
Abstract
Proteus mirabilis is an opportunistic human pathogen. In this study, a novel SXT/R391 integrative and conjugative element (ICE), named ICEPmiChnS012, was identified in the multidrug-resistant P. mirabilis strain S012 that was isolated from retail chicken in China. Whole genome sequencing revealed that ICEPmiChnS012 carried 22 resistance genes including aac(6′)-Ib-cr, fosA3, bla OXA-1, bla CTX-M-65, and bla HMS-1. ICEPmiChnS012 harbored 10 copies of IS26 and IS26-mediated genetic new rearrangements caused variations in HS4 region. To our knowledge, an unusual gene cassette array dfrA1-ereA1-aadA2 was found in P. mirabilis in this study for the first time. And this is the first report of identification of aph3-VI and bla HMS-1 in VRIII region in P. mirabilis. The conjugation experiments proved that ICEPmiChnS012 could be transferred to Escherichia coli EC600 through conjugation. These findings demonstrated that ICEPmiChnS012 was a special ICE that carried the largest number of antimicrobial resistance genes in the family of SXT/R391 ICEs. This element could serve as an important vehicle for the dissemination of antibiotic resistance genes and should receive great concern.
Introduction
P
P. mirabilis is naturally resistant to several antibiotics, including colistin, nitrofurans, tigecycline, and tetracycline (Stock, 2003). In recent years, P. mirabilis strains resistant to other antibiotics are increasingly reported (Girlich et al., 2020). Integrative and conjugative elements (ICEs) are crucial for the acquisition of resistance genes. ICEs are self-transmissible mobile genetic elements that have the ability to integrate into their host's chromosome (Johnson and Grossman, 2015). Several types of ICEs have been described to date. The SXT/R391 family of ICEs is one of the largest families with the most abundant diversity among bacterial species (Carraro and Burrus, 2014). All the members of the SXT/R391 family integrate into chromosome at the 5′ end of the gene prfC, which encodes the peptide chain release factor 3 (RF-3). RF-3 is a GTP-binding protein that stimulates translational termination. SXT/R391 ICEs share similar genetic structures including five hot spots, five variable regions, and core genes involved in integration/excision, conjugation, and regulation of ICEs (Li et al., 2016).
Many SXT/R391-type ICEs have been identified in strains of P. mirabilis (Bie et al., 2017). Lei et al. (2018) isolated a clinical P. mirabilis strain from fecal swab of diseased pig with diarrhea in 2016 in Sichuan province of China. A novel SXT/R391 ICE, ICEPmiChnBCP11, harboring 22 different antibiotic resistance genes, was characterized by whole genome sequencing (Lei et al., 2018). As ICEs could be transmitted among Proteus, V. cholerae, and other Enterobacteriaceae and promote the acquisition of resistance genes, it is necessary to continuously monitor the antimicrobial resistance and related mobile elements among P. mirabilis isolates (Aberkane et al., 2016).
In this article, we reported a multidrug-resistant P. mirabilis strain isolated from retail chicken in China. A novel SXT/R391 ICE carrying 22 different resistant genes was described.
Materials and Methods
Strain isolation and antimicrobial susceptibility assay
P. mirabilis strain S012 was isolated from retail chicken in May, 2017, in Shennongjia Forestry District, Hubei Province, China. Antimicrobial susceptibility testing was performed using the Kirby–Bauer disk diffusion test with Mueller–Hinton agar according to instructions from the Clinical and Laboratory Standards Institute guidelines (CLSI, 2018). The antibiotic disks were purchased from Hangzhou Binhe Microorganism Reagent Co., Ltd, China. A total of 14 antibiotics including nalidixic acid, sulfisoxazole, ampicillin, streptomycin, tetracycline, chloramphenicol, rifampicin, and ceftazidime were evaluated. Escherichia Coli ATCC25922 was used as a quality control. After incubation, the inhibition zone diameters (in millimeters) around disks on Mueller–Hinton agar were measured and interpreted according to respective CLSI breakpoints.
