
Abstract
Streptococcus suis remains a serious threat to the worldwide swine industry and human health. In this study, rapid assays for the detection of three common virulence-related factors (mrp, epf, and sly) were developed, evaluated, and applied. Loop-mediated isothermal amplification (LAMP) primers were designed using Primer Explorer V5 software. The sensitivity and specificity of the LAMP assays were determined based on sample turbidity. For all three genes, LAMP assays were performed at 62°C with a reaction time of 60 min. The detection limit of conventional polymerase chain reaction (PCR) was 1 ng/μL, 10 pg/μL, and 100 fg/μL for the epf, sly, and mrp genes, respectively. For the LAMP assays, the detection limits were 10 pg/μL, 10 fg/μL, and 100 fg/μL for epf, sly, and mrp, respectively, representing sensitivities 100–1000 times higher than those of the PCR assay. Furthermore, when the LAMP assays were applied to clinical strains, the results were consistent with those of the PCR assay, confirming the LAMP assays as rapid and reliable detection techniques. In conclusion, the LAMP assays described in this study have the potential to become standard methods to detect the virulence factors mrp, epf, and sly. To the best of our knowledge, this is the first study to report the application of LAMP to detect the mrp, epf, and sly genes.
Introduction
As a normal inhabitant of the upper respiratory tract of pigs, Streptococcus suis causes a variety of clinical manifestations, such as meningitis, arthritis, endocarditis, polyserositis, and septicemia in piglets and growing pigs (Staats et al., 1997). S. suis is also a zoonotic pathogen and causes septicemia and meningitis in humans involved in the pork industry, particularly in Southeast Asia (Huong et al., 2014; Dutkiewicz et al., 2018). S. suis has gained increased attention after the two large infection outbreaks that occurred in 1998 and 2005 in China (Du et al., 2017), which revealed the enhanced zoonotic transmission and virulence of S. suis to humans.
In the past few years, research on S. suis has mainly concentrated on its virulence, and large amounts of data have demonstrated that the disease is associated with the serotype and the potential virulence-associated factors. In previous reports, 35 capsular serotypes have been identified for S. suis, but in recent years, S. suis serotypes 20, 22, and 26 are reclassified as Streptococcus parasuis (Nomoto et al., 2015), and the serotype 33 of S. suis is reclassified as Streptococcus ruminantium (Tohya et al., 2017). Among these serotypes, serotype 2 (SS2) is the most common serotype, and most likely to express virulence. Screening of virulence-associated factors of S. suis focused on the presence of the genes mrp, epf, and sly, encoding the virulence-associated markers muraminidase-released protein (MRP), extracellular protein factor (EF), and suilysin (SLY), respectively (Fittipaldi et al., 2009; Li et al., 2012; Wongsawan et al., 2015). Although there is limited evidence of the importance of these factors in the virulence of S. suis, they have shown positive associations with specific serotypes and clinical signs of S. suis infection (Gottschalk et al., 2007).
Conventional methods, such as polymerase chain reaction (PCR) (Zhang et al., 2015; Oh et al., 2017) and multiplex PCR (Silva et al., 2006; van der Wal et al., 2018) have been successfully used to detect the mrp, epf, and sly genes. However, the loop-mediated isothermal amplification (LAMP) method provides several advantages over PCR-based methods. It is capable of amplifying DNA within 60 min with high specificity and sensitivity under isothermal conditions at 60–65°C (Notomi et al., 2000; Nagamine et al., 2002), allowing an incubator or water bath to be used, opposed to a thermal cycler, and thereby reducing the barriers to implementation of this molecular amplification method in resource-limited settings. To date, several LAMP assays have been developed to detect S. suis, SS2, and 89K pathogenicity island (PAI) (Huy et al., 2012; Zhang et al., 2013; Arai et al., 2015). However, no LAMP assay to detect virulence-associated factors in S. suis has yet been reported.
To improve and accelerate the detection of S. suis strains that harbour these virulence genes, a LAMP method was developed in this study. Furthermore, we evaluated the specificity and sensitivity of the method by comparison with the PCR method and its application for the rapid screening of clinical isolates. To our knowledge, this is the first report of a rapid, specific, and sensitive method for the detection of the mrp, epf, and sly genes by LAMP assay in S. suis.
