
Abstract
In recent years, methicillin-resistant Staphylococcus aureus (MRSA) has gained interest in veterinary medicine due to its zoonotic potential. Currently, little information is available on the genotypic and virulence characteristics of MRSA isolates detected in Nigerian abattoirs. To better understand the epidemiology of MRSA associated with the abattoir food chain environment in Nigeria, a total of 18 isolates (humans: n = 5, slaughter animals: n = 5, and environment: n = 8), previously spa typed, were recovered and characterized by Staphylococcus cassette chromosome mec (SCCmec) typing, and phenotypic and genotypic antimicrobial susceptibility testing. In addition, 10 of the 18 MRSA strains with a new spa type (t16571) were subjected to multilocus sequence typing. The similarity of strains was analyzed based on the results of the DNA microarray analysis. The 18 MRSA strains harbored two distinct SCCmec types (IVa and V) and belonged to four clonal clusters (CC1, CC7, CC88, and CC152). All MRSA of the new spa type t16571 (n = 10) harbored the SCCmec type IVa. Seven of the MRSA t16571 strains belonged to ST88, while three other strains were assigned to ST3614. The 18 MRSA isolates were categorized into six virulence profiles, and the detection rate for the Panton–Valentine Leukocidin gene was high (33.3%). The antimicrobial susceptibility profiles of the 18 MRSA varied widely between strains, but phenotypic resistance corresponded to relevant resistance genes harbored. The detection of highly similar MRSA strains in slaughter animals, abattoir workers, and the environment underlines the need to use adequate measures at Nigerian abattoirs to prevent further spread and transmission of MRSA to humans or food.
Introduction
Recently, the epidemiology of Staphylococcus aureus in animals has received significant interest due to the emergence of uncommon clonal lineages of methicillin-resistant S. aureus (MRSA) strains (Lozano et al., 2016). In particular, the frequent detection of livestock-associated (LA-) MRSA lineages (primarily CC398 MRSA) is a matter of concern. LA-MRSA is considered zoonotic, and it often colonizes and infects people who have occupational contact with livestock, such as slaughterhouse workers (van Loo et al., 2007). Hence, food contamination with MRSA could occur at any point during slaughter favoring a potential spread to consumers.
In Nigeria, several studies have reported the circulating lineages of MRSA in clinical settings (Ghebremedhin et al., 2009). Also, S. aureus and MRSA have been detected in Nigerian abattoirs (Odetokun et al., 2018). However, there is currently limited information on the MRSA lineages circulating in slaughterhouses or their molecular characteristics and virulence profiles. For instance, there is lack of information on the presence/absence of the Panton–Valentine Leukocidin (PVL), which plays an important role in the virulence of S. aureus. Nonetheless, since the first report of a PVL-positive MRSA ST88 in a Nigerian clinical setting (Ghebremedhin et al., 2009), there are concerns about the distribution of MRSA in the Nigerian community and food processing environment.
The purpose of this study was to investigate the clonality, virulence-associated characteristics, as well as the phenotypic and genotypic antimicrobial features of a small collection of MRSA isolated from food animals, abattoir workers, and the food processing environment in Nigeria. In addition, the similarity of strains was analyzed to identify possible spread and transmission events between slaughter animals, humans, and the environment, respectively.
Materials and Methods
S. aureus strains
A total of 1671 samples were collected from two municipal abattoirs in Nigeria over a 6-month period by randomly swabbing nasal passages of human abattoir workers at various processing steps (n = 260) and food animals (cattle: n = 500, goat: n = 400, and pigs: n = 81) before slaughter. In addition, samples were taken from various locations in the abattoir environment (i.e., cutting tables and slaughterhouse walls: n = 430). The study protocol was approved by the Ethical Review Committee of the Ministry of Health, Oyo State (AD13/479/1079) and the University of Ibadan Animal Care and Use Committee (UI-ACUREC/App/2016/018). Overall, 109 S. aureus were isolated from the 1671 samples (detection rate = 5.4%), with 18 MRSA (human origin: n = 5, slaughter animals: n = 5, and environment: n = 8) (Fig. 1). Of these 18 MRSA strains, ten isolates belonged to the novel spa type t16571 (Odetokun et al., 2018).
