
Abstract
Bacillus cereus is a common foodborne pathogen that can cause both gastrointestinal and nongastrointestinal diseases. In this study, we collected 603 meat and meat products from 39 major cities in China. The positive contamination rate of B. cereus in the collected samples was 26.37% (159/603), and the contamination level in 5.03% (8/159) positive samples exceeded 1100 most probable number/g. The detection rates of virulence genes were 89.7% for the nheABC gene group, 37.1% for the hblACD gene cluster, 82.3% for cytK-2, and 2.9% for cesB. Notably, all isolates presented with multiple antibiotic resistance, and 99.43% of isolates were resistant to five classes of antibiotics. In addition, the multilocus sequence typing results indicated that all isolates were rich in genetic diversity. Collectively, we conducted a systematic investigation on the prevalence and characterization of B. cereus in meat and meat products in China, providing crucial information for assessing the risk of B. cereus occurrence in meat and meat products.
Introduction
The Bacillus cereus group, also known as B. cereus sensu lato, includes eight closely related species: B. anthracis, B. cereus sensu stricto, B. cytotoxicus, B. mycoides, B. pseudomycoides, B. thuringiensis, B. toyonensis, and B. weihenstephanensis (Helgason et al., 2000; Guinebretière et al., 2013; Pfrunder et al., 2016). Members of the B. cereus group have a major impact on public health, agriculture, and the food industry. For example, B. anthracis is the etiological agent of anthrax. Virulent forms of B. anthracis harbor two plasmids, pXO1 and pXO2 (Okinaka et al., 1999). Bacillus thuringiensis has several plasmids encoding various insecticidal toxins, such as Cry and Cyt, which constitute the parasporal crystals of this species, and have been commercialized as biological pesticides (Bravo et al., 2011). Bacillus cereus, belonging to B. cereus sensu stricto, is a flagellated bacterium that can form stress-resistant spores, thereby promoting its widespread presence in a variety of environments (Marrollo, 2016; Griffiths and Schraft, 2017; Gao et al., 2018).
Bacillus cereus can cause both gastrointestinal and nongastrointestinal infections, where the former includes vomiting and diarrhea (Stenfors Arnesen et al., 2008; Senesi and Ghelardi, 2010; Ceuppens et al., 2012; Ehling-Schulz et al., 2015). Two types of toxins secreted by B. cereus account for the occurrence of gastrointestinal diseases. Enterotoxins, which are heat-labile proteins, cause diarrheal symptoms and mainly include hemolysin BL (HBL), nonhemolytic enterotoxin (NHE), and cytotoxin K (CytK) (Madegowda et al., 2008; Stenfors Arnesen et al., 2008; Clair et al., 2010). Unlike enterotoxins, vomiting toxin cereulide is a cyclic polypeptide synthesized by the nonribosomal peptide synthetase ces gene cluster (Ehling-Schulz et al., 2005), which is a highly thermo- and pH-resistant molecule (Ehling-Schulz et al., 2015). Hence, the inactivation of preformed cereulide by food processing steps is unlikely to occur (Paananen et al., 2002). Cereulide is considered a highly toxic substance, and induces symptoms, such as vomiting and nausea 0.5–6 h after ingesting contaminated food (Andersson et al., 2007; Rouzeau-Szynalski et al., 2020). Therefore, the contamination of B. cereus, especially that of the emetic strain in food, poses a great hazard to human health. Nongastrointestinal diseases caused by B. cereus, such as fulminant bacteremia, meningitis, endophthalmitis, pneumonia, and gas gangrene-like skin infections, are occasionally severe and fatal (Bottone, 2010; Rishi et al., 2013; Ikeda et al., 2015).
Several types of food are often susceptible to contamination by pathogenic B. cereus, resulting in food poisoning (Glasset et al., 2016; Nicholls et al., 2016; Schmid et al., 2016; Lentz et al., 2018; Thirkell et al., 2019). As meat and meat products are essential to daily life, a variety of studies have been conducted on the contamination of B. cereus in meat and meat products in different countries (Eglezos et al., 2010; Tewari et al., 2015; Shawish and Tarabees, 2017). Although meat and meat products are usually stored at low temperatures, there is still a risk of food poisoning once they are contaminated with toxin-producing psychrophilic B. cereus (Fermanian et al., 1997). In addition, inappropriate storage temperatures during transport or sale could promote the propagation of B. cereus, which can create a potential risk to consumers.