Materials and Methods
Bacterial strains and genomic DNA extraction
Three S. suis clinical isolates (S37, sly positive; S41, epf positive; and S78, mrp positive) were isolated from pig farms in Guangdong Province, China, and used to optimize the LAMP reaction system. Five standard bacteria (including Escherichia coli [ATCC 25922], Salmonella enterica [ATCC 13076], Staphylococcus aureus [ATCC 25923], Enterococcus faecalis [ATCC 29212], and Pseudomonas aeruginosa [ATCC 27853]), three S. parasuis reference strains (serotypes 20, 22, and 26), one S. ruminantium reference strain (serotype 33), and the aforementioned three S. suis clinical isolates were used to evaluate the specificity of the LAMP assays. The 60 other known S. suis clinical strains were also included in this study to apply the established LAMP assays for detection. These nonduplicate S. suis isolates were isolated from pigs in Guangdong Province, China, between 2008 and 2010. All of the S. suis strains were typed for serotypes (1, 2, 7, and 9) by multiplex PCR as described previously (Wisselink et al., 2002). Details of these strains could be found in Supplementary Table S1.
A bacteria genome extraction kit [Tiangen Biotech (Beijing) Co., Ltd.] was used for bacterial DNA extraction. All procedures were performed according to the manufacturer's instructions.
Primer design
Primers for mrp, epf, and sly LAMP reactions were designed based on the National Center for Biotechnology Information (NCBI) database sequences of mrp (1071 bp, GenBank No. NC_012926.1), epf (4376 bp, GenBank No. X71881.8), and sly (1494 bp, GenBank No. NC_012926.1). Using Primer Explorer V4 software (
Primers Used in This Study
Using this primer, the amplification products of the different variants were obtained: 747 bp for mrps , 1148 bp for mrp, 1556 bp for mrp* , ∼1600 bp for mrp** , ∼2000 bp for mrp*** , and ∼2400 bp for mrp**** .
Using the same primer, different variants of epf could be obtained: 626 bp for epf, and the amplicon size of epf * were 1278, 1505, 2313, 2537, and 2993 bp.
LAMP, loop-mediated isothermal amplification; PCR, polymerase chain reaction.
LAMP reaction
The LAMP assays were carried out in a reaction mixture with a total volume of 25 μL containing 40 pmol (each) of the primers FIP and BIP, 5 pmol (each) of the primers F3 and B3, 20 pmol of the loop primer, 12.5 μL of 2 × reaction mix, 1.0 μL (8 U) of Bst DNA polymerase and 2.0 μL of target genomic DNA by using the Loopamp DNA Amplification kit (Eiken Chemical Co., Ltd., Shanghai, China).
Optimization of LAMP assays
For determination of the optimum primers, the reaction tube was incubated at 65°C by adding inner primers FIP/BIP and outer primers F3/B3 for 60 min in the Loopamp Real-time Turbidimeter (LA-320c; Eiken Chemical Co. Ltd.), followed by heating to 80°C for 5 min. The primers with the shortest times and highest amplification efficiencies were used in the subsequent optimization assays. Then, the LAMP assays were performed together with the loop primer to ensure the shortest optimal reaction time for amplification.
To determine the most efficient conditions for the mrp, epf, and sly LAMP reactions, a range of reaction temperatures (60–67°C) were tested. The conditions that yielded turbidity at an optical density of 0.1 in the least amount of time and high peak turbidity curve values were considered optimal.
Sensitivity of the LAMP assay
The concentrations of the genomic DNA of each isolate was measured by Nanodrop 2000 (Bio-Rad) and adjusted to 10 ng/μL with distilled water. The sensitivity of each LAMP assay was determined using 10-fold serial dilutions, ranging from 1 ng/μL to 1 fg/μL (10−6 ng/μL), respectively, and a negative control was performed using sterile distilled water. In addition, PCR assays were performed using the same DNA templates. These experiments were done in duplicate.
Results
Primer screening experiment
As shown in Figure 1, among the primer sets, the mrp-3L, epf-1L, and sly-3L primer sets initiated amplification of the target genes in the shortest time. Through the specificity experiments, false positive amplification was observed, so we employed mrp-2L instead and designed a loop primer for mrp-2L. Then, the specificity experiments were carried out with the mrp-2L and mrp-2L+LB, and no amplification was obtained for other nine strains except for S78. In addition, the turbidity curves indicated that the two primer sets (mrp-2L+LB and mrp-3L) began to amplify the mrp gene at the same time (data not shown). Therefore, the mrp-2L+LB, epf-1L+LB, and sly-3L were chosen as the optimal primer sets for LAMP detection in S. suis (Table 1).
