
Abstract
Salmonella Enteritidis is an important foodborne pathogen with high prevalence of resistance to cephalosporins, imposing a serious threat to public health. Therefore, a total of 162 Salmonella Enteritidis isolates collected from child patients in China from 2007 to 2017 were characterized for their resistance to cephalosporins and investigated the transmission characteristics of cephalosporin resistance gene. We found that 15 (9.26%) isolates were all resistant to cefalotin (minimum inhibitory concentration [MIC] ≥512 μg/mL), ceftazidime (MIC 16–128 μg/mL), ceftriaxone (MIC 64 to ≥512 μg/mL), ceftiofur (MIC 64–256 μg/mL), and cefotaxime (MIC 64 to ≥512 μg/mL) with the possession of cephalosporin resistance genes bla CTX-M-55 (n = 13), bla CTX-M-101 (n = 1), and bla CTX-M-153 (n = 1). Molecular typing further revealed that these 15 isolates belonged to sequence type ST11 and shared close pulsed-field gel electrophoresis patterns, suggesting the possibility of clonal spread in Salmonella Enteritidis interspecies. Furthermore, conjugation experiments were successfully performed in 13 of 15 isolates, and bla CTX-M-55 was present on conjugative plasmids with sizes ranging from 54.7 to 173.4 kb. Compared with recipient Escherichia coli C600, transconjugants conferred elevated MICs for cephalosporins ranging from 2- to 2048-fold. The genetic structure surrounding of bla CTX-M-55 gene in transconjugants were ΔISEcp1-bla CTX-M-55-orf477 (n = 8) and ISEcp1-bla CTX-M-55-orf477 (n = 3), respectively. Taken together, bla CTX-M on the plasmids might contribute to cephalosporin resistance in Salmonella Enteritidis, and conjugative transfer of bla CTX-M-55 might facilitate the spread of cephalosporin resistance in Salmonella Enteritidis. Hence, effective mitigation measurements are needed to reduce the threat caused by cephalosporin-resistant Salmonella Enteritidis to public health.
Introduction
S
Salmonella resistance to extended-spectrum cephalosporins is normally associated with the production of extended-spectrum cephalosporinases especially CTX-M (Su et al., 2004). There are currently 150 known bla CTX-M subtypes, of which bla CTX-M-55 has been increasing in China, Korea, and Thailand (Kim et al., 2017; Luk-in et al., 2018; Zhang et al., 2019). In China, bla CTX-M-55 has surpassed bla CTX-M-15 and become the second most frequently identified genotype among human and animal isolates (Lv et al., 2013). More importantly, bla CTX-M is usually located on conjugative plasmids that facilitate its transfer among Enterobacteriaceae (Michael et al., 2006). The primary conjugative plasmid families in which bla CTX-M-55 has been found include IncF (Wang et al., 2018), IncI1 (Xia et al., 2017), IncN (Fu et al., 2020), and IncHI2 (Zhang et al., 2019). In addition, an ISEcp1 transposition unit is often associated with the transfer, mobilization, and expression of bla CTX-M-55 (Qu et al., 2014; Sun et al., 2016).
Despite a number of studies regarding the prevalence and dissemination of cephalosporin resistance in Salmonella from adults and animals, limited information concerning cephalosporin resistance for infections of children is available (Chao et al., 2017; Ma et al., 2018; Feng et al., 2019; Kijima et al., 2019). In our study, we collected Salmonella Enteritidis isolates from children younger than 5 years in China to identify their cephalosporin resistance levels and to determine the transferability of bla CTX-M. This information is expected to provide basis to the consideration of cephalosporin treatment of pediatric Salmonella Enteritidis infections and the development of effective mitigation measurements for cephalosporin-resistant Salmonella Enteritidis.
Materials and Methods
Identification of Salmonella isolates
A total of 162 Salmonella Enteritidis isolates were recovered from stool samples of children younger than 5 years with diarrhea during 2007–2017 in Shanghai, Xinjiang, Guangxi, Fujian, Inner Mongolia, Henan, Shanxi, Chongqing, and Zhejiang, China that were provided by the Shanghai Municipal Center for Disease Control and Prevention. These children consumed contaminated food before visiting hospital according to preliminary information. Salmonella isolates were identified using API20E test strips (bioMerieux) and serotyped using a commercial antiserum (Statens Serum Institute, Copenhagen, Denmark) according to the manufacturer's guidelines.
