to serogroup O157 and is inclusive of all flagellar antigens, including E. coli O157:H7. This review aims at examining the prevalence rates of this pathogen from farm-to-fork, reporting carriage in domestic sheep herds, during abattoir processing and in raw and ready-to-eat meats and meat products. The impact of decontamination treatments and other intervention processes was not addressed in this study.
Research papers and reports containing relevant data included in this review are discussed in the text. Some studies have also been organized into tables. Data presented in the tables have been limited to studies that include a minimum of 95 samples, and that clearly and unambiguously report the identification of Shiga-toxin positive samples, with no preference for the detection techniques employed or the sample type. Studies published before the year 2001 have not been included. Studies that involve mixed-species samples, such as sheep and goat, have not been included where possible. Smaller studies of relevance are also discussed in the text. Studies that did not meet the criteria for inclusion are listed in Supplementary Table S1.
An indication of the relative prevalence, and the observed range of prevalence rates, of STEC O157 and non-O157 STEC at different stages of production was calculated by using the surveys presented in each of the tables. A relative mean was obtained by calculating the mean prevalence observed over the selected surveys, weighted by the sample number. Hence, the relative mean can be expressed as: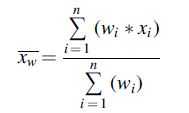
where
Shiga-toxin producing E. coli
STEC are significant enteric pathogens associated with disease, both sporadically and in outbreaks, resulting in the manifestation of serious diarrheal symptoms and the potential onset of hemorrhagic colitis or hemolytic uremic syndrome (Nataro and Kaper, 1998; Petras, 2011; Trachtman et al., 2012). STEC O157 is the most frequently isolated pathogenic STEC serogroup globally, accounting for 36.6% and 41% of STEC infections in the United States and the European Union, respectively, in 2015 (CDC, 2017; EFSA and ECDC, 2018).
Four additional serogroups (O26, O103, O111, O145) are recognized as important adulterants and are collectively referred to, along with E. coli O157:H7, as the “top five” STEC (EFSA BIOHAZ Panel et al., 2020). However, a recent EFSA Scientific opinion (2020) concluded that based on the current available evidence, serogroup cannot be used as a predictor of clinical outcome and that the presence of the intimin gene (eae) is not essential for severe illness and thus that all STEC strains are pathogenic in humans, capable of causing at least diarrhea, and that all STEC subtypes may be associated with severe illness. Ovine ruminants harbor a diverse range of non-O157 STEC, many of which share virulence features with strains recovered from disease-causing outbreaks (Blanco et al., 2003; Aktan et al., 2004; Djordjevic et al., 2004; Cookson et al., 2006; Kalchayanand et al., 2007).
STEC pathogenic factors
STEC are characterized by the production of one or more Shiga toxin(s) (denoted as Stx) that target the globotriaosylceramide (Gb3) receptor found along human renal endothelial cells and monocytes (O'Brien et al., 1982; Phillips et al., 2000; Obrig, 2010; Schüller, 2011; Piérard et al., 2012). Ruminants lack this receptor, possessing a Gb4 receptor variant instead, and can therefore harbor the pathogen asymptomatically for prolonged periods of time (Gyles, 2007). There are two major families of the Stx toxin (Stx1 and Stx2), which share an estimated 55% amino acid sequence homology (Kaper et al., 2004; Caprioli et al., 2005). They are encoded on mobile lysogenic prophages commonly referred to as stx phages (Krüger and Lucchesi, 2015). Strains may express one or both families of the toxins, and several variants of each are known (Melton-Celsa, 2014; EFSA BIOHAZ Panel et al., 2020). Ruminants may preferentially harbor different stx variants (Mora et al., 2012; Bai et al., 2016; Shridhar et al., 2017). The subtypes stx2d and stx1c- are strongly associated with sheep, whereas recent reports have highlighted the increasing prevalence of subtype stx2O118 in ovine ruminants (Koch et al., 2001; Ramachandran et al., 2001; Brett et al., 2003; Alonso et al., 2016).