To date, there are no reports on the overall risk assessment of B. cereus in meat and meat products in China. We, therefore, analyzed the occurrence and contamination level of B. cereus in meat and meat products based on isolates collected from 39 major cities in China from 2011 to 2016 and characterized the potential hazards of these isolates. Our study provides a crucial reference for assessing the risk of B. cereus in meat and meat products in China.
Materials and Methods
Sample collection
Sampling was conducted in accordance with the guidelines of the Chinese Food Safety Standards (The Hygiene Ministry of China, 2010). From 2011 to 2016, 39 cities, including Hong Kong, Macao, and all the capital cities of all provinces in China, were selected (Supplementary Fig. S1). A total of 603 samples of meat and meat products were randomly collected (Table 1), including 541 samples of raw meat and 62 meat products (semifinished products without cooking). After collection, all samples were immediately placed in sterile and self-sealed plastic bags, stored at 4°C, and transported to the laboratory for further analysis.
Contamination Rates and Most Probable Number Values of Bacillus cereus Isolated from Meat and Meat Products in China
Contamination rate = number of positive samples/total samples.
MPN value (MPN/g) = MPN of B. cereus per gram sample.
MPN, most probable number.
Isolation and identification of B. cereus
Qualitative and quantitative detection of B. cereus (total number of vegetative cells and spores) were performed according to the Chinese Food Safety Standards and previous publications (The Hygiene Ministry of China, 2003, 2014; Tallent et al., 2012) with minor modifications. Twenty-five-gram samples were cut into strips, transferred to a sterile homogenizer containing 225 mL of phosphate buffered saline (0.01 mol/L) and homogenized for 2 min at 8000–10,000 rpm. The homogenized solution and its 10 × and 100 × dilutions were used for qualitative and quantitative detection of B. cereus. The homogenized solution and dilutions were incubated at 30°C for 48 h. Bacillus cereus was isolated and initially identified using mannitol-egg-polymyxin agar plates (selective medium; Huankai) and chromogenic B. cereus agar plates (Huankai).
Biochemical identification was carried out in accordance with the Chinese Food Safety Standards (The Hygiene Ministry of China, 2003). The quantitative test was performed using the most probable number (MPN) counting method, which was determined based on the number of positive tubes and the MPN table in each of the three groups (Tallent et al., 2012; The Hygiene Ministry of China, 2014). Enterobacterial repetitive intergenic consensus (ERIC)-PCR was employed to distinguish clonal strains as described previously (Versalovic et al., 1991; Yu et al., 2020).
Virulence gene detection
PCR was used to detect the presence of seven enterotoxin-encoding genes (hblA, hblC, hblD, nheA, nheB, nheC, and cytK-2) and a cereulide synthase gene (cesB). The PCR conditions were set and the primer sequences and amplification procedures were performed as previously described (Hansen and Hendriksen, 2001; Ehling-Schulz et al., 2005; Oltuszak-Walczak and Walczak, 2013) (Supplementary Table S1). Bacillus cereus ATCC 14579, B. cereus NC7401, and B. cereus F4810/72 were used as reference strains.
Antibiotic susceptibility test
The antibiotic susceptibility of different isolates to 20 antibiotics was tested using the Kirby–Bauer paper plate diffusion method as described previously (The Clinical and Laboratory Standards Institute [CLSI], 2010; Yim et al., 2015; Osman et al., 2018; Park et al., 2018, 2020; Fiedler et al., 2019; Gdoura-Ben Amor et al., 2019; Yu et al., 2019; Zhao et al., 2020). The isolates were classified as susceptible (S), intermediate (I), or resistant (R) according to CLSI guidelines, and the zone of inhibition was measured and interpreted according to the regional diameter standard of Staphylococcus aureus in the CLSI reference (CLSI, 2010) (Supplementary Table S2). The multiple antibiotic resistance (MAR) index of a single isolate was evaluated according to a previous report (Krumperman, 1983).