Genome sequencing and assembly
The bacteria cells were directly sent to Shanghai Majorbio Bio-Pharm Technology Co., Ltd, for DNA extraction and genome sequencing. DNA was extracted using Wizard® Genomic DNA Purification Kit (Promega). Whole-genome sequencing was performed using the PacBio RS II and the Illumina HiSeq X-10 platforms (Majorbio, Inc., China). For Illumina sequencing, DNA samples were sheared into 400–500 bp fragments using a Covaris M220 Focused Acoustic Shearer and 400-bp DNA library was constructed from the sheared fragments using the NEXTflex™ Rapid DNA-Seq Kit. For PacBio sequencing, 10-kb DNA library was constructed and run in one single molecule real-time cell. Illumina and PacBio reads were assembled with Unicycler v0.4.7, which depended on SPAdes 3.13.0 and GapCloser 1.12 (Bankevich et al., 2012). The chromosome was assembled into one scaffold using HGAP3 and canu 2.0 (Koren et al., 2017). Final error correction of the PacBio assembly results was performed using the Illumina reads using Pilon 1.23 (Walker et al., 2014). The genome was annotated using Glimmer 3.02 and GeneMarkS4.3. Prophage annotation was done using PHAST and IslandPath-DIMOB 1.0.0 was used to predict genomic islands. Resistance genes were identified by Resfinder 4.1 (
Conjugation experiments
A conjugation experiment was performed by a broth mating method using the rifampicin-resistant E. coli EC600 recipient strain. Transconjugants were selected on MacConkey agar supplemented with rifampicin (200 μg/mL) and streptomycin (32 μg/mL). The transconjugants were further confirmed by the presence of resistance gene strB and susceptibility to streptomycin. Detection of strB was performed by primers strBF (5′-GCGGACACCTTTTCCAGCCT-3′) and strBR (5′-TCCGCCATCTGTGCAATGGCG-3′) (Hou et al., 2016). Polymerase chain reaction (PCR) amplification was performed as follows: 94°C for 2 min, followed by 30 cycles of 94°C for 30 s, 55°C for 30 s, and 72°C for 35 s, and a final extension at 72 C for 2 min. The location of ICE in transconjugant was determined by PCR amplification with primers LE1/LE4 and RE4/RE1 (McGrath et al., 2006). Primers LE1 (5′-GTTTCTTCGTTGCACGAACTGG-3′) and LE4 (5′-GTACAC ACTTTCCGAGGTTACG-3′) were used to amplify regions across the attL junction and should generate a PCR product of 347 bp. Primers RE1 (5′-CGGTCTGAATGGCCTGTCCGAA-3′) and RE4 (5′-CCGCAATACCCT GCAATACCGA-3′) were used to amplify regions across the attR junction and should result in a PCR product of 454 bp. (McGrath et al., 2006). PCR amplification was preceded by an initial denaturation step (95°C, 10 min), followed by 30 cycles of 95°C for 1 min, annealing temperature (Ta)-5°C for 45 s, 72°C for 1 min. Reactions were completed at 72°C for 8-min cycle (McGrath and Pembroke, 2004). The susceptibility to streptomycin for transconjugant was detected using the microbroth dilution method in accordance with the guidelines in CLSI (CLSI, 2018). Mueller–Hinton broth containing serial twofold dilutions of streptomycin was used and final concentrations of streptomycin that ranged from 1 to 1024 μg/mL were examined.
Results and Discussion
P. mirabilis strain S012 had multiple drug resistance profiles. It was resistant to ampicillin (10 μg), amoxicillin (10 μg), ceftriaxone (30 μg), streptomycin (10 μg), tetracycline (30 μg), nalidixic acid (30 μg), sulfisoxazole (300 μg), chloramphenicol (30 μg), gentamicin (10 μg), ciprofloxacin (5 μg), rifampicin (5 μg), and suldimethoxine (5 μg). It was sensitive to ceftazidime (30 μg) and showed intermediate resistance to cefotaxime (30 μg). Complete genome sequence of P. mirabilis strain S012 was obtained by combining Illumina and PacBio reads. Illumina sequencing generated 3,503,781 × 2 paired-end reads with 150 bp in each read. The average read length of the PacBio raw data set was 20,756 bp with a maximum read length of ∼187,511 bp. The genome was assembled and contained 4,072,318 bp, with a GC content of 39.11%. No plasmid sequence had been identified.
A total of 3745 genes including 30 different antimicrobial resistance-related genes were predicted. The results of antimicrobial susceptibility and predicted resistance genes are listed in Table 1. Several antibiotic resistance genes related to macrolide, fosfomycin, and bleomycin and disinfectant resistance genes related to quaternary ammonium compounds were found by Resfinder, whereas the corresponding antimicrobial and disinfectant agents were not checked in the study because of experimental design. The majority of resistance phenotypes showed agreement with the genotypes.
Antimicrobial Susceptibility Assay Results and Resistance Genes Detected in Proteus mirabilis S012
ICE, integrative and conjugative element; MBLs, metallo-β-lactamases; NA, not available.