The ability to adhere to and successfully colonize the lower gastro-intestinal tract of the host is a critical factor in the successful population and persistence of STEC in ovine ruminants (La Ragione et al., 2009). Invading STEC typically induce the formation of attaching and effacing (A/E) lesions to allow intimate adherence between the bacterium and the host intestinal epithelial cells (Cepeda-Molero et al., 2017). The factors responsible for the formation of A/E lesions are encoded on a pathogenicity island (PAI) called the locus of enterocyte effacement (LEE) (Elliott et al., 1998). The major elements encoded include a Type III secretion system (T3SS), the intimin protein, its receptor Tir, and the Esp proteins (Nataro and Kaper, 1998; Garmendia et al., 2005).
STEC may also harbor other important PAIs, including O-island (OI) 122, OI-43/48, and the high pathogenicity island. These encode many important non-LEE encoded (nle) effector genes as well as alternative virulence factors, including the toxin Sen (sen), adhesion factors (efa1, efa2, iha, aid-A1), and antimicrobial resistance genes (ureC, terC) (Schmidt and Hensel, 2004; Bugarel et al., 2010; Ju et al., 2013; Franz et al., 2015; Karama et al., 2019).
STEC and ovine ruminants
Ovine ruminants are recognized as notable contributors to STEC contamination to the food supply chain. Intrinsic factors such as animal age and sex may make an animal more susceptible to STEC infection, whereas extrinsic factors associated with the farm environment are the leading contributors to the animal becoming infected. It has been demonstrated that STEC can survive for extended periods in animal feces and in the farm environment. STEC may persist in drinking troughs, animal feed, soil, or effluent (Soon et al., 2011). The continued exposure to the pathogen results in the indirect infection or re-infection of ovine ruminants from the environment, whereas direct transmission between animals can also occur via the fecal–oral route (Persad and LeJeune, 2014).
Sheep farming is a global practice. Farmers in the northern and southern hemisphere, while still producing ovine meat and wool all year, follow seasonal production times. Both regions observe their busiest lambing period in the spring with seasonal demand peaking toward Easter and Ramadan, whereas the southern hemisphere also observes a second seasonal peak in the early Autumn (MLA Corporate, 2019). Sheep production in the southern hemisphere tends to be more diverse than what is observed in European production systems, with many famers keeping sheep to produce wool and mutton meat as well as lamb production. Slaughtered animals tend to be older in these regions, with lower weaning rates as a result, when compared with other sheep-producing countries in Europe (Behrendt and Weeks, 2019). Lamb producers in countries such as Spain, Greece, or France may have multiple lambing events per annum, and typically slaughter lambs at a much younger age (<3 months of age) than other regions, with consumers preferring younger, preweaned lamb meat (Campo et al., 2016).
A wide variety of ovine sources have been associated with STEC outbreaks. Many of the reported outbreaks are as a result of poor hygiene after interaction with animals at specified events or petting farms or ingestion of fecally contaminated soil (Licence et al., 2001; Bekal et al., 2014; Gobin et al., 2018). Other outbreaks are associated with consumption of contaminated raw meat products or undercooked meat/ready-to-eat ovine produce (Espié et al., 2006; Sekse et al., 2009; Wahl et al., 2011). The diverse range of infection sources highlights the disease-causing potential of ovine ruminants and the importance of obeying good hygiene standards after animal contact and when preparing food (Rowell et al., 2016).
STEC colonization in ovine hosts
STEC have been shown to demonstrate effective colonization at the terminal end of the rectum, termed the recto-anal junction (RAJ), in cattle (Naylor et al., 2003; Low et al., 2005). However, colonization in ovine hosts has been shown to be less specific compared with bovine animals (La Ragione et al., 2009; Vande Walle et al., 2011). Colonization dynamics between ovine hosts and the bacterium are notoriously difficult to investigate due to the absence of a stable ovine colonization model in artificially inoculated animals (Cookson et al., 2002; Sheng et al., 2004; Best et al., 2009).
