
Abstract
Shigella flexneri, a common Gram-negative foodborne pathogen, is widely distributed in fresh-cut fruits and vegetables, unpasteurized milk, and food processing environments. The aims of this study were to evaluate the antibacterial effects of 405-nm light-emitting diode (LED) treatment on S. flexneri and to investigate the possible mechanism. The results showed that LED irradiation (360 min) reduced the number of S. flexneri in phosphate-buffered saline by 3.29 log colony-forming unit (CFU)/mL (initial bacterial count: 6.81 log CFU/mL). The cells in reconstituted infant formula, cells on fresh-cut carrot slices, and biofilm-associated cells on stainless steel surfaces were reduced by 1.83 log CFU/mL, 7.00 log CFU/cm2, and 4.35 log CFU/cm2 following LED treatment for 360, 120, and 120 min, respectively. LED treatment damaged both DNA and cell wall of S. flexneri and changed cell morphology and cell membrane permeability. In addition, LED treatment decreased total cell protein concentration of S. flexneri. These results indicated that 405-nm LED treatment effectively controlled S. flexneri contamination of foods and food contact surfaces and that the bacterial inactivation may be the result of damage to multiple cellular components. These findings highlight the potential of LED technology in controlling S. flexneri during food processing, storage, and preparation.
Introduction
Shigella is a genus of Gram-negative, rod-shaped, non-spore-forming, nonmotile, and facultatively anaerobic bacterial pathogen (Kang et al., 2018). Shigella spp. are mainly transmitted via the fecal–oral route and thus continue to pose a major threat to public health in developing countries with poor sanitation (Fan et al., 2015). Acute intestinal infections caused by Shigella are characterized by diarrhea, often accompanied by bleeding (Ahmed and Shimamoto, 2015). Among the estimated 165 million infection cases each year, resulting in 1.1 million deaths, 99% occur in developing countries (Azmi et al., 2014). The genus Shigella comprises four species: Shigella dysenteriae, Shigella flexneri, Shigella boydii, and Shigella sonnei, with S. flexneri being the most commonly isolated species (Kang et al., 2018).
S. flexneri has a low infectious dose, 10–100 bacteria are sufficient to cause infection (Ranganathan et al., 2019), and is transmitted via fresh fruits and vegetables, raw oysters, cooked meats, unpasteurized milk, and processed foods (Ahmed and Shimamoto, 2015). S. flexneri also has the ability to form biofilms, which help S. flexneri cells to resist various food processing conditions, including low temperature, low pH, and high salinity. The formation of biofilm leads to its constant colonization on the surface of food processing utensils and increases the risk of food contamination (Kang et al., 2018).
At present, S. flexneri control methods can be divided into two main categories: thermal sterilization and nonthermal sterilization. Thermal sterilization methods include radio frequency heating (Zheng et al., 2016), microwave heating (Chen et al., 2016), infrared heating (Moreirinha et al., 2016), and ohmic heating (Cho and Chung, 2016). While these methods have the advantage of rapid and uniform heating, thermal sterilization often causes damage to food products. Nonthermal sterilization inactivates microorganisms at near ambient temperatures, thereby avoiding thermal degradation of food components and maintaining the sensory and nutritional qualities of fresh foods (Olatunde and Benjakul, 2018).
Light-emitting diodes (LEDs) are cost-effective semiconductor devices (Kumar et al., 2016). The mechanism of heat generation in LEDs is the non-radiative recombination process (Luo et al., 2016). Its central structure is a semiconductor. The LED releases a certain amount of energy through the difference between the inherent conduction band and valence band energy levels of the semiconductor, thereby emitting a certain wavelength of light to produce light radiation (Chen et al., 2017). The damage caused by light irradiation depends on many variables, including exposure time, wavelength, and light intensity (Song et al., 2016).
In recent years, LEDs have attracted widespread attention because they are a nonthermal, environmentally friendly, efficient, reliable, long-lasting way to control bacterial contamination (Fan et al., 2015). Illumination with 405-nm LED has been demonstrated to have inactivation effect, such as against Lactobacillus plantarum, Staphylococcus aureus, and Vibrio parahaemolyticus in phosphate-buffered saline (PBS) solution (Kumar et al., 2016), Escherichia coli O157:H7, Listeria monocytogenes, and Salmonella spp. cells on the surfaces of mangoes (Kim et al., 2017c), and Campylobacter spp. on chicken skin (Sommers et al., 2017).
