
Abstract
Antibiotic-resistant infections caused by extended-spectrum β-lactamases (ESBLs) and carbapenemases are increasing worldwide. Bacteria resistant to extended-spectrum cephalosporins and last resort carbapenems have been reported from food animals and their environments. Other concentrated nonfood-producing animals such as mink farming can be a reservoir of bacteria resistant to these critically important antibiotics. The objective of this study was to determine the prevalence of ESBL-producing bacteria and carbapenem-resistant (CR) bacteria from mink fecal (n = 42) and feed (n = 8) samples obtained from a commercial mink farm in the United States. The most prevalent ESBL-producing bacteria identified from the fecal samples were Escherichia coli (93%), Klebsiella pneumoniae (76%), and Proteus species (88%). E. coli (100%) and K. pneumoniae (75%) were also the most prevalent ESBL-producing bacteria identified from feed samples. All ESBL E. coli isolates were resistant to penicillin and most cephem beta-lactam antibiotics. Among the ESBL E. coli isolates, co-resistance was observed to ciprofloxacin (33%) and gentamicin (28%) indicating multidrug resistance. ESBL E. coli isolates predominantly carried bla CTX-M-14 and bla CTX-M-15 genes. Although all feed K. pneumoniae isolates carried bla CTX-M-9, all fecal K. pneumoniae isolates carried bla SHV. CR Pseudomonas species (7%), Hafnia alvei (24%), and Myroides odoratimimus (9.5%) were detected from fecal samples. H. alvei (37.5%) was the only CR bacteria detected from the feed samples. All CR isolates were polymerase chain reaction negative for the tested carbapenemases that are commonly reported, which may indicate intrinsic rather than acquired resistance. This study indicates that mink production can be a reservoir for bacteria resistant to the highest priority critically important antibiotics for human health.
Introduction
Beta-lactam antibiotics play a significant role for the treatment of bacterial infections since the discovery of the first antibiotics. They represent two-thirds of hospital antibiotic prescriptions in the United States (Bush and Bradford, 2016). Penicillins and cephalosporins are also commonly used β-lactams in food animal production (FDA, 2020). However, the beneficial roles played by the β-lactams is threatened because of increased occurrence of bacterial resistance to these antibiotics. Primary mechanism of bacterial resistance is through the production of β-lactamase enzymes that inactivate the β-lactam ring of these antibiotics (Bush and Jacoby, 2010; Bush and Bradford, 2020). Extended-spectrum β-lactamases (ESBLs) confer resistance to penicillin and extended-spectrum cephalosporins (third and higher generations) that are commonly used to treat serious infections in humans (Bush, 2018).
Resistance to extended-spectrum cephalosporins is conferred by some variants of class A (encoded by bla CTX-M, bla SHV and bla TEM genes) and class D (encoded by bla OXA) β-lactamases (Bush and Jacoby, 2010; Bush, 2018). ESBL-producing Enterobacteriaceae widely occurred in humans. They caused an estimated 197,400 hospitalizations, 9,100 deaths, and $1.2 billion attributable health care costs in 2017 in the United States alone. Infections caused by ESBL-producing Enterobacteriaceae have significantly increased during 2012–2017. As a result, the U.S. Centers for Disease Control and Prevention (CDC) considers ESBL-producing Enterobacteriaceae as serious public health threats (CDC, 2019).
Carbapenems are the newest β-lactams of the last resort to treat bacterial infections resistant to most antibiotics including those caused by ESBL-producing bacteria (Ogunrinu et al., 2020). However, carbapenem resistance (CR) is increasing owing to their increased use because of increased occurrence of infections caused by ESBL-producing bacteria (Bush and Bradford, 2020). CR Acinetobacter and CR Enterobacteriaceae are among the five bacteria that were identified as urgent threat by the CDC. In the United States alone, CR Enterobacteriaceae caused an estimated 13,100 hospitalizations, 1100 estimated deaths, and $130 million attributable health care costs in 2017 (CDC, 2019). Third-, fourth-, and fifth- generation cephalosporins, carbapenems, and other penems are categorized by the World Health Organization as the highest priority critically important antimicrobials (WHO, 2019). Although carbapenems have not been approved for use in food animals in the United States, CR bacteria have been reported from food animals and their environments (Webb et al., 2016; Mollenkopf et al., 2017, 2018).
