
Abstract
Antimicrobial resistance (AMR) is a major public health challenge and spreads through humans, animals, and the environment. Many reports show that AMR genes (ARGs) or phenotypes can be transferred from food animals to humans. However, the level and correlation of AMR in different nodes of the poultry meat supply chain are still poorly understood. Herein, 225 Escherichia coli isolates were recovered from chilled chicken samples from markets (123) and chicken fecal samples from farms (102) in Zhejiang Province, China. The dominant sequence types (STs) were ST155 (8.89%), ST48 (7.56%), and ST10 (7.11%), which are common in chicken and fecal samples. Antimicrobial susceptibility testing (AST) analysis showed that the E. coli isolates from fecal samples and retail chickens were resistant to ampicillin (61.77% and 63.42%, respectively) and trimethoprim (56.87% and 52.85%). Moreover, 36.59% of the E. coli isolates from chilled chickens and 39.22% of the isolates from fecal samples were resistant to three or more antimicrobial agents. A total of 59 ARGs were identified in sequenced E. coli genomes, including the mcr-1 gene involved in colistin resistance. The E. coli from farms and markets could be clustered in the same branch according to core single nucleotide polymorphisms. In addition, toxin genes astA and hlyE were also predicted in 86.5% (32/37) and 13.5% (5/37) of the above genomes, respectively. Taken together, these findings demonstrated that E. coli isolates from markets and farms showed similar AMR patterns, suggesting that E. coli strains in markets may originate from farms.
Introduction
Antibiotics have been used for decades in animal and poultry feed to promote growth and increase feed efficiency (Sugden et al., 2016; Wang et al., 2017a; Ministry of Agriculture and Rural Affairs, P.R. China, 2019). The widespread use of antibiotics in humans and animals is the main cause of the emergence and spread of antibiotic resistance (Sugden et al., 2016; Van Boeckel et al., 2017). Antimicrobial resistance (AMR) has become one of the most severe threats to human health worldwide and seriously impairs the treatment of life-threatening infections (Sun et al., 2016; National Academies of Sciences, Engineering, Medicine, 2017). Escherichia coli is a widespread member of the family Enterobacteriaceae that is mainly found in the environment and the intestinal tract of humans and animals. Although E. coli is generally a harmless intestinal commensal bacterium, certain pathogenic strains can cause various diseases in humans and animals (Kaper, 2004; Duffy et al., 2008).
E. coli is also an important reservoir of AMR genes (ARGs) and is often used as an indicator organism for resistance monitoring (Soufi et al., 2011; Poirel et al., 2018). ARGs have recently received increased research interest because they pose a serious threat to human health (Hammerum and Heuer, 2009). E. coli-borne ARGs can easily spread to other bacterial species through horizontal gene transfer (Lu et al., 2016; Poirel et al., 2018). The presence of antimicrobial-resistant E. coli in poultry is an important risk factor for human infection (Wu et al., 2014; Tong et al., 2015; Eltai et al., 2018). Although there have been many studies on E. coli contamination in chickens, few have performed detailed AMR comparisons of E. coli from different sources in the chicken supply chain.
In this study, E. coli isolates recovered from retail chicken and chicken fecal samples from farms in Zhejiang Province, China, between June 2016 and November 2017 were analyzed using antimicrobial susceptibility testing (AST) and multilocus sequence typing (MLST). Randomly selected isolates were subjected to whole-genome sequencing and analyzed for ARGs, potential virulence factors, and plasmid replicon types.
Materials and Methods
Isolate information
Between June 2016 and November 2017, 150 chilled chicken samples were collected from 15 supermarkets and 29 farmers' markets in the following 6 cities in northern Zhejiang Province: Ningbo, Hangzhou, Jinhua, Shaoxing, Huzhou, and Jiaxing. In addition, 120 broiler chicken fecal samples were collected from 7 layer farms and 5 broiler farms in the following 7 regions: Ningbo, Hangzhou, Jinhua, Quzhou, Taizhou, and Wenzhou (Fig. 1A). Five to ten whole chilled chicken carcasses from each supermarket were bought and were put in separate sterile plastic bag. Fecal samples were collected from fresh feces with sterile cotton swabs.