Analysis of the genome of P. mirabilis S012 indicated that the isolate harbored an SXT/R931 type ICE that was designated ICEPmiChnS012. ICEPmiChnS012 was 147,957 bp in size with a GC content of 49.25% (corresponding to bases 3312546–3460502 in the chromosome). Structural analysis of ICEPmiChnS012 revealed that it contained five hotspot regions and five variable regions. BLASTn analysis showed that ICEPmiChnS012 shared 99.95% nucleotide identity within 84% query coverage with ICEPmiChnBCP11 (GenBank accession: MG773277.1). ICEPmiChnBCP11 was a novel SXT/R931 type ICE characterized in P. mirabilis of swine origin in China in 2016 (Lei et al., 2018).
SXT/R391 ICEs can generally excise from the chromosome, form the circular intermediate, and then transfer to new hosts. Int, a tyrosine recombinase, enables site-specific integration of SXT/R391 ICEs into the 5′ end of prfC. Aided by the recombination directionality factor Xis, Int also mediates ICE excision from the host chromosome by recombining the flanking attL and attR sites to regenerate attP and attB (Wozniak et al., 2009). In this article, the schematic representation of insertion sites of ICEPmiChnS012 is shown (Fig. 1).
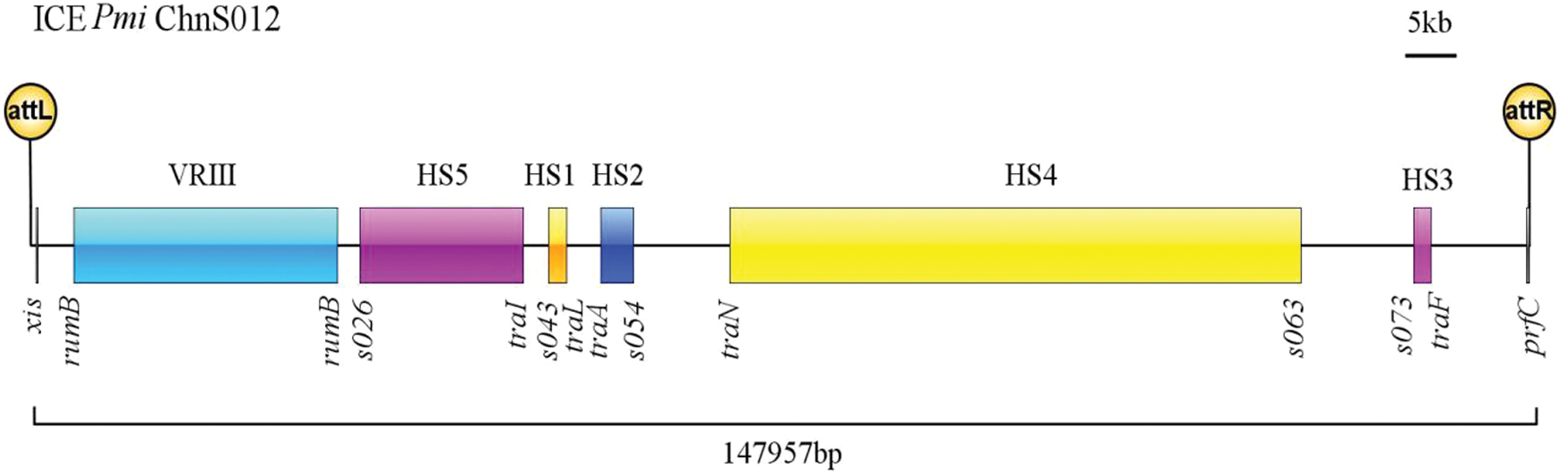
Schematic representation of ICEPmiChnS012. The positions of the site-specific excision (xis) and attachment sites (attL and attR) are indicated. ICE regions (VRIII and HS1-HS5 hotspots) are shown with different colors. Genes located in the left end and right end of each region are represented. ICE, integrative and conjugative element. Color images are available online.
ICEPmiChnBCP11 and ICEPmiChnS012 both harbored a multidrug resistance region in VRIII, containing floR, strA, strB, and sul2 (Fig. 2). ICEPmiChnS012 additionally carried aph3-VI (two copies) and blaHMS-1 , whereas ICEPmiChnBCP11 lacked the two resistance genes in VRIII region. ICEPmiChnS012 contained a new gene cassette array aph3-VI-strB-aph3-VI-strB. The genes floR, strA, strB, and sul2 are frequently found in the VRIII region of SXT/R391 ICE members (Xu et al., 2018). Previous reports showed that bla TEM-1 and bla OXA-1 were the most common and blaHMS-1 was rare in P. mirabilis (Guo et al., 2019). To our knowledge, this is the first report of identification of aph3-VI and blaHMS-1 in the VRIII region in P. mirabilis.