CR bacteria have a limited transmission from nonhuman sources. However, people can acquire infections caused by ESBL-producing Enterobacteriaceae through the food chain, direct contact with infected animals, exposure through recreational water contaminated by animal manure, through runoff from animal production facilities, and ingestion of produce contaminated through the use of animal manure (Crim et al., 2018; Cho et al., 2020). Manure from concentrated animal production facilities including mink production is an important source of bacteria. Environmental, recreational and drinking water contaminations can occur from manure runoff from the mink farms or when improperly treated farm wastes including dead animals are land applied as soil amendment. However, studies on bacteria resistant to critically important antibiotics including extended-spectrum cephalosporins and carbapenems are scarce in the mink production system. Thus, the objective of this study was to investigate the occurrence of ESBL-producing bacteria including Enterobacteriaceae and CR bacteria in mink feces and feed samples.
Materials and Methods
Fecal samples (n = 42) representing seven houses (six samples/house) were obtained from the under the cage of individual mink raised at a large commercial mink farm in the United States. In addition, eight bulk feed samples were obtained. Institutional animal care and use committee review was not required since the fecal samples were collected from the floor beneath the cages with no manipulation of the animals. Detailed sampling and farm descriptions were previously reported. The prevalence of the resistant bacteria and Salmonella was analyzed in Stata 16.1 (StataCorp, LLC, College Station, TX) as the proportion of positive samples.
Detection of ESBL-producing and CR bacteria
Primary enrichments were made by transferring 10 g of feces and feed to filter bags (Nasco Whirl-Pack, Fort Atkinson, WI) and suspending in 90 mL of buffered peptone water (BPW; Becton, Dickinson and Company [BD], Franklin Lakes, NJ). The suspensions were homogenized in a laboratory blender (Stomacher 400 Circulator; Seward Laboratory Systems, Inc., Islandia, NY) at 200 rpm for 15 s. BPW suspensions were incubated at 25°C for 2 h, followed by 42°C for 6 h and held at 4°C until secondary enrichments were performed the following day (Chaney et al., 2017). The BPW pre-enrichments were streaked onto CHROMagar ESBL and CHROMagar KPC (both from DRG International, Inc., Springfield, NJ) for the detection of ESBL-producing and CR enteric bacteria, respectively. Plates were incubated at 37°C for 24 h. ESBLs E. coli were confirmed by a multiplex polymerase chain reaction (PCR) primers and protocol targeting lacY, lacZ, cyd, and uidA genes (Bej et al., 1991; Horáková et al., 2006; Horakova et al., 2008) from a 2 μL isolate DNA obtained by BAX lysis method following the manufacturer's instructions (DuPont Qualicon, Inc., Wilmington, DE). PCR products were analyzed by electrophoresis on 2% agarose gels in 1 × Tris-borate-ethylenediaminetetraaceticacid (TBE) buffer and visualized using BioSpectrum Multispectral imaging system (UVP, Upland, CA). Other ESBL-producing bacteria and CR isolates were identified by matrix-assisted laser desorption ionization time-of-flight mass spectrometry using MALDI Biotyper (Bruker Daltonics, Inc., Billerica, MA) at the National Veterinary Services Laboratories ([NVSL]; United States Department of Agriculture, Ames, IA).
Antimicrobial susceptibility testing
Minimum inhibitory concentrations for 16 antimicrobial agents were determined at the NVSL for each isolate by broth microdilution method using the Trek Sensititre® ESBLs panel (catalog#: ESB1F) in Sensititre automated antimicrobial susceptibility testing system according to the manufacturer's instructions (Trek Diagnostics; Thermo Fisher Scientific, Inc., Cleveland, OH). Except for cephalothin, Clinical and Laboratory Standards Institute (CLSI)'s resistance breakpoints (CLSI, 2020) were used. For cephalothin, National Antimicrobial Resistance Monitoring (NARMS)'s breakpoint of ≥32 μg/mL was used (CDC, 2018). Multidrug resistance (MDR) was defined as resistance to ≥3 antimicrobial classes (Agga and Scott, 2015) based on the six classes included on the ESB1F panel according to the CLSI classification (Supplementary Table S1).
Detection of antibiotic resistance genes
ESBL isolates were tested for ESBLs genes using four multiplex PCR assays (Streck ARM-D Kits, La Vista, NE) following the manufacturer's instructions. The first assay consisted of bla CMY-2, bla CTX-M-14, and bla CTX-M-15. The second assay consisted of bla OXA-48, bla IMP, and bla VIM. The third assay consisted of bla DHA, bla KPC, and bla NDM. The fourth assay consisted of bla TEM, bla SHV, and bla GES genes. Isolates negative for any of the genes in the four assays were tested by PCR for other bla CTX-M alleles using published primers and protocols (Cormier et al., 2019). CR isolates were tested for carbapenemase genes bla KPC, bla NDM, bla OXA-48, bla VIM, and bla IMP (Precit and others 2020), bla KPC-2 (Vikram et al., 2017), bla OXA-58 (Szczepanowski et al., 2009), using published PCR primers and protocols.