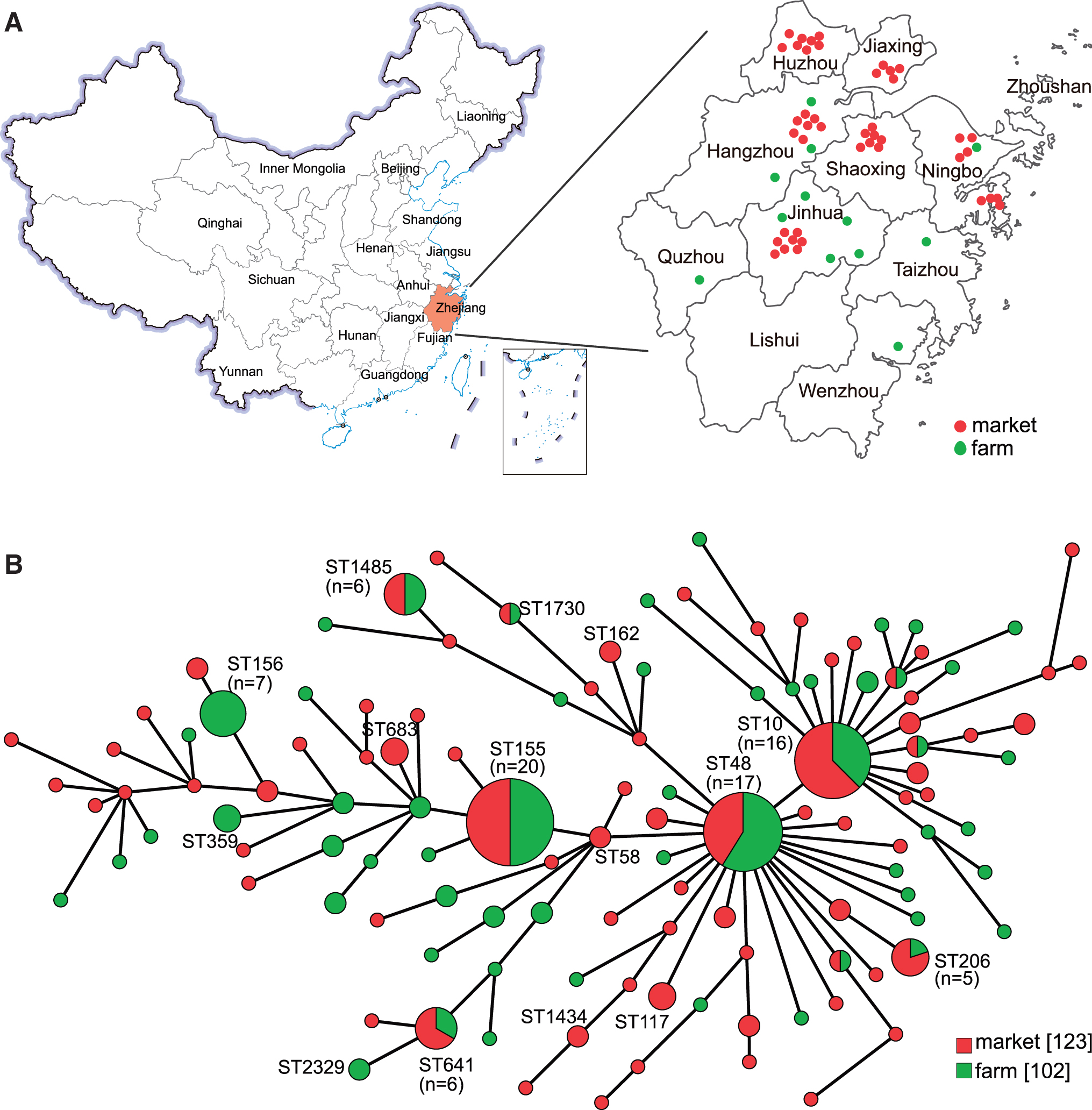
Map of sampling areas in Zhejiang Province, China, and minimum spanning tree of Escherichia coli strains based on MLST.