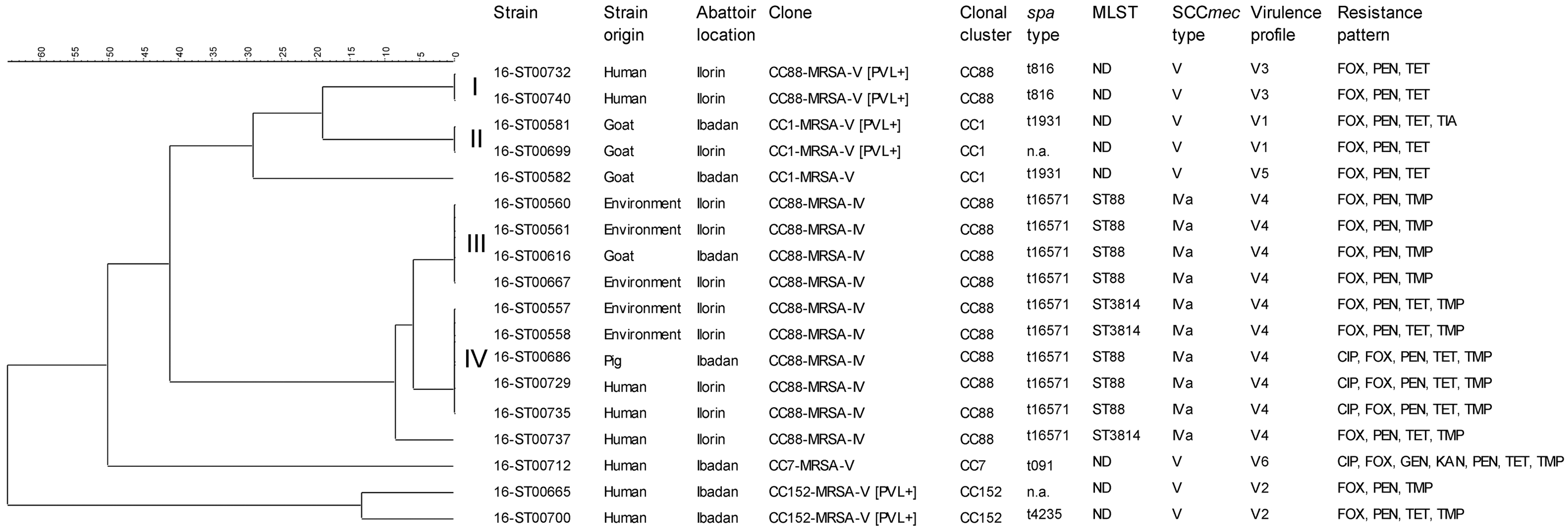
Dendrogram based on similarity between the array hybridization profiles of all 18 MRSA isolates using the Jaccard coefficient and UPMGA. Clades are indicated with I–IV. The virulence profiles are described in Table 1. CIP, ciprofloxacin; FOX, cefoxitin; GEN, gentamycin; KAN, kanamycin; MRSA, methicillin-resistant Staphylococcus aureus; MLST, multilocus sequence typing; n.a., not available; ND, not determined; PEN, penicillin; TET, tetracycline; TIA, tiamulin; TMP, trimethoprim; UPMGA, unweighted-pair group method with arithmetic averages.
Genotypic characterization
SCCmec typing was performed on all MRSA isolates using the multiplex-PCR-based method described by Zhang et al. (2005). Subcultures of the original control strains used for the SCCmec typing method developed by Zhang et al. (2005) were used as controls. Further multilocus sequence typing was performed on 10 MRSA isolates with the new spa type t16571 according to Enright et al. (2000), since their sequence types could not be inferred from the spa type.
