
Abstract
Salmonella is a global foodborne pathogen that causes human diseases ranging from mild gastroenteritis to severe systemic infections. Recently, antimicrobial blue light (aBL) showed effective bactericidal activity against a variety of bacteria (e.g., Salmonella) with varying efficiency. However, the antimicrobial mechanism of aBL has not been fully elucidated. Our previous report showed that the outer membrane (OM) is a key target of aBL. The major component of the OM, lipopolysaccharide (LPS), may play a role in aBL bactericidal effect. Therefore, the influence of LPS truncation on the sensitivity of Salmonella Typhimurium SL1344 to aBL was investigated for the first time. First, the rfaC gene in the SL1344 strain likely involved in linking lipid A to the core region of LPS was inactivated and the influence on LPS structure was verified in the mutant strain SL1344ΔrfaC. SL1344ΔrfaC showed a significant increase in sensitivity to aBL, and the bactericidal efficiency exceeded 8 log CFU at an aBL dose of 383 J/cm2, while that of its parental SL1344 strain approached 4 log CFU. To discover the possible mechanism of higher sensitivity, the permeability of OM was determined. Compared to SL1344, SL1344ΔrfaC showed 2.7-fold higher permeability of the OM at 20 J/cm2, this may explain the higher vulnerability of the OM to aBL. Furthermore, the fatty acid profile was analyzed to reveal the detailed changes in the OM and inner membrane of the mutant. Results showed that the membrane lipids of SL1344ΔrfaC were markedly different to SL1344, indicating that change in fatty acid profile might mediate the enhancement of OM permeability and the increased sensitivity to aBL in SL1344ΔrfaC. Hence, we concluded that disruption of rfaC in Salmonella Typhimurium led to the formation of truncated LPS and thus enhanced the permeability of the OM, which contributed to the increased sensitivity to aBL.
Introduction
S
Antimicrobial blue light (aBL), especially the part with a wavelength ranging from 405 to 470 nm, has been applied in the disinfection of a variety of foodborne pathogens. The aBL light-emitting diodes (LEDs) at 405 or 460 nm have been applied to inactivate Salmonella Typhimurium ATCC 14028 in cantaloupe, and total doses of 1210 J/cm2 and 5356 J/cm2 led to a reduction of 1.1 to 3 log CFU/cm2, respectively (Josewin et al., 2018). LED illumination (405 ± 5 nm) at 1.7 kJ/cm2 decreased to 1.0–1.2 log CFU/cm2 of four Salmonella enterica serovars in papaya (Kim et al., 2017). Besides, more than 5 log 10 of the population of Salmonella Typhimurium ATCC 14028 in milk was achieved by 413 nm aBL irradiation at a dose less than 720 J/cm2 (Dos Anjos et al., 2019). However, as reviewed by Dai et al. (2012), the bactericidal mechanism underlying aBL treatment is still not fully revealed. The action mechanism widely accepted is mediated by the highly cytotoxic reactive oxygen species and subsequent cellular damage (Dai et al., 2012). Our recent report pointed that the outer membrane (OM) and bacterial lipids are important targets during aBL photo-oxidation (Chu et al., 2019). However, it is still unknown which macromolecules within the OM influenced bactericidal efficiency.
OM is an asymmetrical bilayer consisting of an outer leaflet, with lipopolysaccharide (LPS) as a major component and an inner leaflet with a variety of phospholipids. LPS is composed of lipid A, core oligosaccharides, and O-antigen polysaccharide (Whitfield and Trent, 2014). Most enzymes and genes coding for proteins responsible for biosynthesis and export of LPS have been identified. These generally occur in most Gram-negative bacteria based on genetic information (Raetz et al., 2007).
The inner core region of LPS is critical for OM stability (Brabetz et al., 1997). Although the detailed structure of LPS differs from one bacterium to another (Wang and Quinn, 2010), heptose is an important common monosaccharide in the inner-core portion (Wang et al., 2012). In Salmonella and Escherichia coli, the first heptose is added to the inner 3-deoxy-
From the discoveries in aBL and LPS, it could be inferred that LPS truncation by blocking the addition of heptose by WaaC likely influences the bacterial sensitivity to aBL. This is of great significance, considering that bacteria always modify the LPS structure in response to local environments, and that the polysaccharide part of LPS is the most flexible region (Patel et al., 2016).
In the current study, the gene rfaC that was probably responsible for heptose addition on the inner core of LPS in Salmonella Typhimurium SL1344 was knocked out, and its influence on bacterial sensitivity to aBL was investigated for the first time. Later, the permeability of OM and fatty acid profile were also analyzed to reveal the possible reason for the enhanced sensitivity of the LPS mutant. We concluded that the significant fluctuation of fatty acid profile, as well as the stimulation of the permeability of OM, contributed to the sensitivity of mutant Salmonella Typhimurium SL1344 to aBL. The current study will help discover the principal factors influencing bactericidal efficiency of aBL, and the detailed mechanism underlying aBL disinfection.