Multilocus sequence typing and sequence analysis
Seven housekeeping genes (glp, gmk, ilvD, pta, pur, pycA, and tpi; PubMLST
Data analysis
The chi-square test was applied to determine the cumulative difference between the raw meat and meat products using statistical analysis. A p-value <0.05 was considered statistically significant for all the parameters evaluated.
Ethics statement
This study did not involve any animal experiments.
Results
Occurrence and contamination level of B. cereus in meat and meat products
Among the 603 meat and meat products, B. cereus was identified in 159 (26.37%) samples (Table 1), including 22.58% (28/124) from beef, 26.73% (27/101) from chicken, 41.38% (24/58) from ducks, 22.73% (5/22) from lamb, and 25.17% (75/298) from pork. Positive samples were found in all cities except Yinchuan. Moreover, the occurrence of B. cereus was much higher in the processed meat products than in the raw meat products (41.94% vs. 24.58%, p = 0.003) (Supplementary Table S3). Based on quantitative analysis, 67.93% (108/159) of the positive samples were contaminated at levels ranging between 3 and 1100 MPN/g, which were 42.31% (11/26) and 72.93% (97/133) in meat products and raw meat products, respectively (Supplementary Table S3). Notably, 5.03% (8/159) of the positive samples had a contamination level of >1100 MPN/g, which were detected in positive samples of duck (2/24, 8.33%), chicken (2/27, 7.41%), and pork (4/75, 5.33%). As a result of isolation, biochemical characterization, and ERIC-PCR screening, 175 strains were obtained (Supplementary Fig. S1).
Distribution of virulence genes in B. cereus isolates
The distribution of virulence genes is shown in Figure 1. The detection rates of nheA, nheB, and nheC were 99.4%, 98.3%, and 91.4%, respectively, whereas the nheABC gene cluster was found in 89.7% of the population. The hbl gene cluster was only present in some of the strains (37.1%), with hblA, hblC, and hblD in 42.9%, 45.7%, and 53.7% of all isolates, respectively. Furthermore, 5.7% (10/175) were positive for two genes, whereas 38.9% (68/175) of the isolates had no hbl genes (Supplementary Table S4). In this study, at least one gene of each of NHE and HBL complexes was detected in 100% and 61.1% of the strains, respectively. Among the isolates from raw meat, this percentage ranged from 27.6% to 48.3% for hblACD genes, and from 80.0% to 96.6% for the nheABC gene cluster (Supplementary Table S4).