All fecal and chilled chicken samples were kept in the dark and transported to the laboratory at 2–8°C. The fecal material was suspended in 0.9% (w/v) sterile NaCl (physiological salinity), spread on Petri dishes containing MacConkey agar (Oxoid Ltd, Basingstoke, United Kingdom), and incubated at 37°C for 18–24 h. Chilled chicken was mixed with buffered peptone water (Oxoid Ltd). The suspension was cultured at 37°C for 12 h and then streaked on MacConkey agar as described previously. Only one clone was selected from each sample. All isolate identities were confirmed by Matrix-Assisted Laser Desorption/Ionization Time of Flight Mass Spectrometry (MALDI-TOF MS) (Kostrzewa, 2018) and polymerase chain reaction (PCR), and the isolates were stored at −80°C in 2 mL sterile tubes containing Luria-Bertani medium and 20% glycerol for further experiments.
Antimicrobial susceptibility testing
All E. coli isolates were characterized by AST using the Vitek®-2 Compact system (BioMérieux, France) (Simgamsetty et al., 2016). The panel of antimicrobial compounds tested included ampicillin (AMP), amoxicillin (AMC), piperacillin (TZP), cefazolin (CFZ), cefoxitin (FOX), ceftriaxone (CRO), cefepime (FEP), aztreonam (ATM), ertapenem (ETP), imipenem (IMP), amikacin (AMK), gentamicin (GEN), tobramycin (TOB), ciprofloxacin (CIP), levofloxacin (LEV), tigecycline (TGC), nitrofurantoin (NIT), and trimethoprim (SXT), as well as testing for extended-spectrum β-lactamase (ESBL). Colistin susceptibility testing was performed by the broth microdilution method using the Gram-negative panel (Tang et al., 2020). The minimum inhibitory concentration results were interpreted according to the Clinical and Laboratory Standards Institute (CLSI) documents M100-S26. Especially, the breakpoint of TGC for E. coli was interpreted according to the Food and Drug Administration criteria (resistant, ≥8 mg/L).
MLST analysis
DNA was extracted from E. coli isolates using a commercially available bacterial genomic DNA isolation kit (Generay, Shanghai, China) according to the manufacturer's instructions. Primers for seven housekeeping genes (adk, fumC, gyrB, icd, mdh, purA, and recA) were designed as previously described (Wirth et al., 2006) (Supplementary Table S1). The following reaction mixture was used for PCR: 15 μL of premixed Taq (TaKaRa), 0.9 μL of forward primer, 0.9 μL of reverse primer, 13 μL of sterile water, and 0.2 μL of template DNA. The following program was used for PCR amplification: an initial denaturation at 94°C for 4 min; 31 cycles of 98°C for 10 s, 59°C for 20 s, and 72°C for 40 s; and extension at 72°C for 1 min. Amplification products were sequenced by Sanger sequencing using a 3730XL instrument. Finally, MLST was carried out as previously described using MLST 2.0 software (
Whole-genome sequencing and identification of ARGs, virulence factors, and plasmid replicon types
One to two strains were randomly selected from each farm or market source for sequencing. An Illumina sequencing library was generated using a NEXTflex DNA sequencing kit (Bioo Scientific). Genomes were obtained using Illumina sequencing by constructing two paired-end libraries with average insertion lengths of 500–2000 bp. Reads were generated using an Illumina HiSeq 4000 and assembled into contigs and scaffolds using Velvet, with an average depth of coverage of 200 × . All paired-end reads (2 × 150 bp) were checked for quality and trimmed with Trimmomatic v0.36. All low-quality (Q < 20) data were filtered out. ARGs were predicted using ResFinder 4.0 software (
Results
MLST analysis of E. coli isolates
In total, 225 E. coli strains were recovered and confirmed by MALDI-TOF MS and PCR, including 102 from 120 fecal samples and 123 from 150 chilled chicken samples. All E. coli isolates were characterized using MLST analysis by partial sequencing of seven housekeeping genes. A total of 207 sequence types (STs) were identified and 18 different STs were found (Supplementary Table S2). ST155 (9%) was the most common ST, followed by ST48 (8.1%) and ST10 (7.2%) (Fig. 1B). At the same time, some isolates with the same AMR phenotype were assigned to multiple STs.