Materials and Methods
Bacterial strains, plasmids, and growth condition
The bacterial strains and plasmids used in the present study are listed in Table 1. All strains were grown in LB media (10 g/L tryptone, 5 g/L yeast extract, and 10 g/L NaCl at pH 7) at 37°C. As an exception, the strains transformed with the temperature-sensitive plasmid pKD46 or pCP20 were cultured at 30°C. When needed, ampicillin, kanamycin, and chloramphenicol were added to final concentrations of 100, 30, and 25 μg/mL, respectively. The growth rate was determined according to our previous method (Wang et al., 2012) with minor modifications as follows. Salmonella Typhimurium SL1344 was cultivated on LB agar plates (LB media plus 1.5% agar) for 1–2 d. A single colony was then picked up and incubated in 10 mL liquid LB media at 200 rpm to an optical density of 0.5 at 600 nm (OD600). Later, the seed culture was inoculated into 50 mL of LB media, and the initial OD600 was adjusted to 0.02. The OD600 was detected every 2 h, and the biomass curve was plotted.
Bacterial Strains and Plasmids Used in the Current Work
Knockout of rfaC gene by the Red recombination system
Using the sequence of waaC of E. coli W3110 as a template, a BLASTP search against GenBank data revealed that the putative waaC gene encoding heptosyltransferase I in Salmonella Typhimurium SL1344 was rfaC. Therefore, the rfaC gene was inactivated by the Red recombination system reported in our previous article (Wang et al., 2012), with some modifications as follows.
All of the primers used in this work are listed in Table 2. First, three fragments of rfaC-U, rfaC-D, and kanF obtained by PCR were digested with the corresponding endonucleases, respectively, and inserted into pBluescript II SK (+) to create the plasmid pBS-rfaC-kanF (Wang et al., 2012). Next, the DNA fragment CU-kanF-CD was amplified from pBS-rfaC-kanF and transformed into Salmonella Typhimurium SL1344 containing the λ Red helper plasmid pKD46. After homologous recombination, pKD46 in the transformants was then removed by thermal stimulus at 42°C, and the rfaC mutant strain of Salmonella Typhimurium SL1344 was created and designated as SL1344ΔrfaC::kan (Fig. 1). Finally, the kanamycin resistance cassette was removed from the chromosome of SL1344ΔrfaC::kan to yield SL1344ΔrfaC (Fig. 2). The interruption of rfaC in both mutants SL1344ΔrfaC::kan and SL1344ΔrfaC was confirmed by PCR amplification using primer pairs rfaC-U-F and rfaC-D-R (Fig. 2, inset). For the complementation experiment, pWSK29-rfaC was transformed into SL1344ΔrfaC to obtain SL1344ΔrfaC/pWSK29-rfaC.

Procedure of inactivating rfaC gene by Red recombination system. kan, kanamycin resistance gene; FRT, FLP recognition target.
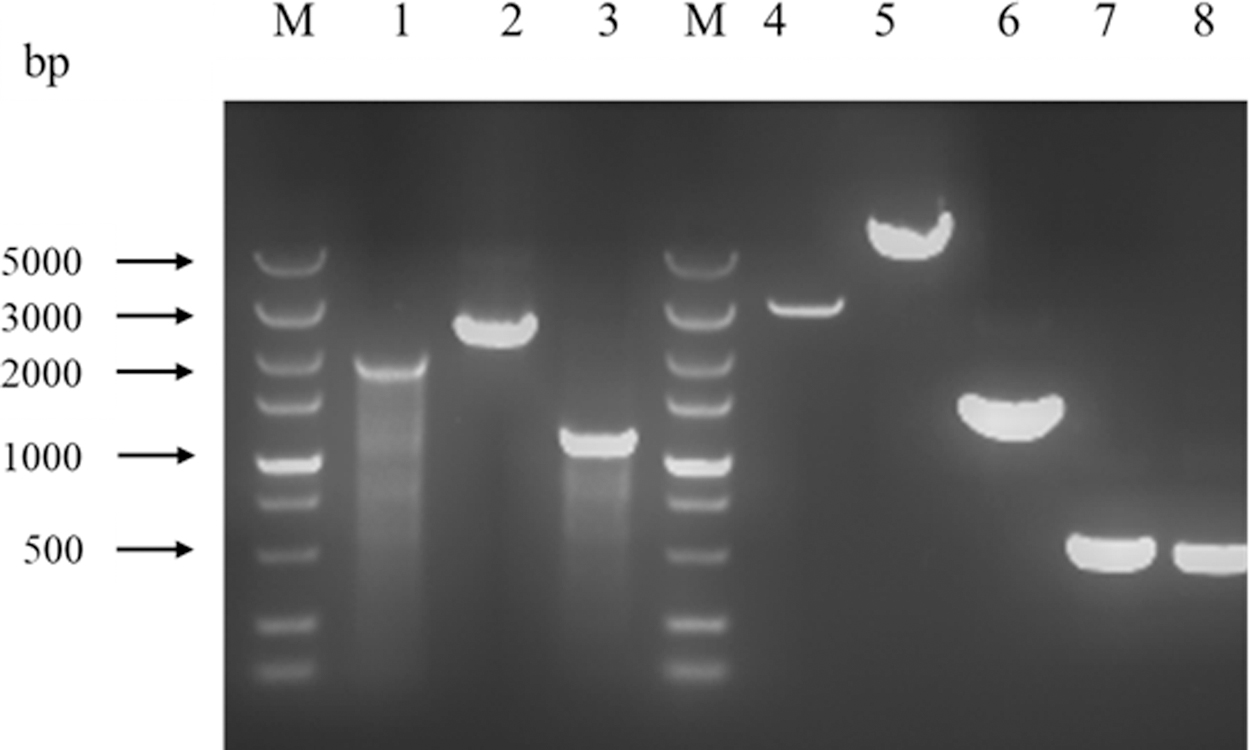