Antimicrobial susceptibility testing and polymerase chain reaction detection of blaCTX-M gene
Cefotaxime (CTX)-resistant Salmonella isolates were selected on Mueller–Hinton agar plates containing 2 μg/mL cefotaxime. Antimicrobial susceptibility testing for cefotaxime-resistant isolates and transconjugants was determined using the agar dilution method according to the Clinical and Laboratory Standards Institute (CLSI) standards (CLSI, 2019). The antimicrobials tested included first-generation (cefalotin [CEF]), second-generation (cefoxitin [FOX]), third-generation (cefotaxime, ceftazidime [CAZ], ceftriaxone [CRO], ceftiofur [TIO]), and fourth-generation (cefepime [FEP]) cephalosporins as well as meropenem, nalidixic acid (NAL), trimethoprim–sulfamethoxazole (SXT), amikacin, ampicillin (AMP), gentamicin, ciprofloxacin, kanamycin (KAN), azithromycin, ofloxacin, tetracycline (TET), chloramphenicol (CHL), and streptomycin (STR). Minimum inhibitory concentration (MIC) for each cephalosporin was determined by antimicrobial susceptibility tests. Escherichia coli ATCC 25922 and Enterococcus faecalis ATCC 29212 were used for quality control strains.
The bla
CTX-M gene was identified using polymerase chain reaction (PCR) as previously reported (Xu et al., 2005) and PCR amplicons were commercially sequenced (Shanghai Sunny Biotechnology, Shanghai, China) for confirmation and further analyzed by Basic Local Alignment Search Tool (BLAST;
Pulsed-field gel electrophoresis and multilocus sequence typing analysis
Genetic relationship between cephalosporin-resistant isolates was determined using pulsed-field gel electrophoresis (PFGE) with XbaI digestion as previously described (Ribot et al., 2006). In brief, bacterial DNA in agarose plugs was digested with 50 U XbaI (Takara, Dalian, China) for 3 h and DNA fragments were separated using a CHEF-Mapper XA PFGE system (Bio-Rad, Hercules, CA) with the following parameters: run time, 19 h; initial switch time, 2.16 s; final switch time, 63.8 s. PFGE patterns were clustered using BioNumerics Software, version 7.6 (Applied Maths, St-Martens-Latem, Belgium). Salmonella Braenderup H9812 universal size standard was used as the PFGE molecular size marker (Hunter et al., 2005).
Multilocus sequence typing (MLST) analysis was carried out by PCR analysis of the housekeeping genes aroC, dnaN, hemD, hisD, purE, sucA, and thrA (Ma et al., 2018). Amplicons were sequenced (housekeeping genes) and the sequencing data were submitted to the MLST website (
Conjugation experiments and plasmid analysis
The transferability of the bla CTX-M gene was examined by conjugation of cefotaxime-resistant Salmonella donors and rifampicin-resistant E. coli C600. In brief, E. coli C600 and Salmonella isolates were combined (4:1) and added to 0.22 μm pore membranes that were then placed on LB agar and incubated for 18h. The overnight culture was then spread on LB agar containing rifampin (200 μg/mL) and cefotaxime (2 μg/mL), followed by incubation for 18 h. Single colonies were then picked and inoculated in LB broth with rifampin (200 μg/mL) and cefotaxime (2 μg/mL) for 8 h. All incubations were carried out at 37°C.
Plasmid replicon typing of transconjugants and Salmonella donor isolates was performed using a multiplex PCR-based method as described previously (Carattoli et al., 2005). The sizes of plasmids in transconjugants and Salmonella donor isolates were determined using S1-PFGE (Zhang et al., 2020).
Genetic structure adjacent to the blaCTX-M gene
The bla CTX-M-55 is often flanked by the insertion sequence ISEcp1 upstream and orf477 downstream (Zhao and Hu, 2013; Qu et al., 2014). We therefore determined the presence of ISEcp1 and orf477 in parental Salmonella donor isolates and transconjugants by PCR mapping to characterize the genetic structure surrounding the bla CTX−M gene as previously described (Hu et al., 2018).