Results
Prevalence of ESBL-producing and CR bacteria
Table 1 shows the prevalence and species composition of ESBL-producing and CR bacteria isolated from mink feces and feed samples. ESBLs E. coli, K. pneumoniae, and Proteus were the most prevalent species detected (>75% prevalence) from the mink fecal samples. While E. coli and K. pneumoniae were also the most prevalent (>75% prevalence) in the feed, Proteus species were not detected. Although low in prevalence, CR bacteria belonging to three genera (Pseudomonas, Hafnia, and Myroides) were also detected. Pseudomonas and Myroides odoratimimus were detected only from the fecal samples; Hafnia alvei was isolated both from feces and feed samples.
Prevalence of Extended-Spectrum β-Lactamase-Producing Bacteria from Mink Fecal and Feed Samples in the United States
Expressed as the number of positive samples (% positive).
ESBL, extended-spectrum β-lactamase.
Antimicrobial resistance of ESBL-producing and CR bacteria
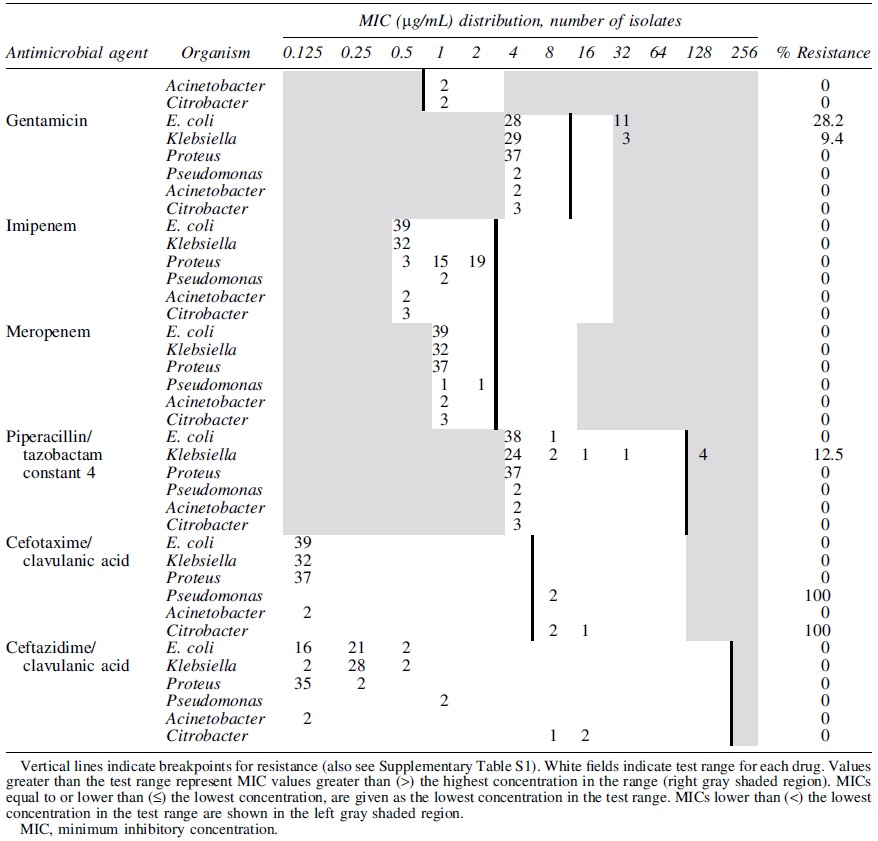
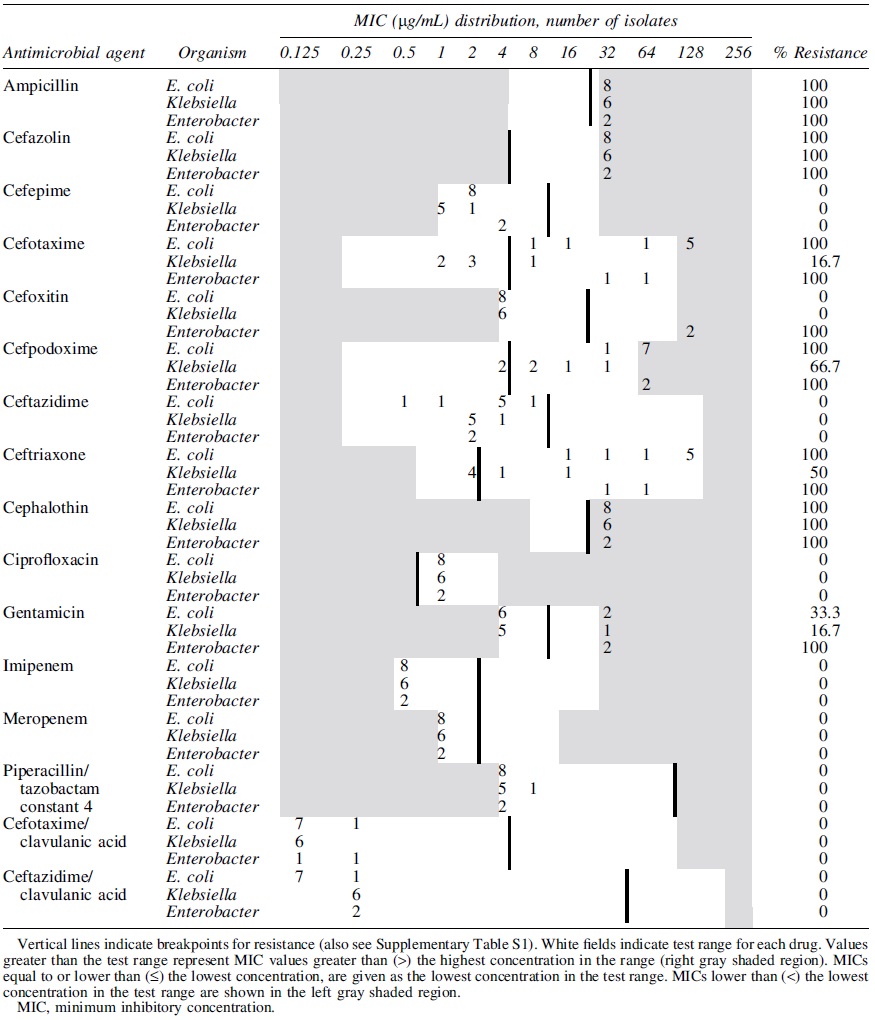
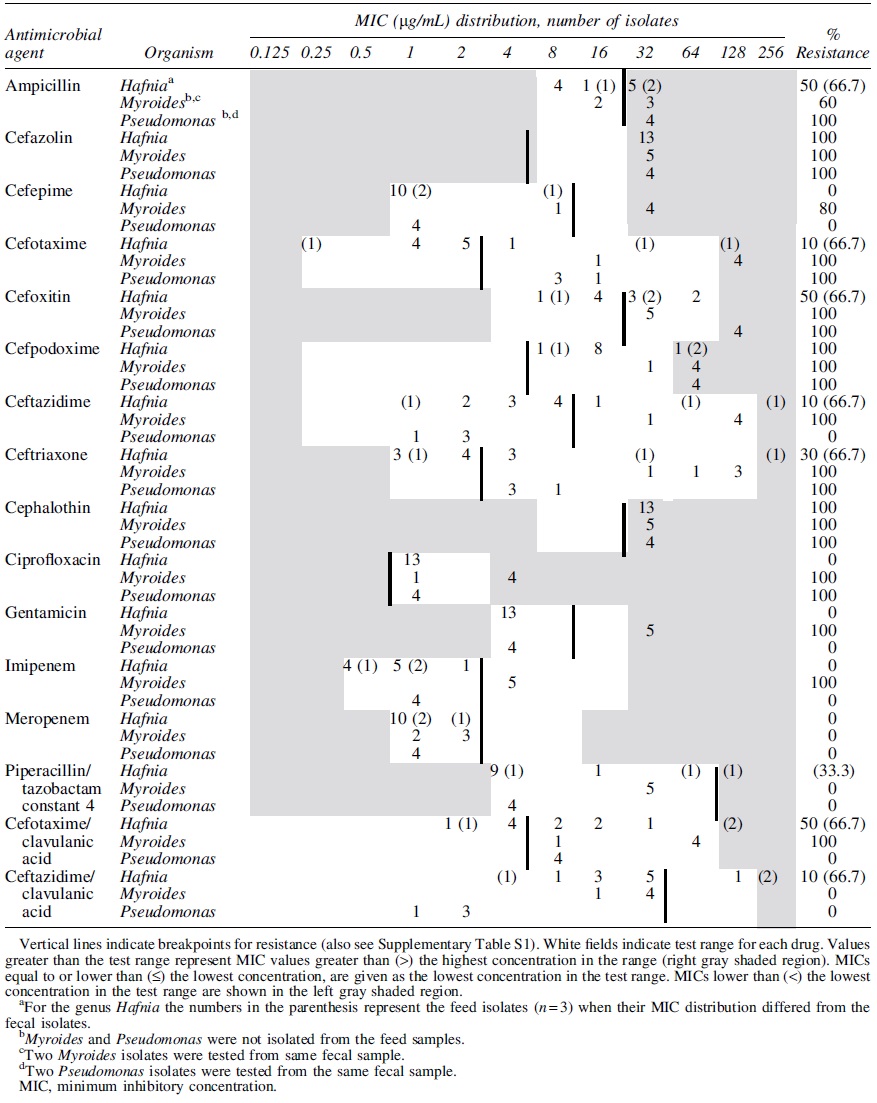
All ESBL E. coli isolates, both from feces and feed, were resistant to penicillin (ampicillin), and to most of the cephems (cefazolin, cefotaxime, cefpodoxime, ceftriaxone, and cephalothin). However, they were susceptible to carbapenems and β lactam/β lactamase