Agarose gel electrophoresis of sizes of DNA fragments by PCR amplification. M, DNA marker; Lane 1, PCR amplification product of SL1344 using primers rfaC-U-F/rfaC-D-R; Lane 2, PCR amplification product of SL1344ΔrfaC::kan using primers rfaC-U-F/rfaC-D-R; Lane 3, PCR amplification product of SL1344ΔrfaC using primers rfaC-U-F/rfaC-D-R; Lane 4, pBluescript digested by XhoI; Lane 5, pBluescript-rfaC-KanF digested by XhoI; Lane 6, PCR product of KanR; Lanes 7 and 8, PCR products of homologous fragment of rfaC.
Primers Used in the Current Study
The restriction site was underlined.
LPS purification and sodium dodecylsulfate polyacrylamide gel electrophoresis analysis
LPS was extracted using the hot phenol/water method (Kido et al., 1990) as follows. The cells of Salmonella Typhimurium (400 mL of overnight culture) were harvested by centrifugation at 4000 rpm for 10 min, then resuspended in 20 mL of ddH2O, and then mixed with 20 mL of preheated 90% phenol at 68°C. The supernatant fluid was transferred into dialysis bags, and dialyzed in dd H2O for 24 h. The fluid was vacuum-freeze-dried to obtain the crude LPS sample (Wang et al., 2020). Finally, LPS was analyzed using silver-stained sodium dodecylsulfate polyacrylamide gel electrophoresis (SDS-PAGE) (Tsai and Frasch, 1982) using the same procedure reported in our previous article (Wang et al., 2012).
The aBL irradiation of SL1344 and the mutant strain
The effect of LPS truncation on aBL sensitivity was studied using the aBL bactericidal method (Chu et al., 2019). First, bacterial cell pellets harvested from overnight culture were washed three times with phosphate-buffered saline (PBS) at 0.2 M and pH 7.4, and the cell suspension (OD600 at 0.1) was prepared. Four milliliters of cell suspension was placed in a Petri dish (bottom outside diameter of 35 mm and height of 16 mm) to receive aBL irradiation. Later, aBL illumination was conducted according to the previous method (Wu et al., 2018). The Omnilux clear-UTM LED array (Photo Therapeutics, Inc., Carlsbad, CA, USA) was used as light source, with a central wavelength of 415 nm and a full width at half maximum of 20 nm. The distance between the LED and the sample was 1.8 cm, and the incremental doses, including 27, 54, 82, 164, 273, 328, and 383 J/cm2, were used. The Petri dish was put on ice to keep the bacterial suspension at 0–7°C. During light irradiation, the suspension was gently stirred by a mini-magnetic bar (Fisher Scientific Co., Norcross, Georgia, USA) at 30 rpm. A portion of the cell suspensions at different time points (20 μL) was analyzed to calculate the survival rate.
Hydrophobicity assay
The cell surface hydrophobicity of the SL1344 and SL1344ΔrfaC cells was assayed using the methods reported by Zavaglia et al. (2002), and Wang et al. (2012). Bacterial cell pellets harvested from overnight culture were washed three times with PBS (pH 7.4), and the OD600 values of the bacterial cells before and after xylene treatment were measured to calculate the hydrophobicity of the bacterial cell.