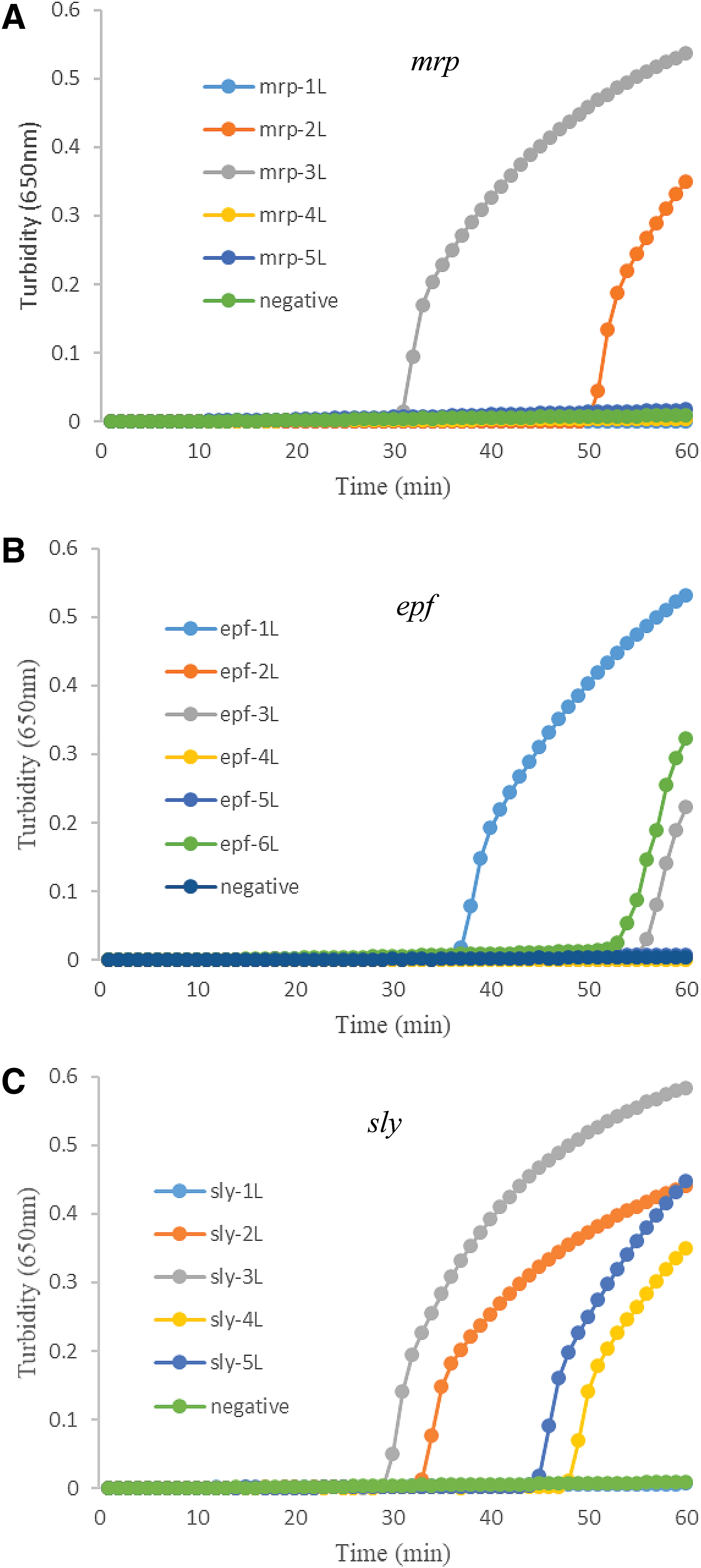
Primer screening of the LAMP assays for mrp, epf, and sly genes (
Optimization of temperature for the LAMP assay
As demonstrated in Figure 2, eight curves were produced for temperatures in the range of 60–67°C. For the mrp gene, the shortest reaction times were observed at 62°C and 63°C (Fig. 2A); for the epf and sly genes, the shortest reaction times were observed at 61°C, 62°C, and 63°C (Fig. 2B, C). As the first and the highest peaks were observed at 62°C, this temperature was selected as the best reaction temperature for these three genes.