STEC colonization is dependent on several factors. The intimin protein is essential for A/E lesion formation and subsequent strain colonization in sheep (Cornick et al., 2000). A/E lesions primarily target intestinal epithelial cells (Cepeda-Molero et al., 2017). Colonizing STEC may also secrete other important adherence factors, including a selection of Esp and Nle proteins (La Ragione et al., 2009; Ojo et al., 2010). The secretion of such proteins is suspected to illicit an immune response in the host, which contributes to the removal of the pathogen from the animal (Sanderson et al., 1999; Yekta et al., 2011). Interestingly, an immune response is only seen after natural or artificial oral infection with STEC, with some suggestions that the bacterium must pass through the small intestine to elicit an antibody reaction (Vande Walle et al., 2011). Colonization in ovine ruminants has also been shown to be positively affected if the invading STEC strain possesses either of the lpf adhesion fimbrieae (Farfan and Torres, 2012).
Tissue tropism has been shown to be an important factor in E. coli O157:H7 pathogenesis and successful colonization in human and bovine studies (Naylor et al., 2003; Sheng et al., 2004). In contrast, STEC colonization studies in small ruminants have identified non-preferential selection of different tissues during colonization. La Ragione et al. (2009) demonstrated that E. coli O157:H7 colonization was specific to the distal intestine in young lambs. Interestingly, this study also demonstrated that E. coli O157:H7 had no specific tropism for lymphoid tissue, something that is commonly observed in bovine hosts (Naylor et al., 2003; Bonardi et al., 2007). Other studies have shown that shedding of E. coli O157:H7 for ≥14 days (persistent shedding) is correlated with colonization throughout the entire gastrointestinal (GI) tract of 6-week-old lambs (Cookson et al., 2002; Woodward et al., 2003). Similar colonization patterns have been observed for STEC O26. A nontoxigenic E. coli O26:K60 strain was orally inoculated in young lambs and found to colonize the whole GI tract during high excretion. However, after day 38, A/E lesions could only be recovered from the distal small intestine (Aktan et al., 2007).
To date, no clear evidence has been presented, which would indicate that STEC has a preferred site of colonization in ovine hosts. Instead, it is capable of colonization along the entire GI tract, including the RAJ, but to a lesser degree than that observed in cattle (La Ragione et al., 2009; McPherson et al., 2015).
Super-shedding ruminants
Ovine ruminants are important contributors to the dissemination of STEC among the herd, environment, and, subsequently, in slaughtering facilities (Persad and LeJeune, 2014). It has been demonstrated that bovine ruminants can transiently shed STEC between 101 colony-forming unit (CFU)/mL and 109 CFU/mL, a characteristic termed intermittent shedding (Fukushima et al., 1999; Chase-Topping et al., 2008; Menrath et al., 2010; McCabe et al., 2019).
Some animals may shed higher numbers of STEC for prolonged periods, in a phenomenon known as “super-shedding,” and these have been shown to pose the greatest risk of contamination to the agri-food chain (Chase-Topping et al., 2008; Karmali, 2017). A super-shedder has many different definitions that are time and study specific (Matthews et al., 2006b; Chase-Topping et al., 2008). However, it is generally accepted that a super-shedding ruminant is any animal that sheds greater than 104 CFU/g feces (Matthews et al., 2006a, 2006b; Munns et al., 2015; McCabe et al., 2019). Ruminants colonized in the RAJ display a higher propensity for super-shedding when compared with non-colonized animals (Lim et al., 2007; Chase-Topping et al., 2008; Persad and LeJeune, 2014). Both E. coli O157 and non-O157 STEC serogroups have been isolated from super-shedding ruminants (Menrath et al., 2010; Paquette et al., 2018; McCabe et al., 2019).
Factors Impacting STEC Carriage in the Host
Age
Several studies investigating STEC shedding in cattle and sheep have reported that as animals age, especially after weaning, the prevalence of STEC decreases (Chapman et al., 1997; Fagan et al., 1999; Menrath et al., 2010; Mir et al., 2016). This has been attributed to the changing composition of the gut microflora as the animal matures (Mir et al., 2016). A year-long study of sheep in the United States found that younger sheep (22.7%) on a feedlot had a significantly higher fecal prevalence of STEC than older animals (0–1.9%) at pasture (Kilonzo et al., 2011). A long-term study of Australian sheep identified newborn lambs (55.2%) as the highest shedders of STEC, and ewes (32.3%) and weaned lambs (25.8%) as the lowest shedders (Djordjevic et al., 2004). A study carried out at a later date in New Zealand also found that the prevalence of STEC in lambs (3.8%) presented at slaughter was much greater than that reported for ewes (0.9%) at pasture (Moriarty et al., 2011).