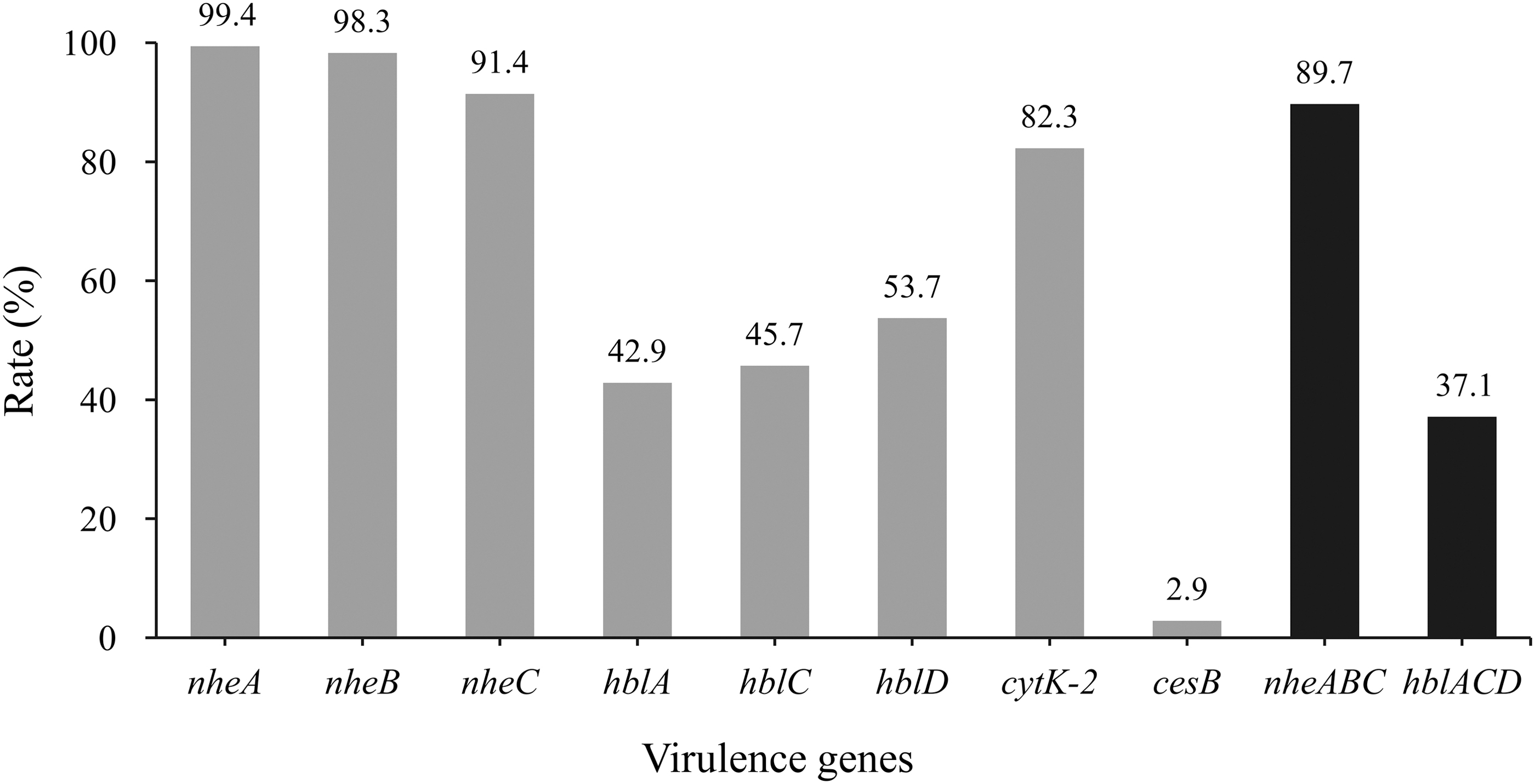
Distribution of virulence genes in Bacillus cereus isolated from meat and meat products in China. The number at the top of the bars represents the positive rate of the corresponding toxin genes. hblACD and nheABC mean that the strains are positive for hblA, hblC, and hblD, or nheA, nheB, and nheC at the same time, respectively. All eight genes are present in the strains with toxin genes.
In the strains obtained from meat products, the hblACD and nheABC gene clusters were present in 32.1% (9/28) and 82.1% (23/28) of all isolates, respectively (Supplementary Table S4). The harboring rate for cytK-2 was also remarkably high, accounting for 82.3% of all strains, including 92.9% (26/28) of meat products and 80.3% (118/147) from raw meat (Supplementary Table S4). In contrast, only 2.9% (5/175) of the strains harbored the vomiting toxin synthase gene cesB. The cesB gene was only detected in isolates from bacon (6.3%), beef (3.5%), chicken (3.5%), duck (3.5%), and pork (1.8%), but not in isolates from dachshund, cured duck leg, meatball, sauce beef, and mutton (Supplementary Table S4).
Antibiotic susceptibility test of all isolates
In terms of antibiotic susceptibility, all of the strains exhibited resistance to the penicillins (ampicillin and penicillin; 100.00%), and most of them were resistant to β-lactam/β-lactamase inhibitor combinations (amoxicillin; 97.14%), ansamycins (rifampicin; 86.29%), and cephems (parenteral), especially cephalothin (75.54%), and cefoxitin (94.86%) (Fig. 2). In contrast, most of the strains were sensitive to cefotetan, imipenem, gentamicin, kanamycin, telithromycin, teicoplanin, ciprofloxacin, chloramphenicol, tetracycline, and trimethoprim-sulfamethoxazole. Notably, according to the definition of multidrug resistance (Magiorakos et al., 2012), all strains were resistant to at least three classes of antibiotics, and 99.43% of isolates exhibited resistance to five or more classes of antibiotics.