DNA microarray analysis
All 18 MRSA strains were differentiated further as described earlier (Fetsch et al., 2016) using a highly discriminatory commercially available microarray kit (Identibac S. aureus Genotyping kit; Alere Technologies GmbH, Jena, Germany) covering 333 target sequences for species-specific markers, virulence genes, and resistance determinants. Furthermore, the hybridization patterns can be used to assign isolates to clonal complexes (CCs) (Monecke et al., 2008). For similarity analysis, a binary cluster analysis using a Jaccard correlation of the microarray data was carried out with the BioNumerics Software (Version 7.6.2).
Antimicrobial susceptibility testing
Minimum inhibition concentrations (MICs) of 19 antibiotics (cefoxitin: FOX, ciprofloxacin: CIP, clindamycin: CLI, chloramphenicol: CHL, erythromycin: ERY, fusidate: FUS, gentamicin: GEN, kanamycin: KAN, linezolid: LZD, mupirocin: MUP, penicillin: PEN, quinupristin/dalfopristin: SYN, rifampin: RIF, streptomycin: STR, sulfamethoxazole: SMX, tetracycline: TET, tiamulin: TIA, trimethoprim: TMP, and vancomycin: VAN) tested were determined by broth microdilution according to the European Committee on Antimicrobial Susceptibility Testing (EUCAST) protocol version 6.1 (
Results
Typing of isolates
The 18 MRSA strains belonged to four CCs (CC88: n = 12; CC1: n = 3; CC152: n = 2; and CC7: n = 1) and were found to carry the mecA gene on two distinct SCCmec types (IVa and V) (Fig. 1). All ten MRSA of spa type t16571 harbored the SCCmec type IVa belonging to ST88 (n = 7) and a novel ST3814 (n = 3). Eight MRSA isolates (spa types t091, t1931, t816, t4235, and two spa nontypeable isolates) harbored the SCCmec type V.
Microarray-based genotyping
The 18 MRSA isolates were categorized into six virulence profiles (V1–6; Table 1). The PVL gene was detected in 6/18 (33.3%) of the isolates (V1, V2, and V3). Other leukocidin encoding genes (lukF, lukS, lukD, lukE, lukX, lukY, and hlgA) were common.
The toxic shock syndrome toxin (tst), exfoliatins (etA/B/D), the epidermal cell differentiation inhibitors (edinA/B/C), the arginine catabolic mobile element (ACME locus), and the egc-cluster (seg, sei, sem, sen, seo, and seu) were not identified in any of the MRSA isolates. However, genes encoding staphylococcal enterotoxin A (sea) and C (sec) were found in strains categorized into V3 and V5, respectively; in addition, V3 strains also carry the staphylococcal enterotoxin L (sel) (Table 1).
Virulence Profiles of the 18 MRSA on the Basis of Selected Virulence Determinants
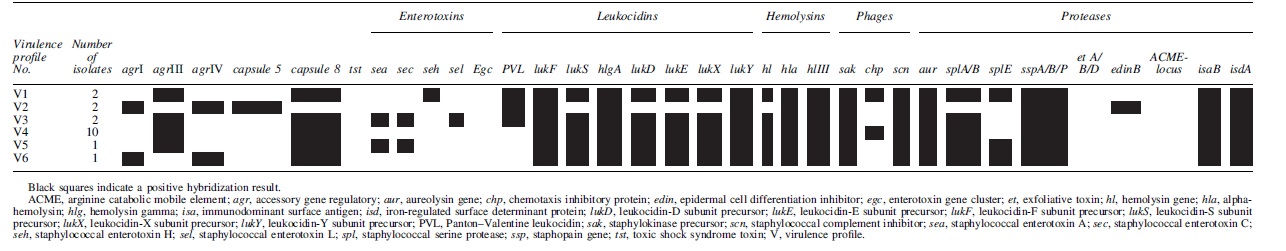
The epidermal cell differentiation inhibitor (edinB) was found only in isolates classified as V2. In contrast, the beta-hemolysin converting phage carrying the immune evasion cluster (IEC) was detected in all the strains. However, only isolates categorized as V1 and V4 carried all of the three genes (sak, scn, and chp). More differences among the strains in the six virulence profiles were seen as regard to the capsule type and the accessory gene regulator allele (Table 1).
For further details on the results of the microarray-based genotyping, see Supplementary Table S1.