OM permeability
The permeabilities of Salmonella SL1344 and SL1344ΔrfaC were assayed according to the reported method (Vaara, 1990; Wang et al., 2012). The cell suspension was quickly mixed with the N-phenylnaphthylamine (NPN) purchased from Sigma-Aldrich (St. Louis, MO, USA), and the fluorescence of the cell mixture was monitored immediately by a spectrofluorometer (Hitachi, Tokyo, Japan). The excitation wavelength, emission wavelength, and slits used were 350, 420, and 5 nm, respectively.
Extraction and analysis of fatty acids
The cellular fatty acids were extracted and analyzed using methods reported previously (Li et al., 2010; Wang et al., 2012). After isolation, the fatty acids were then derivatized by N,O-bis(trimethylsilyl)trifluoroacetamide and analyzed by GC–MS.
Results and Discussion
Knockout of the rfaC gene
In LPS biosynthesis in E. coli, heptosyltransferase I was responsible for adding a heptose to Kdo2-lipid A. Thus, in the current study, BLASTp search was carried out by using waaC sequence of E. coli against the genome of Salmonella Typhimurium SL1344. The gene rfaC encoding a protein with identity of 78% was screened out. This indicated that it was the putative gene waaC in Salmonella Typhimurium SL1344. We constructed the mutants SL1344ΔrfaC::kan and SL1344ΔrfaC by inserting kanF or FRT sequence at the site 952 bp downstream to the 5′ end of the rfaC gene (Fig. 1). SL1344ΔrfaC::kan was resistant to kanamycin, while SL1344ΔrfaC was sensitive due to the removal of the kanamycin-resistant cassette. The inactivation of rfaC was tested by PCR (Fig. 2). The sizes of DNA fragments amplified from both strains were increased, in accordance with the insertions of kanF or FRT in the chromosome. The knock-in of FRT in the chromosome of SL1344ΔrfaC was also confirmed by DNA sequencing.
Verification of LPS truncation by SDS-PAGE and characterization of the mutant strain
The deficiency of RfaC in SL1344ΔrfaC was tested by analyzing the LPS structure (Fig. 3). LPS isolated from strains SL1344 and SL1344ΔrfaC was assayed by SDS-PAGE with silver staining (Tsai and Frasch, 1982). The typical band of O-antigen repeats appeared at the top of the LPS sample from SL1344, indicating that it contained the complete LPS, including lipid A, core region, and O-antigen. However, the samples isolated from SL1344ΔrfaC did not show an O-antigen band, suggesting that it had no O-antigen repeats. Because the O-antigen is connected to the outer core region of the core polysaccharide, the deletion of the rafC gene led to the termination of O-antigen addition. Therefore, SL1344ΔrfaC did not contain an O-antigen band, confirming that O-antigen linkage was interrupted and only the truncated LPS was produced.
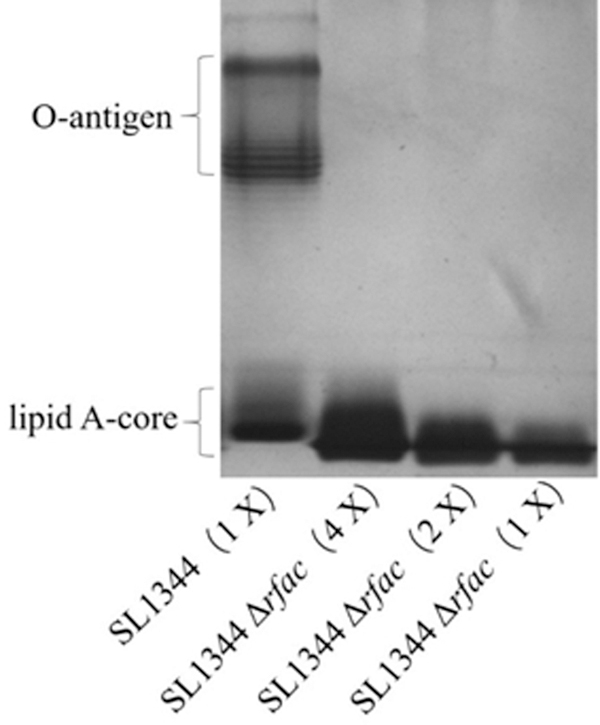
The silver-stained SDS-PAGE analysis of LPS purified from Salmonella Typhimurium SL1344 and SL1344ΔrfaC (1 × , 2 × , and 4 × ). LPS extracted by the hot phenol/water method was analyzed by PAGE in the presence of SDS and then stained with Ag. The classical band of O-antigen was displayed on the top and lipid A-core on the bottom. LPS, lipopolysaccharide; SDS-PAGE, sodium dodecylsulfate polyacrylamide gel electrophoresis.
In the present study on Salmonella, cleavage of LPS slightly delayed the growth. As shown in Figure 4A, compared with SL1344, SL1344ΔrfaC displayed a slightly lower growth rate. The peak biomass of SL1344ΔrfaC was obtained at 3.92 g/L, 4.23% lower than that of SL1344. However, in our previous report on Cronobacter sakazakii, LWW02 strain deficient in WaaC showed a growth rate higher than that of wild-type BAA894 strain (Wang et al., 2012). These results indicated that LPS truncation could exert a strain-dependent influence on growth.