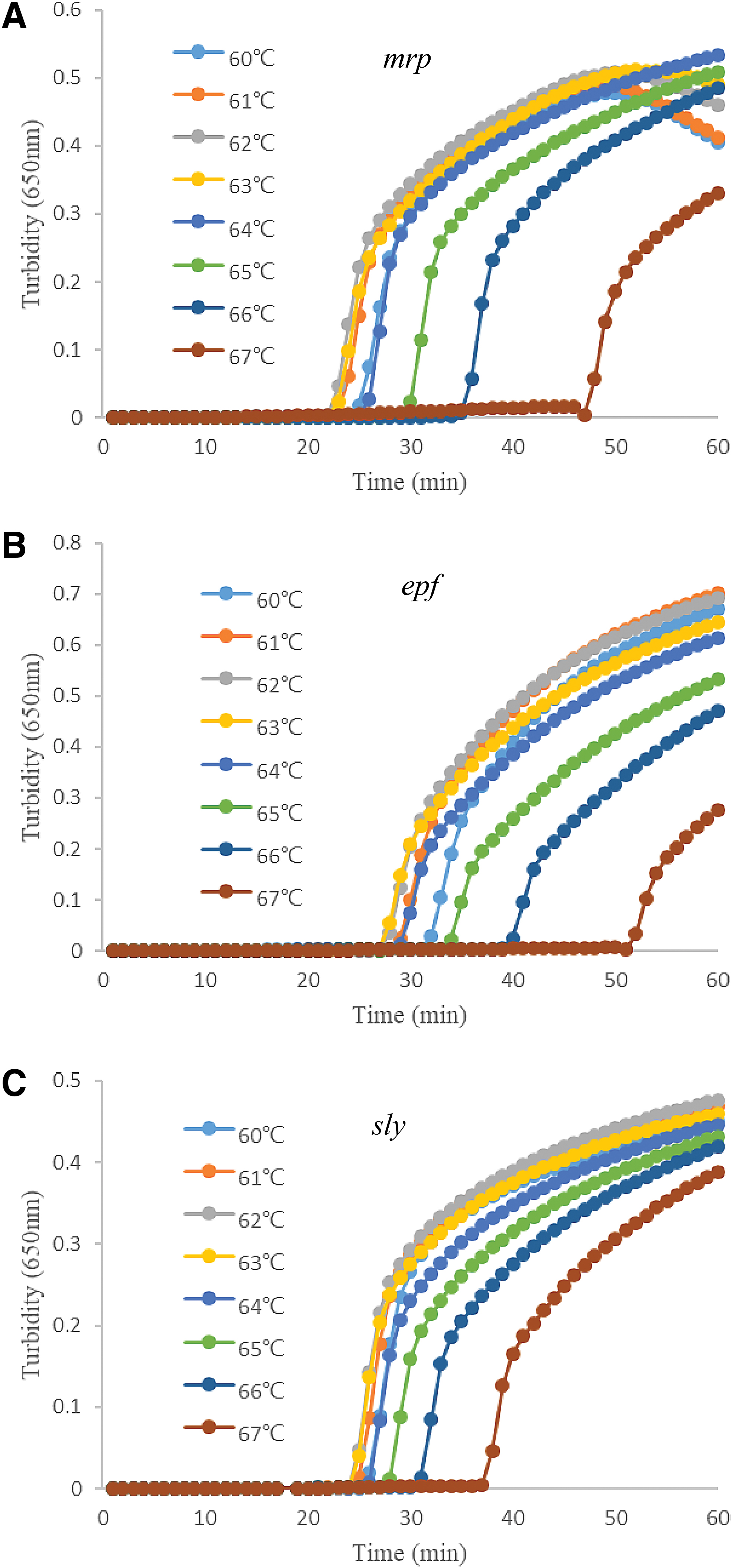
Optimal temperature of LAMP assays for mrp, epf, and sly genes (
Comparison detection limits of LAMP and PCR assays
Comparison of the detection limits of the three LAMP assays with those of conventional PCR, 10-fold serially diluted genomic DNA samples were used. As shown in Figure 3A–C, the detection limits for the LAMP assays were 100 fg/μL (1.5 × 103 copies/μL), 10 pg/μL (1.2 × 104 copies/μL), and 10 fg/μL (2.8 × 107 copies/μL) for mrp, epf, and sly, respectively. In contrast, the detection limits of PCR were 100 fg/μL, 1 ng/μL, and 10 pg/μL for mrp, epf, and sly, respectively (Fig. 3D–F). Therefore, the sensitivities of the LAMP assays for epf and sly were 100–1000 times higher than those of the PCR.