Genetic structure of ICEPmiChnS012 compared with ICEPmiChnBCP11.
Region divergences were also observed in HS5 (Fig. 2). The open reading frames (ORFs) in this hotspot coded for a putative type I restriction–modification system in ICEPmiChnS012 including genes hsdK, hsdS, hsdR, mcr, mcrC, and mrr, whereas restriction–modification system genes met and res and three genes encoding putative protein were found in the corresponding regions in ICEPmiChnBCP11. This fact indicated that ICEPmiChnBCP11 and ICEPmiChnS012 might have a different origin. ICEPmiChnS012 shared HS1, HS2, and HS3 gene contents with ICEPmiChnBCP11 (Fig. 2a).
Both of the ICEs carried an ISPpu12-mediated composite transposon in HS4 containing gene cassettes ISPpu12-tet(A)-tet(R)-IS26 and IS903D-blaCTM-M-65-fip-IS26. Class 1 integron with the dfrA32-ereA1-aadA2 gene cassette array (2763 bp) was seen in ICEPmiChnBCP11. This integron is frequently present in P. mirabilis (Wei et al., 2014; Jiang et al., 2017; Siebor et al., 2018). In ICEPmiChnS012, dfrA1-ereA1-aadA2 gene cassette array (2990 bp) was detected in this study. To the best of our knowledge, this is the first report of dfrA1-ereA1-aadA2 gene cassette in P. mirabilis.
ICEPmiChnS012 and ICEPmiChnBCP11 both carried 10 copies of insertion sequence IS26 (Fig. 2). Previous reports showed that some integrons were bracketed by two copies of IS26 in some P. mirabilis strains (Bontron et al., 2019). They found that increased carbapenemase resistance in P. mirabilis clinical isolates may result from the multiplication of the bla VIM-1 gene through IS26-mediated recombination event. IS26-mediated genetic rearrangements in ICE of P. mirabilis frequently occurred (Wang et al., 2019). We also observed IS26-mediated structures in ICEPmiChnS012 and ICEPmiChnBCP11, such as IS26-fosA3-orf1-orf2-orf3-IS26 and IS26-aac(6′)-Ib-cr-blaOXA-1-catB3-arr2-qacEdelta1-sul1-IS26 (Fig. 2b). Compared with ICEPmiChnBCP11, IS26-fosA3-orf1-orf2-orf3-IS26 was inserted into ICEPmiChnS012 in opposite orientation and different location, indicating an IS26-mediated recombination process.
Similar to ICEPmiChnBCP11, ICEPmiChnS012 harbored 22 different resistance genes including fluoroquinolone resistance gene aac(6′)-lb-cr, fosfomycin resistance gene fosA3, extended spectrum-lactamase gene blaCTX-M-65 , and other resistance genes as given in Table 1. Although ICEPmiChnS012 lacked the clinically important resistance gene cfr (phenicols, lincosamides, oxazolidinones, pleuromutilins, and streptogramin A resistance), it contained more number of resistance gene copies than ICEPmiChnBCP11. ICEPmiChnS012 contained three copies of strB and two copies of aph3-IV, whereas ICEPmiChnBCP11 only contained one copy of strB and one copy of aphA1. In addition, coexistence of bla OXA-1, bla CTX-M-65, and bla HMS-1 was also observed in ICEPmiChnS012 and highlighted the resistance to β-lactams (penicillins and cephalosporins).
In this study, conjugation experiments successfully obtained transconjugants. The conjugation experiment showed that ICEPmiChnS012 could be transferred to E. coli EC600 at a frequency of 3.82 × 10−6 transconjugants per donor. Similar to other SXT/R391 ICEs, ICEPmiChnS012 inserted in the 5′ end of prfC gene (Fig. 3). Breakpoint for streptomycin (minimum inhibitory concentration [MIC] ≥64 μg/mL) was obtained from CLSI. The acquired streptomycin resistance of transconjugants was confirmed by microbroth dilution method (MIC was determined to be 1024 μg/mL).