The aims of the current study were to assess the antibacterial effects of 405-nm LED treatment against S. flexneri and to examine the underlying mechanism of inhibition. Reconstituted infant formula (RIF), fresh-cut carrot slices, and stainless steel surfaces were used as media and contact surfaces to evaluate the inhibitory effects of 405-nm LED treatment. In addition, by measuring the changes in cell morphology and membrane integrity, and by investigating the site of cellular damage and intracellular protein content, possible mechanisms of action were explored.
Materials and Methods
Bacterial strain and culture conditions
Shigella flexneri strain ATCC 12022 was purchased from the American Type Culture Collection (ATCC, Manassas, VA, USA) and was stored at −80°C. Bacteria were subcultured on Luria–Bertani (LB) agar for 18 h at 37°C. Before each experiment, a loopful of S. flexneri was inoculated into LB broth and incubated for 18 h at 37°C. This overnight culture was then centrifuged at 8000 × g for 5 min at 4°C, and the cell pellet was washed twice with PBS (pH 7.4) for use in subsequent experiments.
LED illumination
The 405-nm LED used in this study was purchased from Shenzhen Boya Technology Co., Ltd. (Shenzhen, China). The LED illumination system was designed as reported by Ghate et al. (2013). Briefly, an acrylonitrile butadiene styrene housing unit was used in the LED system to block external light during illumination. To minimize heat transfer to samples and to protect the LED from its own heat, a cooling fan and a heat sink were attached to the LED. To protect the LED from excessive current, 5 Ω of resistance was linked into the circuit.
The LED is placed directly above the sample. The sample is placed in a sterile dish (90 × 15 mm), keeping a distance of 4.5 cm in each experiment to ensure that the entire Petri dish is exposed (Supplementary Fig. S2). The exposure dose was calculated using the equation (Maclean et al., 2009): E = Pt, where E is dose (energy density) in J/cm2, P is the irradiance (power density) in W/cm2, and t is the time in seconds. The power density was measured using a 405-nm radiometer (UHC405; UVATA Ltd., Hong Kong, China), with the value recorded as 61.3 ± 0.2 mW/cm2.
Determination of compensation temperature
A digital thermometer (DTM-280; Shanghai Shuangqiao Instrument Co., Shanghai, China) was used to monitor fluctuations in the temperature of the PBS, RIF, fresh-cut carrot slice, and stainless steel during LED illumination. The LED system and samples were placed in biochemical incubators (SPX-160B; Shanghai Nanrong Laboratory Equipment Co., Shanghai, China) at 4°C or 10°C. Sample temperature was recorded every 1 min for 126 min (PBS), 70 min (RIF), 70 min (fresh-cut carrot slice), or 60 min (stainless steel) to monitor temperature changes. Unilluminated samples (controls) were placed in biochemical incubators set at the corresponding measured temperatures to compensate for the temperature increase during LED illumination.
Bacterial inoculation and LED illumination in PBS
A Shigella flexneri ATCC 12022 cell pellet was resuspended in PBS and diluted to obtain the required concentration (∼6.8 log colony-forming unit [CFU]/mL). The bacterial suspension was illuminated using the 405-nm LED (61.3 ± 0.2 mW/cm2) for 360 min at 4°C. Unilluminated control samples were incubated at 11.9°C for an equivalent period. After 0, 30, 60, 90, 120, 180, 240, and 360 min, aliquots of both the illuminated and unilluminated samples were plated directly or were serially diluted 10-fold with PBS, and 100 μL of the diluents was spread plated in duplicate on LB agar plates. Five 200 μL aliquots of the undiluted samples also were spread plated onto five LB agar plates to achieve a detection limit of 1 CFU/mL. The samples were incubated for 18 h at 37°C.
The plates were incubated for 18 h at 37°C for colony enumeration. Bacterial counts were calculated in log CFU/mL, and the rate of decline was determined by plotting the change in log CFU/mL counts over time.
LED illumination of S. flexneri in RIF
A commercially available brand of infant formula was purchased from Suzhou Greenbell Trading Co. and reconstituted as per the instructions on the label. Briefly, 26.4 g of powdered formula was reconstituted in 180 mL of sterile distilled water and pasteurized at 63°C for 30 min, before being cooled to room temperature for later use. The samples were tested and proved that there was no background microflora in pasteurized RIF. A Shigella flexneri ATCC 12022 cell pellet was resuspended in PBS to obtain a concentrated cell culture. A 2 mL aliquot of the above bacterial suspension was then inoculated into 98 mL of the infant formula to obtain a final concentration of ∼6.8 log CFU/mL.