Phylogenetic analysis using PFGE pattern
Dice coefficient was used to compare the obtained PFGE fingerprints, whereas cluster was used to the unweighted-pair group method with arithmetic mean. Patterns indistinguishable by BioNumerics Software and visual inspection were assigned the same pattern designation, and the tolerance was set to 1.3%. According to 97% similarity, different PFGE pattern was marked with different letters.
Results
Antimicrobial susceptibility phenotypes and identification of the bla CTX-M gene
Of the 162 Salmonella Enteritidis isolates from children during 2007–2017 in China, 15 (9.26%) isolates were resistant to CTX (MIC ≥64 μg/mL) in antimicrobial susceptibility testing. All 15 isolates also exhibited resistance to CEF (MIC ≥512 μg/mL), CAZ (MIC 16–128 μg/mL), CRO (MIC 64 to ≥512 μg/mL), TIO (MIC 64–256 μg/mL), and CTX (MIC 64 to ≥512 μg/mL). The 15 isolates were sensitive to FOX (MIC 8–16 μg/mL). In particular, 12 of 15 isolates exhibited resistance to the fourth-generation cephalosporin cefepime with MICs ranging from 16 to 32 μg/mL. These 15 cephalosporin-resistant isolates also exhibited resistance to NAL, SXT, AMP, as well as KAN (9/15, 60.00%), STR (7/15, 46.67%), TET (4/15, 26.67%), and CHL (4/15, 26.67%). All were susceptible to meropenem, amikacin, gentamicin, azithromycin, ciprofloxacin, and ofloxacin. Of note, these 15 cephalosporin-resistant isolates were resistant to at least 4 antimicrobial classes, and 3 isolates (SJTUF14365, SJTUF14920, and SJTUF14971) among them were resistant to 7 antimicrobial classes. The antimicrobial resistance profile NAL-SXT-AMP–KAN -CEF-CAZ-CRO-CTX-TIO-FEP most frequently occurred in five isolates (Table 1).
Characteristics of 15 Cephalosporin-Resistant Salmonella Enteritidis ST11 Isolates
AMP, ampicillin; CAZ, ceftazidime; CEF, cefalotin; CHL, chloramphenicol; CRO, ceftriaxone; CTX, cefotaxime; FEP, cefepime; FOX, cefoxitin; KAN, kanamycin; NAL, nalidixic acid; ND, not determined; PFGE, pulsed-field gel electrophoresis; ST, sequence type; STR, streptomycin; SXT, trimethoprim–sulfamethoxazole; TC, transconjugant; TET, tetracycline; TIO, ceftiofur.
The major subtype for the bla CTX-M gene was bla CTX-M-55 that occurred in 13 isolates and the remaining 2 isolates were bla CTX-M-101 and bla CTX-M-153, respectively (Table 1). Of note, bla CTX-M-153 found in Salmonella has not been previously reported.
PFGE and MLST analysis
An overall similarity of 78.96% was identified in these 15 isolates. According to 97% similarity, 13 PFGE patterns (A–M) were identified from 15 cephalosporin-resistant isolates in our study; 80% (12/15) isolates reached 85% pattern similarity. In particular, SJTUF14365 (Shanghai, 2016), SJTUF14367 (Shanghai, 2016), and SJTUF14903 (Shanxi, 2017) shared the PFGE pattern “A,” although they were recovered from different districts and years (Fig. 1). These three isolates might be from single clone showing similar resistance profile. The resistance profiles of SJTUF14365, SJTUF14367, and SJTUF14903 were NAL-SXT-AMP-STR-KAN-TET-CHL, NAL-SXT-AMP-KAN, and NAL-SXT-AMP-KAN, respectively. The MLST results indicated that all 15 isolates were classified as ST11 (Table 1). These findings indicated that cephalosporin resistance in Salmonella Enteritidis might persist for many years and probably resulted from clonal spread.