Similarity analysis
This study also focused on the identification of possible spread and transmission events between slaughter animals, humans, and the environment. Therefore, a similarity analysis was conducted based on all hybridization results of the 18 MRSA strains obtained with the microarray; the outcome is presented in the dendrogram (Fig. 1). Several clades of strains displaying a high similarity were identified: clade (1) with two CC88-MRSA-V-(PVL+) strains, both isolated from the abattoir environment; clade (2), a CC1-MRSA-V-(PVL+) strain isolated from a goat was highly similar to a strain found in the slaughter environment; clade (3) consists of four CC88-MRSA-IV strains, three of human origin and one isolated from a goat; and clade (4) with five CC88-MRSA-IV strains, two of human origin, two isolated from environmental samples, and one from a pig.
Phenotypic and genotypic characterization of antimicrobial resistance
The susceptibility of the 18 MRSA to antimicrobials was tested by broth microdilution. In addition, the microarray was used to determine the presence of designated resistance determinants. All 18 isolates demonstrated 100% phenotypic resistance to β-lactam antibiotics (cefoxitin and penicillin). These strains also showed high phenotypic resistance to tetracycline and trimethoprim, respectively (72.7%, each), and low resistance to gentamicin, kanamycin, and tiamulin (5.6%). Phenotypic resistance of 22.2% was found among the strains against ciprofloxacin. Overall, six different phenotypic resistance patterns were exhibited by the MRSA isolates (Fig. 1). Phenotypic resistance patterns FOX-PEN-TMP (n = 5), FOX-PEN-TET (n = 4), and CIP-FOX-PEN-TET-TMP (n = 3) were commonly found among isolates from goats, slaughterhouse walls, and abattoir workers, respectively.
At the gene level, blaZ (the blaI and blaR operons) was commonly detected in the MRSA isolates of the CC88 as well as in the other five PVL-positive MRSA isolates. Other antimicrobial resistance genes detected include the tetK, dfrS1, aacA-aphD, aphA3, and sat resistance genes.
Discussion
This study presents the first report of MRSA strains with novel ST3614 (CC88) and PVL-positive MRSA of CC88 and CC152 in Nigeria. The ST88/CC88 MRSA has been named the “African clone” as it predominates in west, central, and east Africa among clinical human isolates (Schaumburg et al., 2014), with limited reports from the abattoir setting in Nigeria. Also, in this study, all human isolates belonged to the CC88. Nevertheless, MRSA CC88 with spa types that are different from those found in this study has been reported in pigs and sheep (Fall et al., 2012). Similar to the findings of this study, Okorie-Kanu et al. (2020) also reported that PVL-positive MRSA of CC1 strain was isolated from food animals and associated products in Nigeria. CC7-MRSA-V of spa type t091 was previously identified in human patients in Kenya (Omuse et al., 2016). The same spa type was also detected as methicillin-sensitive S. aureus in broilers and poultry farm workers (Vandendriessche et al., 2014) and cattle (Bergonier et al., 2014) in Europe. CC398 remains the most common type of LA-MRSA in most European countries and other parts of the world excluding Africa. This could be due to the variation in the epidemiology of LA S. aureus across geographic regions (Smith, 2015).
The 18 MRSA strains of our study were categorized into six virulence profiles based on the presence/absence of some important virulence determinants (i.e., enterotoxins, leukocidins, hemolysins, phages, and proteases). Overall, the leukocidins, hemolysins, phages, and proteases were common among the MRSA isolates of this study, while enterotoxin encoding genes were less often detected.
Leukocidin encoding genes are usually responsible for the lysis of blood cells during S. aureus pathogenesis (du Toit, 2019). The detection rate for PVL in MRSA in this study is comparable with the findings of Okorie-Kanu et al. (2020), who recorded rates of 20.0% and 36.4% PVL-positive MRSA in slaughter pigs and poultry, respectively, in Nigeria. However, Fall et al. (2012) recorded a higher detection rate (58.9%) of PVL-positive MRSA among pigs and livestock farmers in Senegal.
sea, sec, seh, and sel were the only enterotoxin encoding genes found among the 18 MRSA strains. The enterotoxins SEC and SEL are associated with bovine mastitis and skin and soft tissue infections (Saiman et al., 2003; Fang et al., 2019), while SEA is one of the five classical enterotoxins most frequently involved in staphylococcal food poisoning (Argudín et al., 2010). Among the protease genes found in the study strains, edinB facilitates the translocation of S. aureus into the bloodstream during pneumonia (Courjon et al., 2015). The phage-associated sak, scn, and chp genes belong to the IEC of S. aureus (van Wamel et al., 2006).