Other reports have suggested that the impact of an animal's age on STEC prevalence is unclear (Heuvelink et al., 1998; Evans et al., 2011; Mir et al., 2015; EFSA and ECDC, 2018). One Scottish study reported a significantly higher prevalence of the “ top five” STEC serogroups among slaughtered sheep (10%) than lambs (3%), but did not clarify whether adult sheep had a higher overall STEC prevalence than lambs for all stx-positive E. coli serogroups (Evans et al., 2011). An earlier study in Spain also reported a greater prevalence of STEC in ewes (33.1%) than lambs (13.8%) (Orden et al., 2003). However, this study defined lambs as animals younger than 4 weeks of age. Most other studies classify lambs as animals up to 8 and a half months old, which likely influences the reported prevalence rates (Kudva et al., 1997).
Changes in housing arrangements, differences in rearing practices between surveys, and changes to the animals' diet as it matures could account for a large portion of the observed affect (Gunn et al., 2007). Regular contact with other ruminants and wildlife may also increase the risk of herd shedding or increase the likelihood of a herd becoming infected (Venegas-Vargas et al., 2016; Ahlstrom et al., 2017).
Diet
The animals' diet has been shown to influence STEC colonization and subsequent shedding in both bovine and ovine carriers (Kudva et al., 1997; Diez-Gonzalez et al., 1998; Hovde et al., 1999; Cornick et al., 2000; Chaucheyras-Durand et al., 2010; Mir et al., 2016). Food intake and the fiber content of the fodder provided can exert an impact on the bacterial populations of the gut (Grau et al., 1969; Kudva et al., 1995; Chaucheyras-Durand et al., 2010, 2012). Concentrate diets have been shown to correlate with a higher concentration of STEC shedding in lambs and sheep (Kudva et al., 1995; Lema et al., 2001; Gutta et al., 2009). A longitudinal survey investigating STEC prevalence in lambs fed on concentrates or reared at pasture reported a prevalence range of between 15.6% and 40.6% (Djordjevic et al., 2004). This variation can largely be attributed to the pasture-based diets with significant differences observed between lambs finished at grass (15.6%) and on concentrated feed (40.6%).
Seasonality
There is evidence to suggest that seasonal effects are likely to influence prevalence rates, with several surveys observing seasonal peaks in STEC shedding (Franco et al., 2009; Fraser et al., 2013; Kamel et al., 2015). The observed changes are likely due to the associated changes in temperature.
Higher ambient temperatures, as opposed to increased solar exposure, have been linked with elevated STEC shedding rates (Sheng et al., 2004; Milnes et al., 2008; Tahmasby et al., 2014; Williams et al., 2015). Investigations in bovine hosts have suggested that enhanced transmission rates at elevated temperatures are also influenced by the increased activity of flies, animals drinking more water, and hormonal and behavioral changes in the host (Berry and Wells, 2010; Dawson et al., 2018). Kilonzo et al. (2011) reported that STEC is 3.2 times more likely to be isolated during the higher summertime temperatures, despite not recording any significant differences in shedding between seasons. Higher summertime temperatures have been associated with increased STEC isolation rates in sheep in Great Britain, Egypt, Italy, and the United States (Ogden et al., 2002; Franco et al., 2009; Fraser et al., 2013; Kamel et al., 2015). In addition, Evans et al. (2011) observed clear seasonal differences between the shedding of STEC O157 and O26 in Scottish sheep and failed to detect any STEC O157 isolates between January and March during their study.
Prevalence and Carriage of STEC in Ovine Feces
Ovine fecal samples can be collected in different ways. Live animal feces have been collected from pasture, sheds, or lairage floors. Alternatively, the rectum of the animal may be swabbed. Slaughtered animals may also be swabbed at the rectum pre-evisceration, or samples from the colon, rectum, or ileum may be collected post-evisceration. Fecal contents can also be collected from the lower GI tract after evisceration. Selected surveys on the prevalence of STEC in ovine feces are presented in Table 1 and have been arranged in order of sample type.