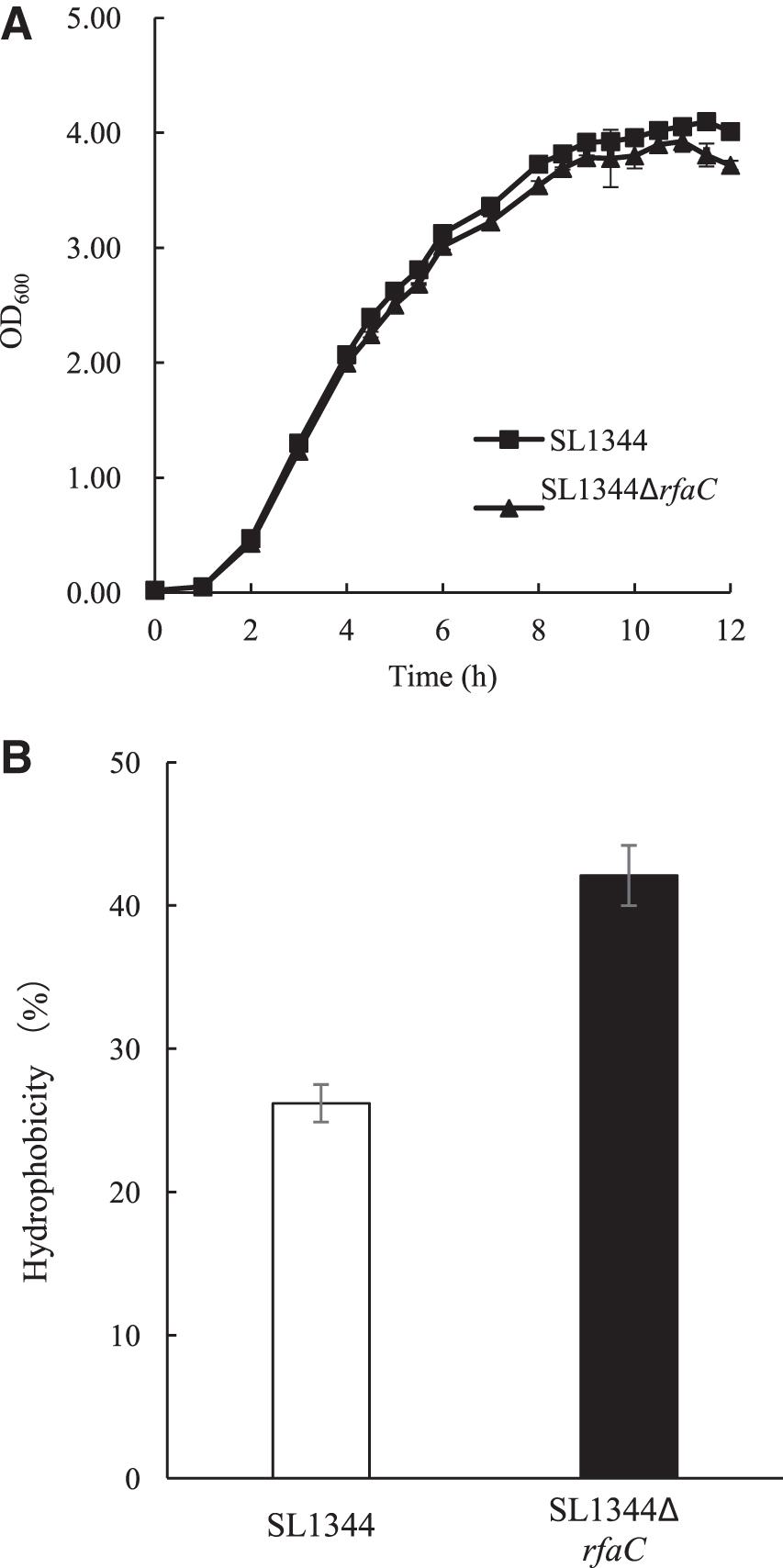
The growth rate
Because LPS was the major macromolecule covering the outer leaflet of the OM, SL1344ΔrfaC most likely had a cell surface with distinct characteristics, therefore, the hydrophobicity of the cell surface was assayed (Fig. 4B). As expected, the surface of the SL1344 cells showed hydrophobicity of 26.2%, while SL1344ΔrfaC cells displayed 60.6% higher hydrophobicity at 42.1%. The enhancement of hydrophobicity was mainly due to the loss of the hydrophilic core region and O-antigen of LPS, which is similar to the results of our previous work on C. sakazakii (Wang et al., 2012).