The results of the similarity analysis provide further evidence that spread and transmission events of pathogenic MRSA strains can occur in the slaughter environment in Nigeria. This may also have an impact on the food safety of Nigerian meat and meat products. It has already been shown in an earlier Nigerian study that ST88 MRSA isolated from pigs and humans displays a high genetic similarity (Otalu et al., 2018). Furthermore, it has been shown that MRSA has the potential to spread along the food chain to the human population in Nigeria (Okorie-Kanu et al., 2020).
The 100% phenotypic resistance to cefoxitin and penicillin is expected, while the high level of resistance to tetracycline and trimethoprim (found in 72% of the study isolates) is a concern. Tetracycline is a common drug and is one of the most misused antibiotics in human and veterinary practice in Nigeria (Alhaji and Isola, 2018; Oloso et al., 2018; WHO, 2018). The high phenotypic resistance to ciprofloxacin (found in 22% of the study isolates) is a public health concern. Ciprofloxacin, a fluoroquinolone, is considered to be a critically important therapeutic antimicrobial agent in humans (WHO, 2019). Overall, the MRSA in our study exhibited six distinct phenotypic resistance patterns, indicating the circulation of multidrug-resistant MRSA. This can aggravate the occurrence of infection and under this setting promote the persistence of antimicrobial resistance.
The CC88 has been found to usually carry the blaZ gene (Monecke et al., 2011). The blaZ gene alongside blaI and blaR operons, as detected in MRSA isolates in this study, is responsible for the observed resistance to penicillin/ampicillin in MRSA. Other studies have also documented the detection of the blaZ gene in MRSA strains from cattle and pigs (Feltrin et al., 2015), dairy sheep, and in-contact humans (Carfora et al., 2016). The detection of tetK, dfrS1, aacA-aphD, aphA3, and sat resistance genes may indicate widespread use of antibiotics such as tetracycline, trimethoprim, gentamicin, tobramycin, neomycin, kanamycin, and streptothricin in Nigeria.
Conclusions
As far as we know, this is one of the foremost studies describing the genotypic and virulence characteristics of MRSA isolated at Nigerian abattoirs. In addition, several clades of highly identical isolates were identified indicating a potential spread and transmission among slaughter animals, humans, and the food processing environment. We present the first report of abattoir MRSA strains with a novel ST3614(CC88) and PVL-positive MRSA isolates belonging to the CC88, and CC152 clonal lineages in Nigeria. Detection of MRSA, especially of PVL-positive strains, from slaughterhouse workers and slaughter animals is a public health concern. Contamination of meat with these pathogens at any point in the food chain could result in the spread of multiresistant MRSA to consumers. Therefore, suitable hygienic measures, including personal hygiene when handling food, are needed at all processing steps of the Nigerian meat chain.
Footnotes
Authors' Contributions
I.A.O.: Conceptualization, formal analysis, methodology, investigation, roles/writing—original draft, and writing—review and editing. S.M.: Formal analysis, methodology, and writing—review and editing. V.O.A.: Conceptualization, methodology, supervision, and writing—review and editing. A.F.: Formal analysis, resources, methodology, project administration, supervision, roles/writing—original draft, and writing—review and editing.
Acknowledgments
This work was carried out under the guest scientist program of the German Federal Institute for Risk Assessment (BfR). We acknowledge the excellent technical supports from B. Ballhausen and D. Leeser.
Disclosure Statement
We declare no conflict of interest.
Funding Information
Part of this work was funded under the Guest Scientist Program of the German Federal Institute for Risk Assessment (BfR), Berlin.
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.