inhibitor combination drugs (Tables 2 and 3). Co-resistance to antibiotic classes other than β-lactams was observed to ciprofloxacin (33%) and gentamicin (28%) among the fecal ESBLs E. coli isolates indicating MDR. All ESBLs fecal and feed K. pneumoniae isolates and all fecal ESBLs Proteus spp. were resistant to ampicillin, cefazolin, and cephalothin but they were all susceptible to carbapenems and β lactam/β lactamase inhibitor combination drugs (Tables 2 and 3). All or some of the three fecal ESBLs Citrobacter isolates were resistant to penicillin (ampicillin), most cephem (cefazolin, cefotaxime, cefoxitin, cefpodoxime, ceftazidime, ceftriaxone, and cephalothin) classes, and cefotaxime/clavulanic acid combination (Table 2). Feed ESBLs Enterobacter isolates were resistant to penicillin (ampicillin), most cephem (cefazolin, cefotaxime, cefoxitin, cefpodoxime, ceftriaxone, and cephalothin), and aminoglycoside (gentamicin) antimicrobial classes indicating MDR (Table 3). Although all CR bacteria were resistant to cefazolin, cefpodoxime, and cephalothin (cephems), they all were susceptible to ciprofloxacin, gentamicin, imipenem, and meropenem (Table 4).
Minimum Inhibitory Concentration Distribution of Extended-Spectrum β-Lactamase-Producing E. coli (n = 39), K. pneumonia (n = 32), and Proteus spp. (n = 37), Pseudomonas fulva (n = 2), Acinetobacter spp. (n = 2) and Citrobacter spp. (n = 3) Isolated from Mink Feces (n = 42) in the United States