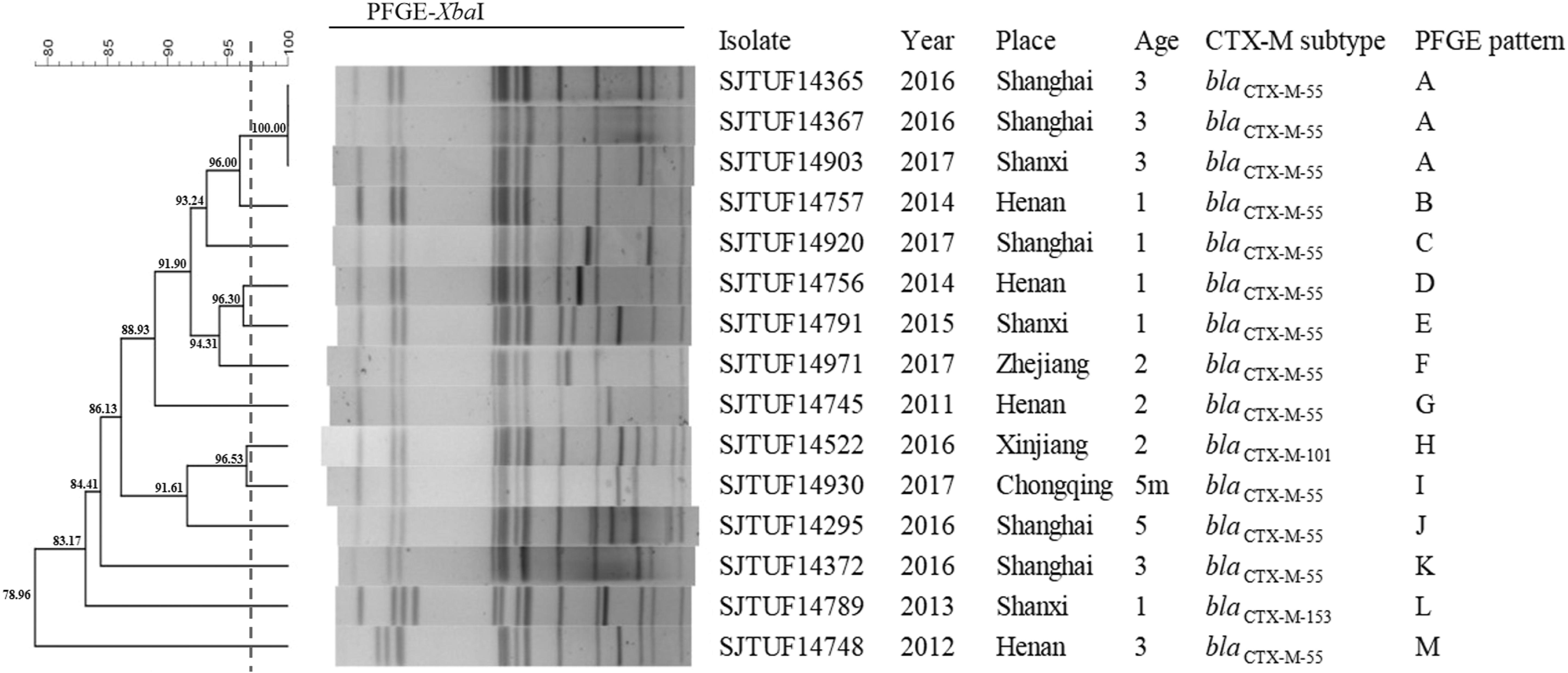
PFGE XbaI patterns of 15 cephalosporin-resistant Salmonella Enteritidis isolates. The dashed vertical line stood for the critical value (97%) to distinguish PFGE Pattern A and Patterns B-M. PFGE, pulsed-field gel electrophoresis.
Transferability analysis of bla CTX-M gene and plasmid characterization
Transfer of bla CTX-M by conjugation was successfully obtained from 13 isolates but was unsuccessful for the isolates SJTUF14756 and SJTUF14791. The 13 transconjugants exhibited at least 2, 64, 64, 128, 128, 256, and 256-fold increases in their MICs for cefoxitin, cefalotin, ceftiofur, cefepime, ceftazidime, ceftriaxone, and cefotaxime, respectively, compared with E. coli C600 (Table 1). These results demonstrated that the cefotaxime-resistance phenotype was accompanied by transfer of bla CTX-M-55, bla CTX-M-101, and bla CTX-M-153 to the transconjugants and that these three genes are necessary for cefotaxime resistance.