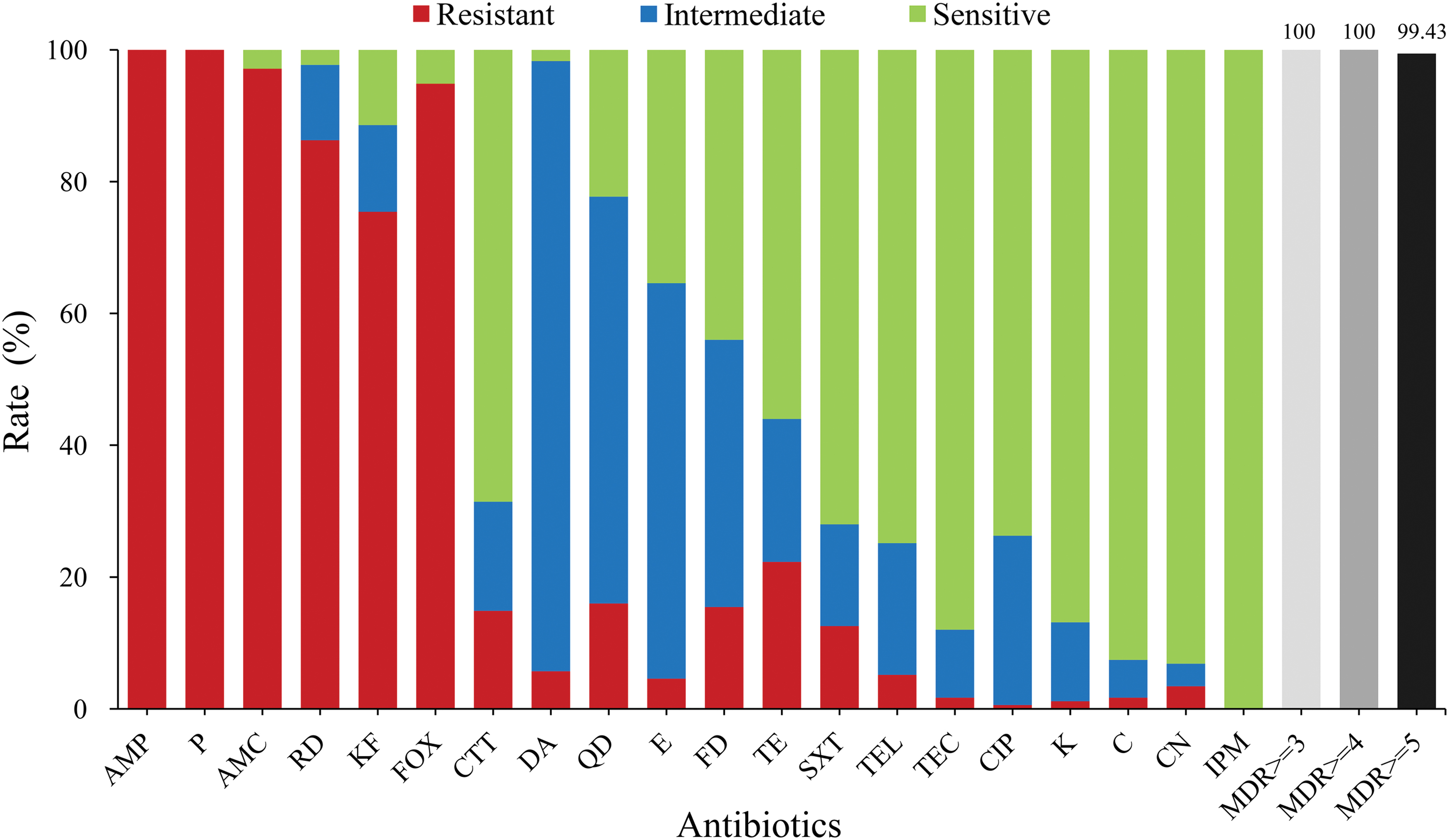
Antibiotic susceptibility of Bacillus cereus isolated from meat and meat products in China. The red, blue, and green bars represent the proportion of resistant, moderately resistant, and sensitive strains, respectively. The light gray, gray, or dark bars represent the proportion of strains with multiple drug resistance. Color images are available online.
Overall, MAR indices ranged from 0.15 to 0.50. We found that 46.86% (82/175) isolates had MAR indices ≥0.25 and 21.71% (38/175) had MAR indices >0.3. A single strain (named 1238-2-Bc) had an MAR index of 0.5.