Minimum Inhibitory Concentration Distribution of Extended-Spectrum β-Lactamase-Producing E. coli (n = 8), K. pneumonia (n = 6), and Enterobacter spp. (n = 2) Isolated from Mink Feed (n = 8) in the United States
Minimum Inhibitory Concentration Distribution of Carbapenem-Resistant Fecal (N = 10)-and Feed (N = 3)-Hafnia alvei Isolates, Fecal Myroides odoratimimus Isolates (N = 5), and Fecal Pseudomonas spp. (N = 4) Isolated from Mink Feces (N = 42) and Feed (N = 8) in the United States
Antibiotic resistance genes detected from ESBL-producing and CR bacteria
The distribution of the ESBLs genes detected from ESBL-producing bacteria is given in Table 5. All fecal ESBL E. coli isolates were positive for one of the bla CTX-M (CTX-M-15, CTX-M-2, or CTX-M-14) genes tested. Fecal ESBLs E. coli isolates that carried bla TEM were also positive for bla CTX-M-15. Feed E. coli carried either the CTX-M-14 or CTX-M-15. Although all fecal ESBL K. pneumoniae isolates were positive for bla SHV, feed isolates were positive for bla CTX-M-9. All ESBL Proteus spp., Acinetobacter spp., and Pseudomonas fulva isolates were positive for bla TEM gene. The carbapenemase genes we tested were not detected from any of phenotypically CR bacterial isolates.