It was also found that the replicon types of the transferred plasmids included IncFIIs (n = 5), IncI1 (n = 5), IncN (n = 2), and IncFrepB (n = 8). The combination types IncFIIs-FrepB (n = 4), IncN-FrepB (n = 1), and IncN-FIIs-FrepB (n = 1) were identified in 6 transconjugants (Table 1). The sizes of the IncFrepB, IncN-FrepB, and IncN-FIIs-FrepB plasmids were ∼86.9, ∼95.7, and ∼173.4 kb, respectively. In addition, the sizes of IncI1 (n = 5) plasmids were determined as ∼86.9 kb (n = 4) and ∼104.5 kb (n = 1), whereas the IncFIIs-FrepB sizes were ∼78.2 kb (n = 1) and ∼138.9 kb (n = 1), respectively. Two plasmid types (IncFIIs and IncFrepB) were present in two transconjugants (SJTUF14365-TC [transconjugant] and SJTUF14903-TC) simultaneously. The bla CTX-M-101 and bla CTX-M-153 genes were located on ∼86.9 kb IncI1 and ∼104.5 kb IncI1 plasmids, respectively (Fig. 2).
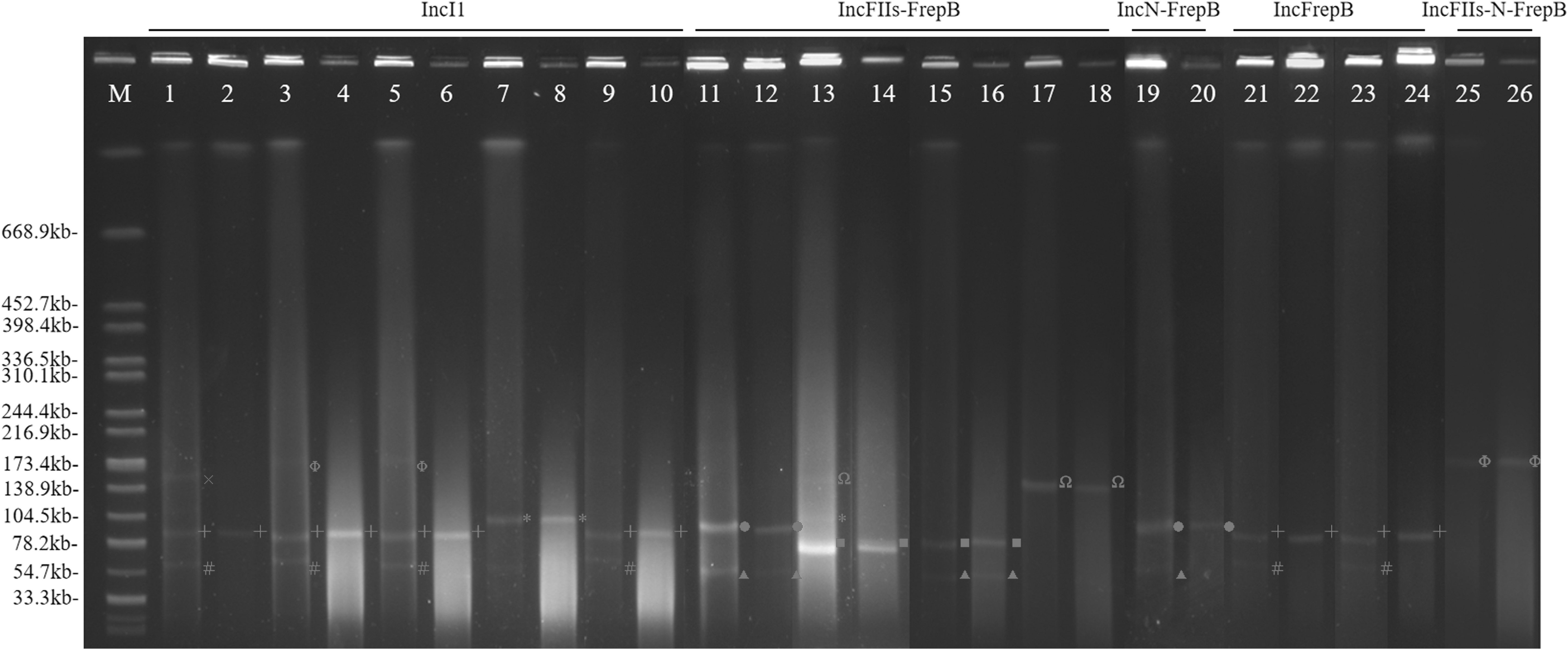
Plasmid profiles of Salmonella Enteritidis isolates and transconjugants determined by S1-PFGE. Lane M, H9812 molecular size marker; Lanes 1–26 correspond to isolates as follows: 1: 14295; 2: 14295-TC; 3: 14522; 4: 14522-TC; 5: 14748; 6: 14748-TC; 7: 14789; 8: 14789-TC; 9: 14930; 10: 14930-TC; 11: 14365; 12: 14365-TC; 13: 14757; 14: 14757-TC; 15: 14903; 16: 14903-TC; 17: 14971; 18: 14971-TC; 19: 14745; 20: 14745-TC; 21: 14367; 22: 14367-TC; 23: 14372; 24: 14372-TC; 25: 14920; 26: 14920-TC. Plasmid sizes are denoted as follows: ▲ ∼54.7 kb; # ∼62.5 kb; ■ ∼78.2 kb; + ∼86.9 kb; ● ∼95.7 kb; * ∼104.5 kb; Ω ∼138.9 kb; Χ ∼156.2 kb; Φ∼173.4 kb.