Selected a Surveys on the Prevalence of Shiga-Toxin Producing Escherichia coli in Ovine Feces
Surveys were selected based on specific criteria; see the Introduction section for details.
Number of samples examined.
Percentage of samples recorded as positive.
AIMS, automated IMS; ELISA, enzyme linked immunoassay; IMS, immunomagnetic separation; IMViC, indole, methyl red, Voges Proskauer and citrate; MLVA, Multi Locus VNTR Analysis; PCR, polymerase chain reaction; RAJ, recto-anal junction; RAMS, recto-anal mucosal swabs; STEC, Shiga-toxin producing E. coli; VCA, vero cell assay; VLA, Veterinary Laboratories Agency.
From the results presented in Table 1, the fecal prevalence of both STEC O157 and non-O157 STEC varies considerably between surveys. The relative prevalence of STEC O157 is 4.4% and ranges from 0.2% to 28.1%, while the relative prevalence of STEC is 33.3% and ranges from 0.9% to 90.0%. The wide range of reported prevalence rates may be influenced by several factors, including study design, the number of samples collected, and the isolation techniques employed. However, it is more likely that a large portion of the variation is due to intrinsic factors associated with the animal, such as their age, sex, and immune status, as well as extrinsic factors such as the season, diet, and climate (Dewell et al., 2005).
Sample type may also be an important factor when estimating STEC prevalence. Some of the lowest prevalence rates reported for both E. coli O157 and non-O157 STEC were observed in surveys conducted in Brazil, Scotland, New Zealand, and Greece. Each of those surveys collected matter from fecal pats or pellets excreted by lambs or sheep in farmhouses, at pasture, or in abattoirs. The STEC have been shown to have a heterogenous distribution in excreted fecal matter collected from pats in bovine enclosures, and there is a risk of underestimating the true STEC prevalence (Pearce et al., 2004; Robinson et al., 2005). In addition, the increased exposure to the environment after excretion may result in some bacterial decay.
Similar investigations have not yet been carried out in ovine ruminants, but it is an important point to consider when assessing the reduced prevalence observed in these studies. In comparison, studies in both ovine and bovine hosts investigating STEC O157 prevalence have shown that fecal samples (collected via fecal palpitation) and recto-anal mucosal swabs are most suitable for the detection of STEC (Rice et al., 2003; McPherson et al., 2015).
Some of the observed variance could also be due to the wide array of STEC detection and characterization techniques employed among the presented surveys. Molecular techniques such as polymerase chain reaction (PCR) may be at risk of potentially overestimating the true prevalence of STEC due to its inability to differentiate live and dead cells, resulting in DNA amplification from dead STEC cells, and interference from background flora present in the sample (Macori et al., 2020). Other methods, such as immunomagnetic separation (IMS), agglutination techniques, and selective agars, may underestimate certain serogroups or atypical strains due to their high degree of specificity and reliance on the expression of typical STEC phenotypes (Noll et al., 2015; Hallewell et al., 2017). In addition, many surveys employ an enrichment step during the STEC detection protocol. The temperature at which this is carried out may be preferential to certain serogroups and mask the presence of others (Wang et al., 2013; Conrad et al., 2016). In general, it is important to employ a multi-hurdle approach during the detection and culture of an STEC isolate to ensure all typical and atypical strains present in a sample are accounted for. The highest rates of STEC carriage have been observed in ovine herds located in the southern hemisphere.
Several surveys have investigated the prevalence of STEC in Australian sheep and lambs and have observed STEC carriage to be higher than the relative mean reported for STEC (33.3%), and it ranges from 15.6% to 73.3% (Djordjevic et al., 2001, 2004; Yang et al., 2017; Oporto et al., 2019). Interestingly, high prevalence rates were observed across all ovine herds and animals tested, regardless of animal age, diet, or sample type. High STEC prevalence rates have also been observed in herds in Brazil and New Zealand from fecal samples collected on farm (Cookson et al., 2006; Martins et al., 2015). The higher prevalence rates may be influenced by the higher ambient temperatures generally observed in these regions in comparison to other sheep-producing regions in the northern hemisphere (Fleury et al., 2006; Carlton et al., 2016; Philipsborn et al., 2016).