Beta-Lactamase (bla) Genes Detected from Phenotypically Extended-Spectrum β-Lactamase-Positive Bacteria
All bla TEM positive isolates were also positive for bla CTX-M-15.
The two Pseudomonas isolates were obtained from same fecal sample.
na, not tested.
Discussion
The most prevalent ESBL-producing Enterobacteriaceae fecal isolates were E. coli, K. pneumoniae, and Proteus spp., although at low prevalence, Pseudomonas and Acinetobacter spp. that are important human pathogens were also detected from mink feces. Enterobacteriaceae are normal flora of the mammalian intestine, however they can also cause opportunistic infections. For example, bacterial genera isolated in this study such as E. coli, K. pneumoniae, and Pseudomonas spp., have been isolated from clinical samples as pathogens causing diseases in mink (Pedersen et al., 2009; Compo et al., 2017; Nikolaisen et al., 2017).
ESBL-producing bacteria detected both from the mink feces and feed samples carried bla CTX-M, bla SHV, or bla TEM genes. From same mink population using quantitative PCR array, we previously observed bla SHV and bla OXA in 17% and 29% of the metagenomic DNA samples, respectively (Agga et al., 2021). These genes (bla CTX-M, bla SHV, and bla OXA) were also reported from metagenomic DNA obtained from municipal wastewater and livestock manure (Agga et al., 2015), and from the feces of feedlot cattle (Vikram et al., 2017). We note that not all bla SHV, bla TEM, and bla OXA genes are ESBLs and identification of specific alleles requires sequencing (Pitout and Laupland, 2008), which was beyond the scope of this project. All fecal and feed E. coli isolates obtained in this study were positive for bla CTX-M gene. ESBLs genes are mostly plasmid borne; the bla CTX-M gene emerged as clinically significant replacing the SHV and TEM families (Bush, 2018; Lee et al., 2020). From the same mink population, we detected bla CTX-M in 35% of third-generation cephalosporin-resistant (TGCr) E. coli isolates (Agga et al., 2021). The bla CTX-M was detected in 51% of TGCr E. coli obtained from ceftiofur-treated cows as compared with 29% of the isolates obtained from nonceftiofur-treated cows (Agga et al., 2016a). Similarly, bla CTX-M was detected in E. coli from 33% of beef cattle, 86% of which received ceftiofur alone or in combination with tetracycline (Cottell et al., 2013). These studies clearly indicate that the use of TGCs selects for ESBLs. Studies in mink are rare; however, a recent study from China reported ESBLs E. coli carrying the bla CTX-M and/or bla TEM genes from five mink (Zheng et al., 2019). The lack of carbapenemase genes commonly reported in the literature among CR bacteria can be attributed to their intrinsic resistance to the carbapenems (Nikolaisen et al., 2017).
Animal feed can be a source of antimicrobial-resistant (AMR) bacteria in animal production particularly in mink farms whose feed is animal derived (Crump et al., 2002). ESBL E. coli and K. pneumoniae were detected from all or majority of the feed samples. We previously reported bacteria resistant to other antibiotics such as TGCs and tetracyclines from mink (Agga et al., 2021) and beef cattle feeds (Agga et al., 2016b). Although the number of feed samples analyzed was small, this study indicates the importance of animal feed as a source of foodborne pathogens and bacteria resistant to critically important antibiotics for human health.
Conclusions
Although this study was limited to a single large commercial mink farm owing to access, it is the first to report the occurrence of bacteria resistant to the highest priority critically important antibiotics in mink production. However, our findings highlight the need for more studies to better understand the epidemiology of bacteria resistant to critically important antibiotics for public health. Mink farmers can be exposed to ESBL-producing and CR bacteria through direct contact with the animals. A study from the Netherlands found that people who lived in close proximity to mink farms had higher ESBLs E. coli and K. pneumoniae carriage prevalence than those who lived farther away (Wielders et al., 2017). Improperly treated mink manure when used for soil amendment or through runoff from mink farms can disseminate these bacteria into the environment. The finding of ESBL bacteria in the mink feed indicates that animal feed can serve as important vehicle for the dissemination of AMR bacteria including the critically important antibiotics.
Footnotes
Acknowledgments
The authors thank Rohan Parekh, Anna Wilkin, and Maggie Mesker for the technical support.
Disclaimer
Mention of trade names or commercial products in this publication is solely for providing specific information and does not imply recommendation or endorsement by the U.S. Department of Agriculture. USDA is an equal opportunity provider and employer.
Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported by the U.S. Department of Agriculture, Agricultural Research Service (Project No. 5040-12630-006-00D).
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.