Genetic structure surrounding of bla CTX-M gene
To further understand the genetic structure adjacent to the bla CTX-M gene, PCR mapping was performed on the 13 transconjugants and the corresponding parental strains. The ISEcp1 elements upstream and orf477 downstream were the most prevalent insertions that flanked bla CTX-M-55. A complete ISEcp1 sequence was identified in four transconjugants and an incomplete ISEcp1 was identified in nine transconjugants (Fig. 3a). Hence, ΔISEcp1-bla CTX-M-55-orf477 (n = 8) was the most common, followed by ISEcp1-bla CTX-M-55-orf477 (n = 3). In addition, ISEcp1 was identified as the upstream of bla CTX-M-101 and bla CTX-M-153 genes (Fig. 3b, c). To our knowledge, this is the first report on the genetic structure surrounding bla CTX-M-153 (ΔISEcp1-ΔISSen9-ΔISEcp1-bla CTX-M-153-orf477).
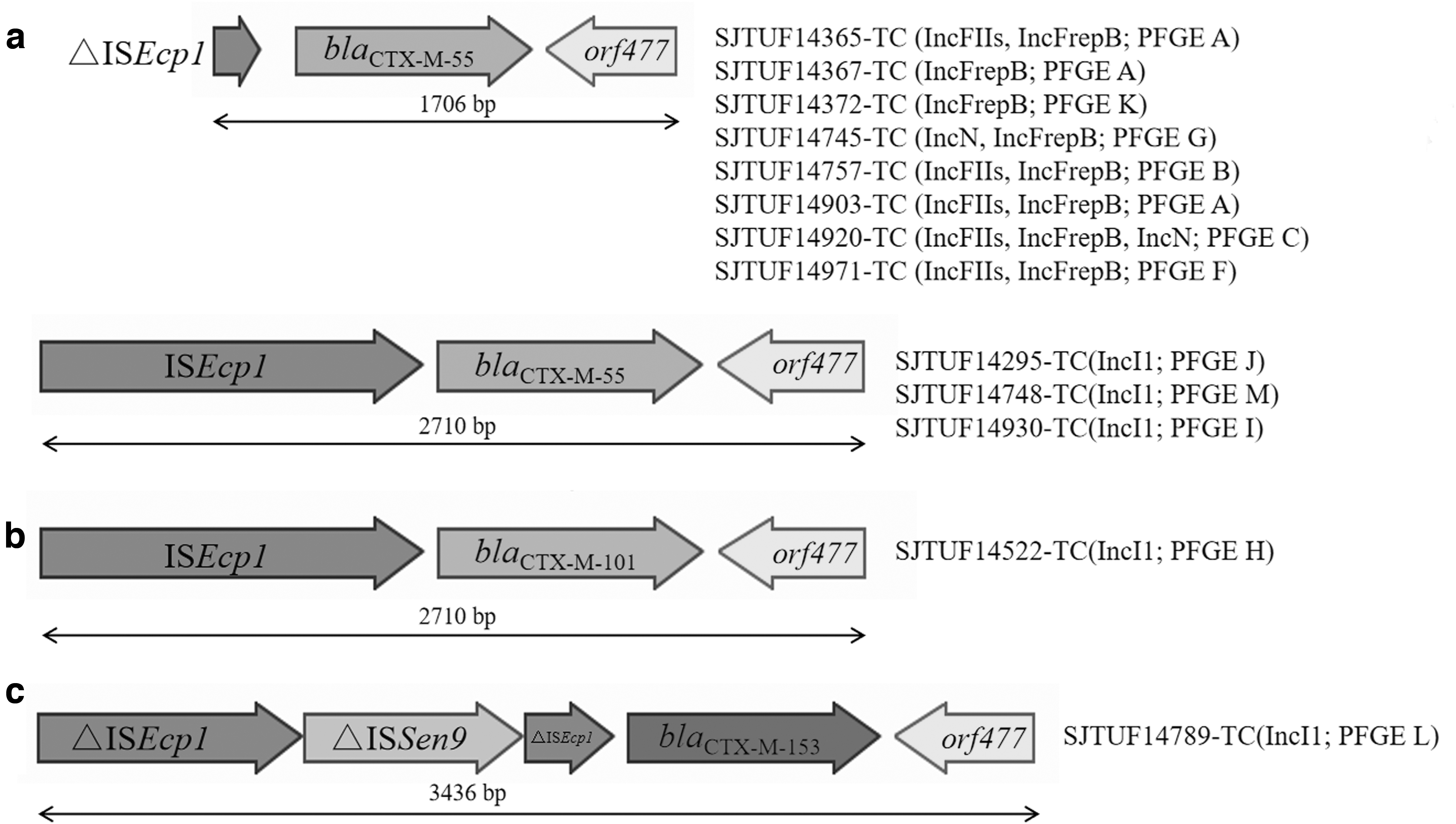
Genetic structure surrounding of bla
CTX-M in transconjugants.
Discussion
Salmonella Enteritidis has become one of the most prevalent foodborne pathogens leading to human and animal infections worldwide over the past two decades (Retamal et al., 2015). In particular, Salmonella infections in children occur at high rates (Jones et al., 2006; Li et al., 2014). Cephalosporins, which are the primary class of antimicrobials for the treatment of diarrhea and pneumonia in children, increased 399% from 2000 to 2015 (Li et al., 2014; Klein et al., 2018; Mi et al., 2018). However, with its widespread clinical use, bacteria develop resistance mainly by producing beta-lactamase, which hydrolyzes the beta-lactam ring to inactivate the antibiotics (Bush, 2018). The rate of resistance to cephalosporins was 3.8% in Japan (Shigemura et al., 2020) and 24.6% in Korea (Jeon et al., 2019). In our study, the