Genetic diversity analysis
According to the multilocus sequence typing (MLST) analysis, 175 strains were assigned to 112 ST types, including 47 new ST types (42%; 47/112) (Fig. 3). The main ST types were ST205 and ST770 (n = 14), followed by ST4 (n = 6), ST26 (n = 6), ST1066 (n = 5), ST90 (n = 5), and ST177 (n = 5). Moreover, several of the strains were grouped into six defined clonal complexes (CCs), including ST-205, ST-142, ST-18, ST-97, ST-111, and ST-365 (Fig. 3 and Supplementary Fig. S2). The largest CC (ST-205) contained 20 STs (n = 40 strains), but singletons had the largest population, containing 69 STs (n = 103 strains).

Minimum spanning tree and genetic diversity of Bacillus cereus isolated from meat and meat products in China. Circles of assorted colors represent different clonal complexes and monomers. The numbers in the circle indicate the multilocus sequence typing type, and the size of the circle represents the proportion and number of strains. Color images are available online.
Phylogenetic analysis of 175 isolates and five type strains (B. cereus ATCC 14579, B. anthracis ATCC 4728, B. thuringiensis ATCC 10792, and two clinical emetic-type strains B. cereus NC7401 and B. cereus F4810/72) were performed based on the MLST alleles. All isolates and type strains were categorized into 14 clusters when the similarity threshold was set to 50% (Supplementary Fig. S2). The fourth cluster contained the largest population of the isolates (n = 47), which also contained the reference strains F4810/72 and NC7401. It is worth noting that all isolates with the cesB gene were also clustered in the fourth cluster. Only one strain, namely 1238-2-Bc, was grouped into the 13th cluster; however, this strain showed the most abundant antibiotic-resistant profile, which was found to resist 15 classes of antibiotics.
Discussion
Meat is one of the main sources of nutrients for humans due to its high protein content and micronutrient richness. Meat and meat products may be contaminated by pathogens during processing and preparation. In addition, storing meat products at 25°C and above for several hours will increase the ability of micro-organisms to proliferate, thereby increasing the risk of foodborne diseases. Hence, contamination of meat and meat products by foodborne pathogens is still a major public health concern. Bacillus cereus is a common foodborne pathogen, which is ubiquitous in the environment and commonly presents in a variety of food; however, the contamination of B. cereus in meat and meat products in China has rarely been reported. In this study, 603 samples of meat and meat products were collected from 39 major cities in China, and B. cereus-positive samples were distributed in 38 cities except for Yinchuan, indicating a wide range of B. cereus contamination. The contamination level was similar to that reported in Uttarakhand, India, and Egypt (Tewari et al., 2015; Shawish and Tarabees, 2017), and was higher than that in Australia (Eglezos et al., 2010).
Although B. cereus was detected in all types of meat and meat products, its presence in duck meat was much higher than that in other kinds of meat and meat products (Table 1). The overall finding is consistent with that of previous reports (Rather et al., 2012 Shawish and Tarabees, 2017), which may be due to ambient temperature, food storage, and/or improper cooking before food consumption. These mentioned conditions promote the germination of endospores and lead to a rapid increase in the total number of B. cereus (Rather et al., 2012; Shawish and Tarabees, 2017). In addition, during the processing of meat products, additives, condiments, and spices may be added, which are considered to be potential risk factors and can increase the number of Bacillus spores, thereby increasing the incidence of B. cereus and food poisoning (Tewari et al., 2015; Shawish and Tarabees, 2017; Berthold-Pluta et al., 2019). Although the incidence of B. cereus in meat products was higher than that in raw meat, all the samples with MPN exceeding 1100 MPN/g belonged to raw meat.