Sensitivity to aBL
In recent years, aBL irradiation has emerged as a promising alternative to antibiotics. It thus has been of interest, being a nonspecific bactericidal method for killing wounds and oral pathogenic bacteria, including various drug-resistant bacteria (Dai et al., 2012). Recently, aBL was also shown to have effective bactericidal activity against foodborne pathogens, and the underlying mechanism was explored (Kim et al., 2015, 2016; Ghate et al., 2017; Kim and Yuk, 2017; Wu et al., 2018; Chu et al., 2019). Our previous work revealed that lipids and membrane damage play a critical role in the bactericidal effect (Chu et al., 2019). Thus, in current study, the role of LPS, the major component on the cell surface of Gram-negative bacteria, was further investigated. The inactivation curves of SL1344 (with complete LPS), SL1344ΔrfaC (with truncated LPS), and the complementation strain SL1344ΔrfaC/pWSK29-rfaC under aBL treatment were compared and shown in Figure 5.
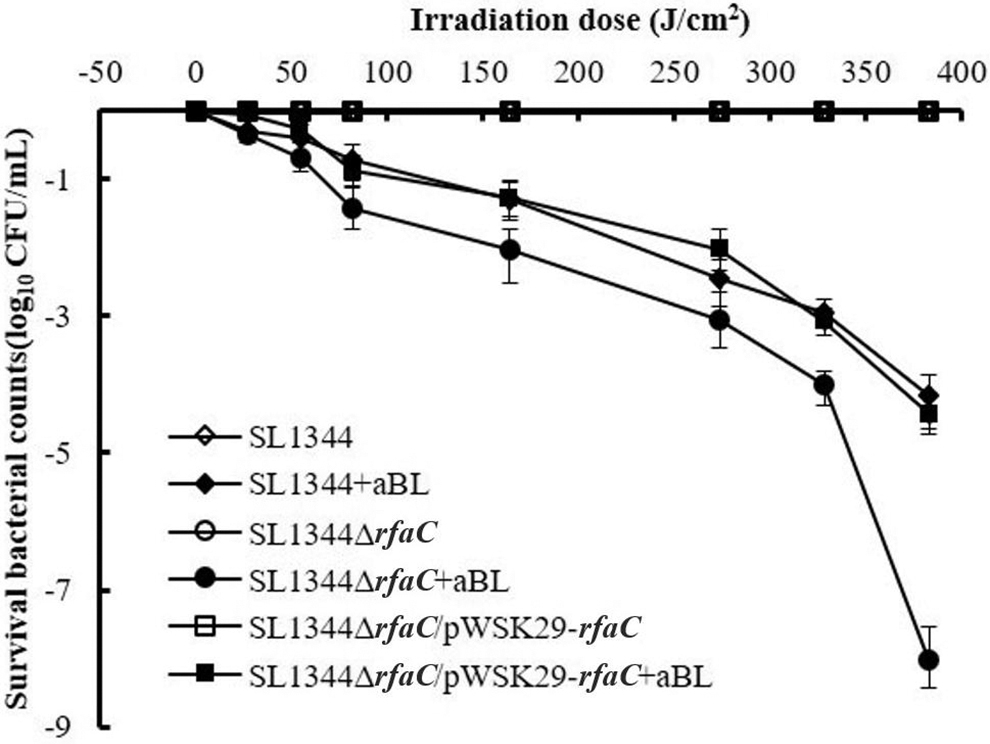
Comparison of bactericidal curves of Salmonella Typhimurium SL1344, SL1344ΔrfaC, and SL1344ΔrfaC/pWSK29-rfaC by aBL. At different irradiance doses of 0, 27, 54, 82, 164, 273, 328, and 383 J/cm2, 20 μL of suspension was withdrawn, and CFU was then assayed by serial dilutions on LB agar plates. Experiments were performed in triplicate, and the data are presented as means ± standard deviation. aBL, antimicrobial blue light.
As shown in Figure 5, the population of nonilluminated Salmonella remained almost unchanged, while that of aBL-illuminated Salmonella decreased gradually along with irradiation of aBL. Moreover, SL1344 and SL1344ΔrfaC showed different sensitivities. For example, at 164 J/cm2, the survival rate of SL1344 was detected as 22.4%, while that of SL1344ΔrfaC was 4.5%. When the aBL dose was elevated to 383 J/cm2, the logarithmic reduction of SL1344 cells approached 4 log CFU, whereas that of SL1344ΔrfaC exceeded 8 log CFU, significantly higher than that of the former. Thus, it could be seen that LPS truncation in SL1344ΔrfaC significantly enhanced the sensitivities to aBL. To confirm the effect of rfaC on the sensitivity to aBL, the bactericidal curve of the complementation strain SL1344ΔrfaC/pWSK29-rfaC was also studied. As expected, the complementation strain showed comparable sensitivity to SL1344.