AMR patterns
E. coli isolated from fecal and chicken samples were most highly resistant to AMP, with resistance rates of 61.77% and 63.42%, respectively, followed by SXT, with resistance rates of 56.87% and 52.85%, respectively. TZP resistance and ETP resistance were detected only in isolates from fecal samples. No E. coli isolates were resistant to TGC.
A total of 42 AMR profiles were identified in the 123 E. coli isolates from chilled chicken samples from markets. Of these, 92 isolates (74.80%) were resistant to at least one antimicrobial agent, whereas 31 isolates (25.20%) were susceptible to all tested antimicrobials. The main AMR profile was AMP-SXT (19 isolates, 15.45%). Forty-five isolates (36.59%) were multidrug resistant (Supplementary Table S3); 26.82% isolates were ESBL positive (33 of 123). On the contrary, 32 AMR profiles were detected among the 102 isolates from broiler chicken fecal samples collected at farms. A total of 70 of these isolates (68.63%) were resistant to at least one antibacterial agent, and 40 isolates (39.22%) were resistant to at least 3 antibiotics. The main AMR profile was AMP-SXT (17 isolates, 16.67%), followed by AMP (7 isolates, 6.87%) (Supplementary Table S4). Moreover, 19.61% (20 of 102) of the chicken farm isolates were ESBL positive. In summary, the AMR rates were similar in the strains from both sources (Fig. 2), except for those for several antibiotics (quinolones: CIP and LEV; cephalosporins: CFZ, FOX, and FEP).
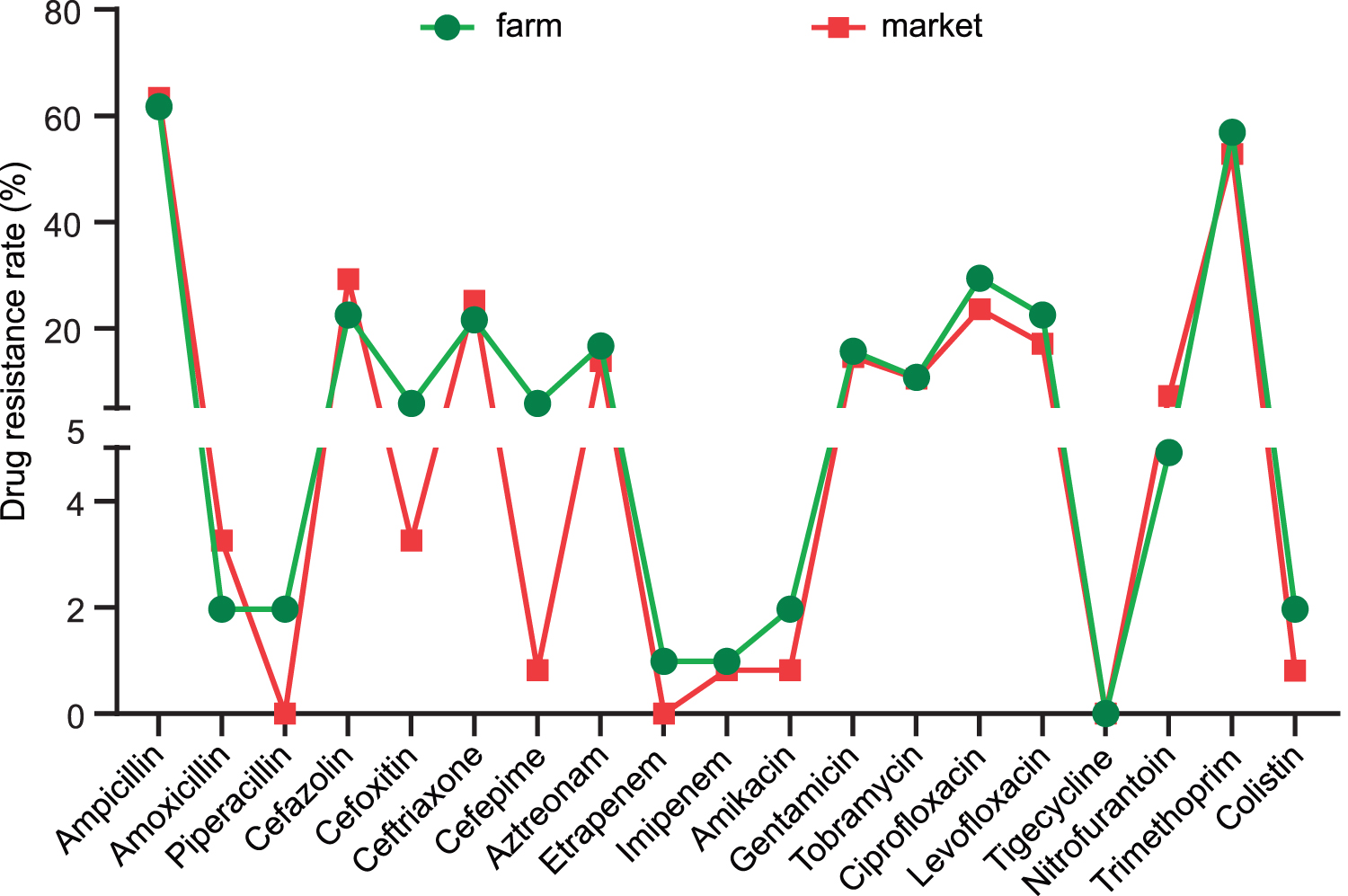
AMR rates of Escherichia coli strains isolated from markets and farms. AMR, antimicrobial resistance. Color images are available online.