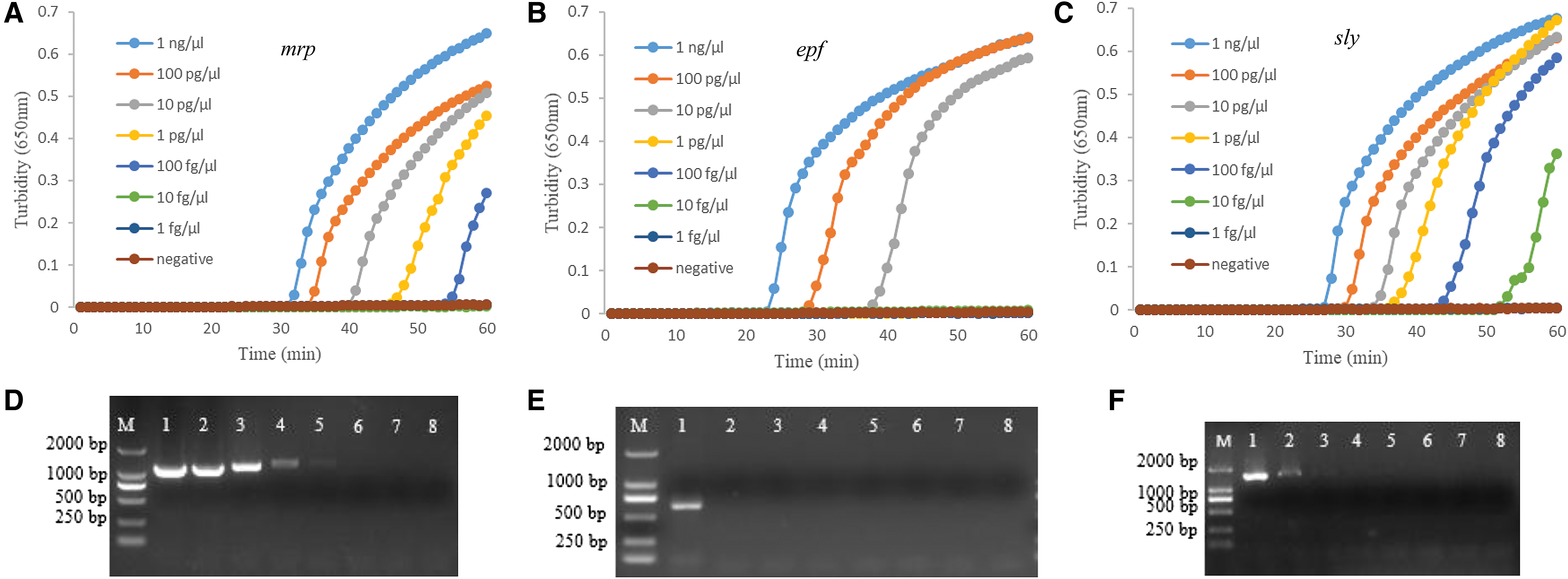
Sensitivity of LAMP and PCR assays for mrp, epf, and sly genes. Various amounts of template DNA were used in the LAMP reaction and PCR.
At a template concentration of 1 ng/μL, the reaction time was 33, 25, and 29 min for mrp, epf, and sly, respectively. Furthermore, the reaction time was 57 min for 100 fg/μL in the case of mrp, 40 min for 10 pg/μL in the case of epf, and 56 min for 10 fg/μL in the case of sly (Fig. 3A–C). The reaction time here refers to the time needed for the turbidity of each sample to exceed 0.1 in the turbidity curve.
Specificity of the LAMP assays
The results of the specificity experiment showed that the mrp gene was detected only in the mrp-positive strain; no false positive amplification was observed for the other five standard bacteria. Similar results were obtained for the epf and sly genes.
Application of LAMP assays on clinical isolates
To evaluate the accuracies of the LAMP assays, 60 clinical strains from a previous study were analyzed. Among the 60 clinical strains, 44 strains (73%) were positive for mrp, 33 strains (55%) were positive for epf, and 39 (65%) were positive for sly. These results were consistent with the PCR results (Table 2). Seven phenotypes were identified, and the most prevalent genotype was mrp+epf+sly+ (29/60, 48.33%), followed by mrp−epf−sly− (21.67%) and mrp+epf-sly+ (13.33%). The other profiles identified in this study were mrp+epf−sly−, mrp+epf+sly−, mrp−epf−sly+, and mrp−epf+sly− (Table 3).