The well-studied diarrheal toxins secreted by B. cereus are encoded by nheABC, hblACD, and cytK-2 (Jeßberger et al., 2015). Among the different enterotoxic genes, the presence of the nhe cluster was very high with 89.7% of the isolates harboring this gene cluster, which is similar to the findings of previous studies on B. cereus contamination in other types of food (Gao et al., 2018; Berthold-Pluta et al., 2019; Fiedler et al., 2019; Gdoura-Ben Amor et al., 2019; Yu et al., 2019, 2020). Of the isolates, 82.3% possessed the enterotoxin gene cytK-2, which is similar to that observed in previous studies (Chon et al., 2015). In contrast, only 2.9% of all strains (five strains) harbored the emetic toxin encoding gene cesB, which was lower than that reported in other studies (Biesta-Peters et al., 2016; Owusu-Kwarteng et al., 2017). Nevertheless, both diarrheal and emetic strains were found in meat and meat products in China, suggesting the potential pathogenicity of these isolates.
Currently, antibiotics remain as one of the major approaches for the treatment of serious infections caused by different pathogens. However, various pathogens have developed antibiotic resistance that leads to the failure of clinical treatments. Therefore, characterization of antibiotic susceptibility can provide a crucial reference for clinical treatment. β-lactam antibiotics are the first-line treatment for clinical infection. For example, ampicillin is used in the treatment of septicemias and respiratory and urinary tract infections, and is widely used in animal production (Kadri et al., 2018; Gdoura-Ben Amor et al., 2019). However, most of the β-lactam antibiotics, as well as rifampin, were not effective against most of the strains and should not be clinically applied for the treatment of B. cereus. Likewise, some strains were resistant to gentamicin and ciprofloxacin, which have been categorized as critically important drugs for human medicine by the World Health Organization (WHO 2019). Furthermore, at least 46% of isolates had MAR indices >0.25, which is considered to originate from high-risk sources of contamination. These high-risk sources of contamination may be caused by the abuse of antibiotics in animal breeding, and the abuse of antibiotics may cause the spread of drug-resistant pathogens in both livestock and humans, posing a significant public health threat (Ibrahim et al., 2020; Van Boeckel et al., 2015). Taken together, antibiotic resistance exhibited by B. cereus in meat and meat products must be given attention.
Of the isolates studied, 175 strains were assigned to 112 ST types, indicating the overall diversity of B. cereus in meat and meat products. From the clustering results in the evolutionary analysis (Supplementary Fig. S2), 1238-2-Bc and B. anthracis ATCC 4728 were grouped into different single clusters. This suggests that the phylogenetic relationship between them and other strains is remote and may have evolved from different origins. Different virulence gene profiles and antibiotic resistance profiles were randomly distributed across different branches, suggesting that isolates may have evolved from different origins. In addition, some STs (ST4, ST26, ST78, and ST92) assigned to the strains have been reported to cause gastrointestinal disease outbreaks, indicating that these isolates have potential hazards (Hoffmaster et al., 2008). At the same time, clinically pathogenic strains and all isolates with cesB were clustered in the fourth cluster, demonstrating that the evolutionary relationships among cesB-positive strains are much closer and the cesB-positive strains identified in this study may have a higher potential for pathogenicity.
Conclusion
Bacillus cereus can be easily identified in meat and meat products in China, with an overall contamination rate of 26.37%. Notably, all isolated strains exhibited resistance to multiple antibiotics, indicating that the development of antibiotic resistance within B. cereus is severe. Some isolates have the same STs as the reported foodborne B. cereus that caused a gastrointestinal disease outbreak, indicating that these isolates may pose a potential risk to cause gastrointestinal diseases. Therefore, it is necessary to continuously monitor B. cereus contamination in meat and meat products in China.
Footnotes
Authors' Contributions
Y.D., Q.W., J.Z., and J.W. conceived the project and designed the experiments. L.K., S.Y., X.Y., C.L., P.Y., H.G., S.W., Q.Y., T.L., X.Y., Y.Z., X.W., and H.Z. performed the experiments. Y.D., and Q.W. supervised the project. L.K., S.Y., and Y.D. analyzed the data and wrote the article. Y.D., Q.W., J.Z., and J.W. revised the article. All authors have read and agreed to the published version of the article.
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the Guangdong Technological Innovation Strategy of Special Funds (Key Areas of Research and Development Program, grant no. 2018B020205003), National Natural Science Foundation of China (grant nos. 31730070 and 31701195), GDAS' Special Project of Science and Technology Development (grant no. 2019GDASYL-0201001), the 1000-Youth Elite Program, and New Zealand–China Scientist Exchange Programme (2019).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.