Whole-genome sequencing and ARGs
A total of 37 E. coli strains from different sampling locations were randomly selected (14 isolates from chicken and 23 isolates from fecal samples) for further analysis. The whole genome sequences of these strains were obtained and examined for ARGs. A total of 59 ARGs were detected from 34 drug-resistant strains (Fig. 3). ARGs associated with aminoglycosides were most common, with 18 genes detected. In addition, 13 β-lactam resistance genes, 6 fluoroquinolone resistance genes, 1 fosfomycin resistance gene, 5 SXT resistance genes, 4 phenicol resistance genes, 2 rifampicin resistance genes, and 3 each of macrolide–lincosamide–streptogramin B (MLS), sulfonamide, and tetracycline resistance genes were detected. No fusidic acid, glycopeptide, nitroimidazole, or oxazolidinone resistance genes were detected in any isolate. Moreover, strains CMNB20b and CFRA112 were predicted to carry the mcr-1 gene.

Acquired resistance genes and phylogenetic SNP tree of Escherichia coli strains with whole genome sequences.
E. coli from farms and markets could be clustered in the same branch according to core single nucleotide polymorphisms (SNPs), indicating that some of the bacteria in markets might come from farms (Fig. 3). For example, strains CFYH060, CMJH107b, CFRA108, and CMJH97b were clustered in one branch, CFRA112, CFRA109, and CMJH104a were clustered in another cluster, and so on (Fig. 3B). In addition, a total of 23 plasmid replicon types were detected. The most common plasmid replicon type was IncFIB (AP001918), followed by p0111 and IncFIC (FII) (Supplementary Table S5).
Virulence factor genes of E. coli isolates
Virulence factors involved in this study were divided into five categories: adherence, invasion, iron uptake, secretion systems, and toxins. The most common types of virulence factors among the 37 strains were adhesion and invasion factors. Seventeen isolates were predicted to include a type 6 secretion system (T6SS; more than 14 of the 20 genes could be predicted). Seven astA genes (encoding EAST-1 heat-stable toxin) were found in strains CMNB12b, CMHUZ85a, CFQJ047, CFQJ049, CFQJ051, CFYH075, and CFFY125, which is a typical feature of enteroaggregative E. coli (Boisen et al., 2020). Moreover, hlyE, encoding a hemolysin, was predicted in 32 of 37 strains. hlyA and cysC1 were predicted in 2 isolates (Supplementary Table S6).
Discussion
E. coli can spread from the intestines to animal carcasses and other chicken products during the slaughter and processing of chickens, circulate through the poultry industry chain, and pose many threats (Canizalez-Roman et al., 2013; Park et al., 2019; Sary et al., 2019; Li et al., 2020). We examined the AMR, MLST, and virulence factors of E. coli isolated from retail chicken and chicken fecal samples from farms in a province in China. The results of this study illustrated that E. coli is common in the chicken industry chain and that the AMR profiles of E. coli isolates from chilled chicken at markets are similar to those of isolates from broiler chicken fecal samples from farms.