Comparison of the Results of the Two Methods for Detection of Clinical Isolates
Using mrp-F and mrp-R, only the result of 1148 bp is shown here. In addition, three strains carried mrp s variant.
Using epf-F and epf-R, only the result of 626 bp is shown here. In addition, there was no variant of epf gene in this study.
Distribution of Different Genotypes Among the 60 Clinical Isolates
Discussion
S. suis is responsible for important economic losses in the swine industry worldwide and is a zoonotic pathogen that can be transferred from pigs to humans. According to the results of two previous studies (de Greeff et al., 2011; Dong et al., 2015), the researchers concluded that epf and sly can be considered virulence marker genes for SS2. In another report, three virulence-associated factors (mrp, epf, and sly) were found to be significantly associated with SS2 (Li et al., 2012). In addition, multilocus sequence typing (MLST) has been used worldwide to determine the sequence types (STs) of S. suis strains, and global MLST studies identified ST1, ST25, and ST28 as the most prevalent STs in swine (de Greeff et al., 2011; Schultsz et al., 2012), but in China, ST7 was predominant, and followed by ST1 (Huang et al., 2015). One study found that significant correlation existed between virulence-associated gene profile VP26 (mrp+epf+sly+impdh+covR+StrA+dpp+dltA+saoM) and ST1 (p < 0.05), and VP26 was detected in 71.4% of the SS2 clinical pig isolates (Sanchez del Rey et al., 2016). Therefore, it is important to establish a convenient method for the rapid detection of these three genes.
In this study, LAMP assays were designed to detect three virulence-associated factors (mrp, epf, and sly). To reduce the reaction time, loop primers (LB) were added for mrp and epf; with the loop primers, the reaction was 15–20 min faster than in their absence (Nagamine et al., 2002). This feature is very important for rapid detection. Consequently, three LAMP assays could be finished within 60 min, and the time should be shorter for epf gene (about 40 min). In addition, compared with conventional PCR, the LAMP assays have shorter reaction times and in some cases increased sensitivity, and they do not require a gel electrophoresis step or expensive instruments, being performable with Bst DNA polymerase under constant conditions. Therefore, it is possible to utilize this method as a rapid test for detecting virulence-producing strains.
When we used the LAMP assays to screen for three virulence factors in S. suis isolates, we found that the most predominant gene was mrp (73%), followed by sly (65%) and epf (55%). This prevalence of three virulence factors among these isolates is slightly higher than that reported in two previous studies in Korea and Spain (Sanchez del Rey et al., 2016; Oh et al., 2017), and lower than that reported in other studies in China (Dong et al., 2015; Zhang et al., 2015). In addition, the virulence-associated gene profiling showed that mrp+epf+sly+ was the major genotype in this study. This finding is consistent with the results of two other studies in China (Wei et al., 2009; Zhang et al., 2015) but inconsistent with those of studies in other countries (Sanchez del Rey et al., 2014; Meekhanon et al., 2017), indicating that the genotype of virulence varies among different countries.
The results of LAMP assays in this study were detected by real-time turbidimetry of LAMP reactions. However, since 2001, researchers have found that during LAMP reactions, high levels of magnesium pyrophosphate form (Mori et al., 2001), such that the results can be observed by the naked eye. In this study, we also made a preliminary assessment based on the level of turbidity observed by naked eye (data not shown); this approach may enhance the feasibility and application of the LAMP method.
Conclusions
LAMP assays for the detection of virulence genes (mrp, epf, and sly) were established and demonstrated to have advantages of sensitivity, speed, and ease of operation. The test can be used for the detection and genotyping of virulence in S. suis cilinical isolates, which will be a useful tool for the molecular characterization of S. suis for epidemiological purposes. Therefore, LAMP assays have potential as a novel and standardized method in routine clinical investigation.
Footnotes
Acknowledgments
We thank Professor Yahong Liu in South China Agricultural University for providing us S. suis clinical strains used in this study. In addition, we are grateful to Dr. Kaicheng Wang (China Animal Health and Epidemiology Center, Qingdao, China) for kindly providing the genomic DNA of four reference strains (serotypes 20, 22, 26, and 33).
Disclosure Statement
No competing financial interest exist.
Funding Information
This study was supported by the National Key Research and Development Program of China (2016YFD0501301, 2018YFD0500500), and the Major National Science and Technology Program (2018ZX10733402-013).
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.