The lowest rates of STEC O157 and non-O157 STEC carriage have generally been observed in European herds. Herds sampled in Great Britain reported an STEC O157 prevalence of between 0.7% and 28.1%. Many of the surveys reported STEC O157 carriage to be below the relative mean of 4.4%, with the exception of one long-term study that recorded an STEC O157 prevalence of 28.1% in ovine feces collected on open farms in England and Wales (Pritchard et al., 2009). Studies involving Spanish herds recorded the highest rates of STEC carriage among ovine herds in Europe. It is likely that the higher prevalence rates can in part be attributed to the age of the lambs when slaughtered. Lambs younger than 3 months of age, often preweaned and milk fed, account for many of the slaughtered lambs in Spain (Campo et al., 2016). In other regions of Europe lambs are much older, often between 6 and 8 months old, before they are sent for slaughter.
A small number of surveys have also investigated the prevalence of a selected number of serogroups among sheep and lambs. The prevalence of the “top five” serogroups (O157, O26, O103, O145, O111) in the surveyed herds is generally lower than the relative mean observed for STEC O157 (4.4%). Overall, there are a limited number of surveys investigating the prevalence of non-O157 STEC serogroups, and only a small number of the wider STEC prevalence surveys have proceeded to serotype the isolated STEC colonies.
STEC Prevalence in Ovine Abattoirs
STEC and ovine fleece contamination
The fleece is a major source of STEC and microbial contamination within slaughtering facilities, and carcass de-pelting has been identified as a critical control point (CCP) for carcass contamination (EFSA Panel on Biological Hazards (BIOHAZ), 2013). Fleeces can also be interchangeably referred to as the pelt or hide. They primarily become soiled along the abdomen, rump, and in the area surrounding the hind legs (Lenahan et al., 2007; Hauge et al., 2011; Thomas et al., 2013). A wide variety of materials may contaminate the fleece, including mud, dust, sand, feces, and soil. Contamination may occur on farm, in sheds, or at pasture, during transport, or when the animal is held in lairage (Hauge et al., 2011). Table 2 presents selected surveys describing the prevalence of STEC on ovine fleeces.
Selected a Surveys on the Reported Prevalence of Shiga-Toxin Producing Escherichia coli on Ovine Pelts, Fleece, or Hides
Surveys were selected based on specific criteria; see the Introduction section for details.
Number of samples examined.
Percentage of samples recorded as positive.
AIMS, automated IMS; IMS, immunomagnetic separation; PCR, polymerase chain reaction; STEC, Shiga-toxin producing E. coli.
From the surveys presented in Table 2, the relative prevalence of STEC O157 on the ovine fleece ranged from 0.8% to 12.8% and had a relative mean of 7.6%. It is interesting to note that the relative prevalence of STEC O157 was much higher on the fleece than what was reported for fecal samples and declines considerably after de-pelting. This can in part be attributed to the fact that the fleece is easily soiled and fecal matter may build up over time. A similar trend was observed in a wide-ranging review of pathogens along the bovine production chain (Rhoades et al., 2009).
Only one study investigating STEC carriage on ovine fleeces was suitable for inclusion in this review and reported a non-O157 STEC prevalence of 86.2% from a sample size of 846 sheep (Kalchayanand et al., 2007). One Irish survey report on the prevalence of the “top five” serogroups on the fleece of Irish sheep noted a much lower STEC prevalence of 1.8% (Thomas et al., 2013).
STEC contamination on ovine carcasses
Carcass dressing immediately follows fleece removal or de-pelting. First, the gut contents are discarded during the evisceration process (EFSA Panel on Biological Hazards (BIOHAZ), 2013; FAO, 2018). Evisceration is a major CCP for the transfer of bacteria from the intestines to the carcass, with various reports citing an increase in bacteria, including E. coli, being transferred to the carcass surface at this point (Alonso et al., 2007; Sheridan, 2007; Milios et al., 2011). The carcass may then be subjected to numerous interventions, which differ depending on the legislation employed in each specific region, before it is chilled, boned, and processed into retail cuts (EFSA Panel on Biological Hazards (BIOHAZ), 2016). Selected surveys on the prevalence of STEC on de-pelted ovine carcasses and meat samples collected from abattoirs are presented in Table 3.