rate of resistance to the third-generation cephalosporins was 9.26% in 162 Salmonella Enteritidis isolates from children younger than 5 years in China. Furthermore, 13 of 15 of our Salmonella Enteritidis isolates showed high-level resistance to ceftriaxone (MIC ≥128 μg/mL) and cefotaxime (MIC ≥128 μg/mL). In addition to cephalosporin resistance, some isolates (SJTUF14365, SJTUF14920, and SJTUF14971) were also resistant to other antimicrobial classes including both aminoglycosides (streptomycin and kanamycin), tetracycline (tetracycline), and phenicols (chloramphenicol). These results indicated that efforts should be made to ensure reasonable antimicrobial usage in the clinic for the treatment of salmonellosis infections in children and in food animal production industry.
In our study, 15 cephalosporin-resistant Salmonella Enteritidis isolates from various areas in China showed highly similar PFGE patterns suggesting the possibility of interspecies clonal spread. In particular, the single PFGE pattern “A” was shared by three cephalosporin-resistant isolates SJTUF14365 (Shanghai, 2016), SJTUF14367 (Shanghai, 2016), and SJTUF14903 (Shanxi, 2017), although these isolates were collected from different districts and in different years. In addition, all 15 of our isolates belonged to the single sequence type ST11, further suggesting that these isolates might be derived from the same ancestor and be locally persistent. ST11 is the most common genotype associated with the Salmonella Enteritidis isolates from human and food animals across the world (Chao et al., 2017; Yang et al., 2019). The combined MLST and PFGE results indicated that these 15 isolates were most likely clonally transmitted and this might lead to further salmonellosis outbreaks.