ARGs of E. coli can be transferred horizontally by mobile elements, such as plasmids, phages, and integrons, leading to the acquisition of drug resistance by other bacteria in the environment (Von Wintersdorff et al., 2016; Partridge et al., 2018; Rozwandowicz et al., 2018). Thus, AMR of E. coli strains has become more serious owing to the widespread use of antibiotics as feed additives in China (He et al., 2019). In China, ∼50,000 tons of antibacterial agents were applied in food animals, and more than half of them were used for growth promotion (Zhang et al., 2015; Ministry of Agriculture and Rural Affairs, P.R. China, 2019). According to previous reports, for each ton food animals are produced, 318 and 140 g antibiotics are consumed in 2013 (Zhang et al., 2015) and 2018 (Ministry of Agriculture and Rural Affairs, P.R. China, 2019), respectively. β-lactams, macrolides, tetracyclines, phenicols, aminoglycosides, and fluoroquinolones were the most frequently used veterinary antibiotics in farms. The level of AMR of zoonotic bacteria is related to the use of drugs in livestock and poultry breeding (Zhang et al., 2015; Wang et al., 2020a). The level of resistance to drugs that are not used in animal farms, such as TGC and IMP, was low (Wang et al., 2017b; Sun et al., 2019). In contrast, E. coli has a high level of resistance to aminoglycosides and β-lactams. In this study, the resistance to aminoglycosides (GEN and TOB), quinolones (CIP and LEV), and cephalosporins (CFZ and CRO) was 10.6–29.4%. The proportions of multidrug resistance of isolates from the two sources were very similar (36.59 vs. 39.22%), and the main AMR patterns were the same, with resistance to AMP-SXT and AMP. Surprisingly, the isolation rate of colistin-resistant strains was 1.3%, which was lower than that reported in 2016–2017 in China (Wang et al., 2017a, 2018, 2020b), reflecting the diversity of livestock and poultry breeding in China. The antibiotic resistance of E. coli from farms is slightly higher than that of E. coli from supermarkets, which is similar to previous report (Lei et al., 2010). TZP and ETP resistance was detected only in samples from farms. In detail, more E. coli isolates from farms were resistant to FOX, FEP, CIP, and LEV. In general, AMR phenotypes were similar between E. coli isolates from markets and farms, and the overall AMR is similar to that in other areas of China (Zhang et al., 2017). Combined with the SNP phylogenetic analysis, it is easy to speculate that the bacterial resistance of isolates from supermarkets comes from farms.
In this study, the acquired ARGs of the strains were consistent with their AMR phenotypes, and no acquired ARGs were detected in three nonresistant strains. Nevertheless, more genome sequencing is the next important plan to predict ARGs, which can well explain or even predict AMR phenotypes. E. coli isolates in this study contained many molecular polymorphisms. MLST analysis revealed that the most common types were ST155, ST10, and ST48, which was consistent with a previous report (ST155: 8/110, ST48: 8/110, ST10: 6/110) (Aworh et al., 2021). In addition, Hoek et al. suggested that ST155 and ST10 were most often found in broiler farms (Van Hoek et al., 2018). Not surprisingly, ST155 was the most common in several reports (Foster-Nyarko et al., 2021; Nguyen et al., 2021). In addition, seven ST156 strains were found only on farms, and they may be hosts for the transmission of the carbapenem resistance gene bla NDM, as previously reported (Tang et al., 2019). Nonetheless, the correlation between ST and the AMR phenotype needs to be further explored.
Conclusions
This study analyzed the AMR of E. coli from different sources and provided support for E. coli traceability in the chicken industry chain. Our findings suggest that bacterial resistance monitoring should be emphasized, especially to control the drug resistance of bacteria in breeding.
Accession Numbers
The whole genome sequences of 37 strains have been deposited in the GenBank database under Accession Numbers RDCM00000000–RDDW00000000.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Natural Science Foundation for the Youth (31700007), State Key Laboratory for Managing Biotic and Chemical Threats to the Quality and Safety of Agro-products (2010DS700124-ZZ2008), and Key Research and Development Program of Zhejiang Province (2020C02031).
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
Supplementary Table S5
Supplementary Table S6
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.