Selected a Surveys on the Prevalence of Shiga-Toxin Producing Escherichia coli on Ovine Carcasses at Different Stages of Abattoir Processing
Surveys were selected based on specific criteria; see the Introduction section for details.
Number of samples examined.
Percentage of samples recorded as positive.
AIMS, automated IMS; IMS, immunomagnetic separation; N. A, not available; PCR, polymerase chain reaction.
The prevalence of E. coli O157 and non-O157 STEC declines after abattoir processing and the application of suitable interventions. A relative mean prevalence of 2.1% and a range of 0.2% to 9.8% is reported for STEC O157 on ovine carcasses from the studies shown in Table 3. The relative mean prevalence for all STEC on ovine carcasses was 58.7% and ranged from 2.0% to 81.6%.
From the surveys presented in Table 3, the reported prevalence rates were affected in some instances after the application of intervention processes to reduce or arrest the growth and proliferation of STEC and other microbes on the carcass surface. The STEC prevalence rates were largely unchanged after carcass chilling. A small reduction in STEC O157 prevalence was observed in a survey of Ethiopian ovine carcasses after carcass washing (Mersha et al., 2010). Evisceration of the carcass resulted in a marginal increase in both STEC O157 and non-O157 STEC on ovine carcasses sampled during a large survey of lamb carcasses in three American abattoirs, despite the application of antimicrobial interventions (Kalchayanand et al., 2007). However, this study also reported a reduction between 2 and 2.5 log CFU/100 cm2 in indictor bacteria and Salmonella after the application of intervention processes and carcass chilling. The reduced interference from background flora likely leads to easier STEC isolation, particularly as the study used molecular methods such as PCR and IMS, and is an important point to consider when analyzing STEC prevalence at different processing stages.
STEC contamination in raw ovine meat
Raw meat products are defined as raw meat mixes composed of muscle meat and sometimes animal fat. The products have not been cured or processed but may occasionally contain nonmeat products, such as flavourings or fruits (FAO, 2018). These products may include trimmings, offal, and butcher cuts such as steaks, mince, and chops (Rhoades et al., 2009). Contamination of raw meat products generally occurs due to carcass contamination and the subsequent dissemination of the pathogen from the kill line to the boning hall. Selected surveys on the prevalence of STEC in raw, unprocessed ovine meat products are presented in Table 4.
Selected a Surveys on the Prevalence of Shiga-Toxin Producing Escherichia coli in Raw Unprocessed Ovine Meat Products
Surveys were selected based on specific criteria; see the Introduction section for details.
Number of samples examined.
Percentage of samples recorded as positive.
IMS, immunomagnetic separation; PCR, polymerase chain reaction; STEC, Shiga-toxin producing E. coli.
From the surveys presented in Table 4, STEC O157 had a relative mean prevalence of 1.9% and ranged from 0.2% to 6.3% in raw ovine meat. A relative mean prevalence of 15.4% and a range between 2.7% and 35.5% was reported for STEC. It is interesting to note that the prevalence of STEC O157 on raw ovine products is similar to that observed for ovine carcasses (2.1%), whereas the observed STEC prevalence is much lower. The weighted mean prevalence observed for STEC O157 is likely an inflated rate and may be influenced by the low number of surveys available for inclusion, as well as the variable hygiene standards in the sampled premises. However, the low prevalence rates are indicative of how effective application of postharvest intervention processes in slaughtering facilities has been in stopping the spread of the pathogen to raw meat products and subsequently to the food supply chain.