To our knowledge, this is the first report of the influence of LPS length on bacterial sensitivity to aBL. In response to the highly variable and complex microenvironment, Gram-negative bacteria used multiple mechanisms (both at the genetic and epigenetic levels) to enrich the LPS diversity by modifying LPS to adapt to the ever-changing circumstances (Maldonado et al., 2016). Recent work showed that the dynamic structure of LPS was highly dependent on local environments, with the polysaccharide region being the most flexible (Patel et al., 2016). On the basis that the length of LPS was dependent on environmental stimuli, it was of great significance to discover why LPS truncation could affect bacterial sensitivity to aBL. In the following section, therefore, the possible reason is explored.
Permeability of OM
In our previous studies, 415 nm aBL irradiation higher than 5 J/cm2 caused the cell surface damage in methicillin-resistant Staphylococcus aureus (Wu et al., 2018), and an aBL dose of more than 30.06 J/cm2 led to inactivation of C. sakazakii BAA894 (Chu et al., 2019). The permeability of OM, an indicator of injury of cell surface, was first measured herein. In the treatment with NPN, NPN penetrated the cell surface and located at OM. The changes in fluorescence were thus correlated with the penetrability of the OM.
As shown in Figure 6, the permeability of the OM was not significantly changed during 20 min of light-treatment experiment. SL1344 and SL1344ΔrfaC strains displayed markedly different permeabilities. Before aBL irradiation, SL1344ΔrfaC showed 2.1-fold greater permeability than that of SL1344, and during the process of aBL illumination, SL1344ΔrfaC also showed a higher increase rate. Consequently, SL1344ΔrfaC had 2.7-fold higher permeability at 20 J/cm2, Therefore, increased permeability may be the main reason for the higher sensitivity to aBL. The knockout of gene ESA_04107 in C. sakazakii ATCC BAA-894 led to cleavage of LPS and caused OM defects (Wang et al., 2012). Similarly, in the current study, inactivation of rfaC in Salmonella also resulted in LPS truncation and OM injury.
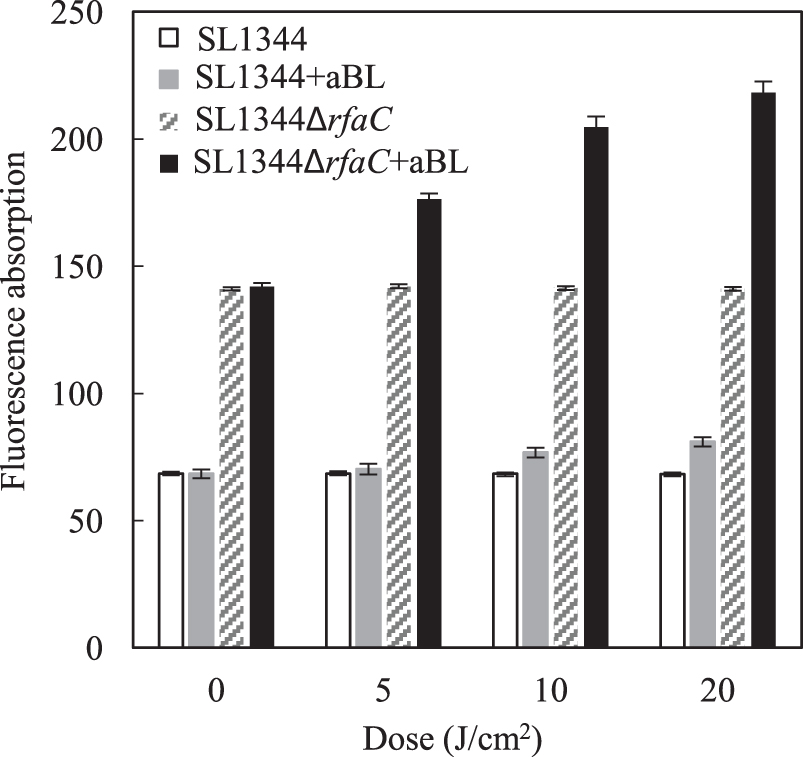
Comparison of the permeabilities of OM between Salmonella Typhimurium SL1344 and SL1344ΔrfaC. At sublethal doses of BL at 0, 5, 10, and 20 J/cm2, 200 μL of the bacterial suspension was collected and centrifuged. The bacterial cells were washed and resuspended in PBS, and the hydrophobic probe NPN was added to the bacterial suspension. NPN was localized at the OM, and thus the fluorescence changes were recorded to reflect changes in OM permeability. Experiments were performed in triplicate, and the data are presented as means ± standard deviation. BL, blue light; NPN, N-phenylnaphthylamine; OM, outer membrane; PBS, phosphate-buffered saline.