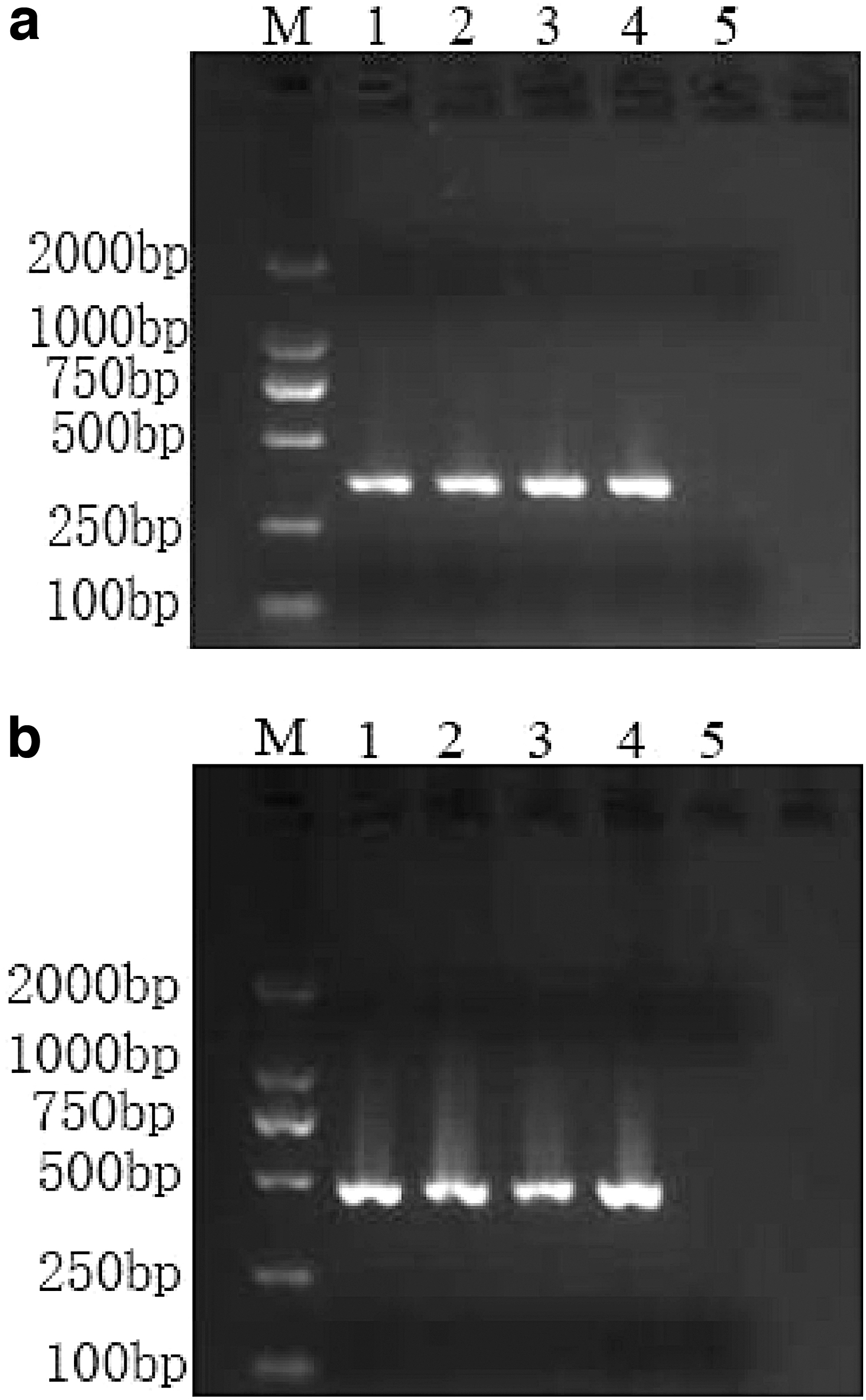
PCR to detect ICEPmiChnS012 insertion in Escherichia coli EC600.
Conclusions
Previous studies involving SXT/R391 elements were focused in multiresistant P. mirabilis isolates from clinical specimens and food-producing animals (Lei et al., 2018; Bontron et al., 2019). However, there is a growing concern over the possibility of transmission of SXT/R391 elements through the food chain. A food-origin P. mirabilis carrying a novel SXT/R391 ICE was reported in this study. To our knowledge, ICEPmiChnS012 was one of the SXT/R391 ICEs that carried the largest number of resistance genes found in P. mirabilis. ICEPmiChnS012 was self-transmissible, indicating it was an important mobile mediator for disseminating resistance genes. ICEs could further enhance the dissemination of resistance genes among Proteus and other Enterobacteriaceae and should receive more attention.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was funded by the National Key R&D Program of China (Grant No. 2017YFC1600100) and partially supported by Chongqing performance guidance fund (19539).