Overall, the lowest STEC prevalence rates were observed in raw ovine meat and meat products. No significant differences were observed in STEC prevalence between lamb or mutton meat or meat mixes. However, as was the case for the observed fecal prevalence, some of the highest prevalence rates for STEC were observed in the southern hemisphere. One survey in Australia reported an STEC prevalence of 40.0% on lamb cutlets collected from butchers' premises, whereas a later study recorded a prevalence of 55.6% in raw mutton collected from supermarkets and farmers markets in China (Barlow et al., 2006; Bai et al., 2016). Other surveys have reported much lower prevalence rates (2.7–22.3%) for lamb and mutton meat sampled in India and Iran (Momtaz et al., 2013; Kumar et al., 2014; Tahmasby et al., 2014). The detection techniques employed by each research group may influence isolation rates, as molecular techniques may over-estimate the true STEC prevalence. In addition, the sample type may also influence the observed prevalence, with some areas of the carcass being more susceptible to potential STEC contamination than others.
Discussion
It is clear from these data that the prevalence of both STEC O157 and non-O157 STEC generally decreases along the ovine production chain. The highest rates of carriage are observed on the ovine fleece, with the prevalence decreasing on the carcass and in raw ovine meat, after carcass dressing and abattoir processing. However, as previously discussed, the small number of surveys that have investigated STEC prevalence in the ovine fleece, carcass, and raw meat means that these figures should be interpreted with caution.
In general, there is a lack of information regarding the prevalence of STEC among ovine ruminants, as most studies have only investigated the carriage of STEC O157. This is particularly concerning, as it is widely accepted that ovine animals host a broad range of non-O157 STEC. To compound this, many of the studies have employed qualitative methodologies, and though useful, have added little to the understanding of the shedding patterns of ovine ruminants, particularly the “super-shedding” dynamics within ovine herds.
Super-shedding ruminants have been demonstrated to excrete high numbers of STEC and have a higher risk of disseminating the pathogen among the herd and within slaughtering facilities. Studies investigating the shedding of STEC O157 in cattle reported that 8% and 9% of the respective herds were classified as super-shedding individuals, but were responsible for 96% and 99% of the total bacteria shed (Omisakin et al., 2003; Chase-Topping et al., 2007). A quantitative study of a Scottish sheep flock identified some individual animals shedding up to 106 CFU g−1 (Ogden et al., 2005). It is likely that STEC exhibits discrete super-shedding dynamics in ovine ruminants, which are anticipated to be distinct from what is observed in other prominent ruminant hosts. Additional investigations on the quantitative shedding of STEC in ovine ruminants and the risk they may pose to the food supply chain are, therefore, required.
There is a noticeable lack of information regarding the prevalence of STEC within the boning hall and in raw processed ovine meat products, with only five surveys suitable for inclusion in this review. A smaller survey conducted in the Dunedin region of New Zealand reported an STEC prevalence of 17.1% from 37 sampled lamb and mutton meat products from butchers and supermarkets premises (Brooks et al., 2001). An Indian study also reported a high STEC prevalence of 40.0% from 50 mutton meat samples (Kiranmayi, 2010). A number of disease cases have also been linked to the consumption of contaminated ovine produce (Espié et al., 2006; Sekse et al., 2013; Bekal et al., 2014; Wilson et al., 2018). In addition, while compiling this review, only a handful of studies could be found that investigated the prevalence of STEC among ovine ready-to-eat products. These reports were not included in calculations for this review due to the small number of samples tested.
Conclusion
Ovine ruminants are a significant STEC reservoir, and further investigations into the colonization and shedding patterns of STEC within this ruminant population are required to improve our understanding on the shedding dynamics of the pathogen in ovine hosts. Despite the relatively high fecal carriage rate of STEC in ovine ruminants, the prevalence of this pathogen declines significantly along the ovine production chain after the application of suitable postharvest interventions and proper carcass dressing. The small number of surveys investigating the prevalence of STEC in raw ovine products is concerning. However, the low prevalence rates reported in raw meat produce are reassuring as to the potential risk of contamination these products may pose to the food supply chain.
Footnotes
Acknowledgment
The authors thank Paula Reid for assisting with the statistical analysis of the data.
Disclosure Statement
No competing financial interests exist.
Funding Information
This project was funded by the Food Institutional Research Measure, administered by the Department of Agriculture, Food and the Marine, Ireland (Grant Number 15/F/629).
Supplementary Material
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.