Fatty acid profile of the wild-type and mutant strains
To reveal the changes in the OM of SL1344ΔrfaC, GC–MS was used to analyze the fatty acids of the two strains. As shown in Table 3, both strains showed distinct distribution patterns of fatty acids, and a large variation was apparent in SL1344ΔrfaC. As shown in Table 3, nine fatty acid species were observed in the control SL1344, while only seven were found in the mutant SL1344ΔrfaC. Three fatty acids were undetectable in SL1344, namely, C12:0, C16:1, and C18:0, while another fatty acid, C15:0 appeared in SL1344. Moreover, the percentages of fatty acids were dramatically changed, such as C14:1 in SL1344ΔrfaC, with an 18-fold higher level compared with SL1344.
Comparison of Fatty Acids Profiles of Salmonella Typhimurium SL1344 and SL1344ΔrfaC
Not detectable.
The biophysical properties of the OM and the inner membrane (IM) are characterized by the fatty acids that are incorporated into the component phospholipids (Zhang and Rock, 2008). Phospholipids refer to lipids containing phosphoric acid, and they are the main components of OM and IM. Bacteria have the ability for adjusting the biophysical properties of their membrane phospholipids in response to and in thriving in a wide range of physical environments. The asymmetric OM of Gram-negative bacteria is composed of phospholipids in the inner leaflet and LPS in the outer leaflet. The variation of LPS may influence the composition of phospholipids, which contain a variety of fatty acids. Bacteria precisely modify the lipid composition within membranes by changing the types of fatty acids and by altering the structures of phospholipids (Zhang and Rock, 2008). The close relationship between bacterial phospholipid biosynthesis and LPS biosynthesis was presented on the basis of a computational model (Emiola et al., 2016). It was interesting that LPS truncation in Salmonella led to a significant change in fatty acid profiles.
Similarly, in our previous report (Wang et al., 2012), C. sakazakii LWW02 deficient in heptosyltransferase I produced only Kdo2-lipid A. The truncation of LPS dramatically affected the cells by changing the properties of the cell surface and cell morphology. In scanning electron microscopy (SEM) analysis, C. sakazakii LWW02 showed more fiber-like materials around cells than did BAA-894 cells (Wang et al., 2012). In the current study, the SEM analysis of Salmonella Typhimurium SL1344ΔrfaC was done in our laboratory to further understand the morphological variation. Results showed that the inactivation of rfaC gene in Salmonella Typhimurium SL1344ΔrfaC also led to more distinct permeability and hydrophobicity of the cell surface compared with the wild-type SL1344.
The truncation of LPS also influenced the bacterial sensitivity to some antimicrobials, according to our previous report (Wang et al., 2015). On the comparison of the susceptibility of the 10 LPS mutant strains of E. coli to erythromycin and novobiocin, the mutants ΔwaaC, ΔwaaF, and ΔwaaG were significantly sensitive to both antibiotics, indicating that the shortened LPS changed the OM structure and made the cell surface more fragile (Wang et al., 2015). Similar results were also observed in the current study: Salmonella Typhimurium SL1344ΔrfaC also became more sensitive to erythromycin and novobiocin (data not shown), although the underlying mechanism needs to be investigated.
On the basis of our data, we hypothesized that the dramatic fluctuation in fatty acid profiles might result from LPS variation. The enhanced permeability of the OM may be the crucial reason for the enhanced sensitivity of SL1344ΔrfaC to aBL, while more investigations are needed in the future to confirm the detailed mechanisms and to reveal the roles of LPS modification in regulating bacterial sensitivity to aBL. In the future, a variety of Salmonella serovars isolates will be collected and their respective LPS mutants will be constructed and analyzed to reveal the role(s) of rfaC gene in bacterial sensitivity to aBL.
Conclusion
The aBL irradiation has been regarded as a novel effective antimicrobial approach against various bacteria, with a strain-dependent bactericidal efficiency. The current study investigated the influence of LPS truncation and explored the possible reason for the first time. Knockout of rfaC in Salmonella Typhimurium SL1344 led to LPS truncation and higher sensitivity to aBL. It is likely that the changes of OM permeability aggravated the damage to the OM by aBL. The enhancement of OM permeability was accompanied by an apparent change in fatty acid profile. It was concluded that rfaC inactivation caused truncated LPS and higher OM permeability, which mostly likely contributed to increased sensitivity of Salmonella Typhimurium to aBL. These findings will help to discover the critical roles of LPS in bacterial sensitivity to aBL and improve the bactericidal efficiency of aBL.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by grants from the National Key R&D Program of China (2017YFC1600100), the National Natural Science Foundation from China (no. 31201290), the China Postdoctoral Science Foundation, the First-Class Discipline Program of Light Industry Technology and Engineering (LITE2018-10), and the Program of SKLFZZB-201905.