Resistance to ceftriaxone in Salmonella is usually owing to the presence of CTX-M group of extended-spectrum beta-lactams (ESBL) (Rodríguez et al., 2009). The bla CTX-M gene was first reported in 1990 and has become the predominant ESBL gene (Bauernfeind et al., 1990; Bonnet, 2004). The emergence of bla CTX-M in cephalosporin-resistant bacteria has resulted in more seriously clinical and public health problems (Qu et al., 2014; Zhang et al., 2016, 2019). In recent years, bla CTX-M-14 and bla CTX-M-15 were previously the most common genotypes among human and animal isolates in China (Li et al., 2011; Wu et al., 2012; Zhuo et al., 2013). The bla CTX-M-15 genotype was also the most prevalent bla CTX-M subtype in Enterobacteriaceae isolates from children (Dolejska et al., 2012; Moremi et al., 2017). Of interest, bla CTX-M-55 differs from bla CTX-M-15 by a single amino acid change (A77V) and has exceeded bla CTX-M-15 as the second most frequently identified CTX-M subtype in human and animal isolates in China (Kiratisin et al., 2007; Xia et al., 2014). Therefore, studies concerning dissemination and transferability of bla CTX-M-55 in clinical practices have rapidly increased in China (Zhang et al., 2019; Fu et al., 2020). In contrast, there are limited studies of bla CTX-M-55 in Salmonella isolates from children. In our study, bla CTX-M-55 was the predominant subtype in Salmonella Enteritidis isolates from children younger than 5 years across several provinces in China. This indicated that bla CTX-M-55 might still be dominating subtype in China and remains a threat to human health. Thus, it is necessary to study the existence and transfer of Salmonella Enteritis carrying bla CTX-M-55 collected from children.
Conjugative plasmids facilitate the horizontal transfer of bla CTX-M gene to other isolates and even to cross species barriers (Hu et al., 2018). Several plasmid families including IncF (Wang et al., 2018), IncI1 (Xia et al., 2017), and IncN (Fu et al., 2020) have been identified as major vehicles for bla CTX-M-55 transmission among Enterobacteriaceae. In our study, the bla CTX-M gene located on conjugative plasmids could transfer to the recipient E. coli C600, and plasmid families included IncI1, IncFIIs, IncN, and IncFrepB. These results suggested that conjugative plasmids played an important role in the horizontal transfer of bla CTX-M-55 in cephalosporin-resistant Salmonella Enteritidis. More seriously, transconjugants exhibited resistance to cephalosporins including cefalotin (MIC ≥512 μg/mL), ceftazidime (MIC ≥16 μg/mL), ceftriaxone (MIC ≥64 μg/mL), ceftiofur (MIC ≥32 μg/mL), and cefotaxime (MIC ≥32 μg/mL). Transconjugant SJTUF14789-TC carrying bla CTX-M-153 showed higher resistance to cephalosporin than other transconjugants. These findings indicated that bla CTX-M--positive plasmids were associated with cephalosporin resistance in Salmonella Enteritidis, and more attention should be paid to bla CTX-M-153 in Salmonella Enteritidis.
An important feature of the bla CTX-M plasmids is the frequent existence of insertion fragments such as ISEcp1, IS26, and IS903 (Lartigue et al., 2004). Insertion sequences, such as ISEcp1, are a heterogeneous class of mobile elements that can promote the translocation of antimicrobial resistance genes (Chen et al., 2006; Miriagou et al., 2006; Sun et al., 2016). In our study, ISEcp1 was found upstream of bla CTX-M-55 gene as both complete and incomplete copies implying a role for this IS in bla CTX-M-55 transposition. Incomplete ISEcp1 in the upstream of bla CTX-M-153 gene was interrupted by incomplete ISSen9, which may influence the transposition of genes. Moreover, ISEcp1 is a strong activator of bla CTX-M-55 expression (Wang et al., 2013). This implies that the high-level MICs exhibited by our transconjugants and parent isolates might be the result of increased gene expression of bla CTX-M-55 mediated by ISEcp1.
Conclusions
Cephalosporin resistance in Salmonella Enteritidis isolates from children was identified in our study and was the result of the predominant genotype bla CTX-M-55 among the CTX-M group of ESBLs. The conjugative transfer of bla CTX-M-55 might contribute to the spread of cephalosporin resistance in Salmonella Enteritidis. Therefore, it is urgently needed to monitor the prevalence of cephalosporin-resistant Salmonella Enteritidis carrying bla CTX-M in clinical treatment settings to better understand and control its threat to global public health.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Key R&D Program of China (Grant No. 2017YFC1600100).