Two aliquots (25 mL) of the inoculated formula were aspirated into sterile Petri dishes (90 × 15 mm). One sample was placed directly into the LED system as previously described and was illuminated using the 405-nm LED for 360 min at 4°C. The second sample (control) was placed in a biochemical incubator at 9.5°C for an equivalent period. After 0, 60, 120, 180, 240, 300, and 360 min, aliquots of two samples were collected and diluted in sterile PBS before being plated onto LB agar plates. The plates were incubated for 18 h at 37°C for colony enumeration.
LED illumination of S. flexneri on fresh carrot slices
Fresh carrots were purchased from a local supermarket in Xianyang, Shaanxi Province, China. Carrots were washed, peeled, and then cut into small slices (1 cm × 1 cm × 0.5 cm). The carrot slices were surface-sterilized with 1% (v/v) sodium hypochlorite for 30 min, rinsed three times with sterile deionized water, and then dried on the benchtop for 5 min (Franz et al., 2006). The samples were tested and proved that there was no background microflora on fresh carrot slices anymore.
A Shigella flexneri ATCC 12022 cell pellet was resuspended in 0.2% (w/v) buffered peptone water (BPW) to obtain a concentrated cell suspension (∼8.6 log CFU/mL). Aliquots (50 μL) of this suspension were inoculated onto the carrot slices, which were then dried on the benchtop for 5 min.
Slices in the treatment group were illuminated using the 405-nm LED at 4°C for 2 h, whereas samples corresponding to the control group were incubated at 15.7°C for the same period. After 0, 20, 40, 60, 90, 120 and 150 min, carrot slices were aseptically placed into sterile homogenizer bags containing 20 mL of 0.2% (w/v) BPW. Each sample was then homogenized for 4 min in a patch-type sterilizing homogenizer (DH-11L; Lawson, Ningbo, China) at a frequency of 12 beats per minute. Samples were then serially diluted in 0.2% (w/v) BPW and plated onto LB agar plates. The plates were then incubated for 18 h at 37°C for colony enumeration.
LED treatment of S. flexneri biofilms
The effects of 405-nm LED treatment on S. flexneri biofilms formed on stainless steel surfaces were determined as described previously by Amalaradjou and Venkitanarayanan (2011), with some modifications. A Shigella flexneri ATCC 12022 cell pellet was resuspended in LB broth to obtain a concentrated suspension (∼8.6 log CFU/mL). Aliquots (30 mL) of this suspension were then added to 50-mL centrifuge tubes containing stainless steel sheets (50 mm × 20 mm) and incubated for 24 h at 25°C to allow mature biofilms to form on both sides of the stainless steel sheets.
Following incubation, the sheets were removed and washed twice with sterile water to remove planktonic bacterial cells. The stainless steel sheets were then illuminated using the 405-nm LED at 10°C for 4 h. Unilluminated stainless steel sheets (control) were incubated at 27.4°C for the same period. To make the biofilms on both sides of the stainless steel sheet be treated by LEDs, the stainless steel sheets were placed under the LED sterilization device perpendicular to the ground. After 0, 30, 60, 120, 180, and 240 min, two samples were transferred to 50-mL centrifuge tubes containing 30 mL of PBS and 3 g of glass beads (G8772, 425–600 μm; Sigma–Aldrich, St. Louis, MO, USA) and vortexed for 5 min. Aliquots of the resulting suspensions were collected, diluted in sterile PBS, and plated on LB agar plates. The plates were incubated for 18 h at 37°C for colony enumeration.
Possible mechanism of action of 405-nm LED treatment
Location of cellular damage caused by LED illumination
To investigate the cellular location of damage in S. flexneri cells caused by 405-nm LED illumination, three different antibiotics targeting different cellular locations were used as metabolic inhibitors (Kohanski et al., 2010; Ha and Kang, 2014), including ampicillin (targets the cell wall), chloramphenicol (targets the 50S ribosomal protein), and nalidixic acid (targets DNA). A Shigella flexneri ATCC 12022 cell pellet was resuspended in LB broth to obtain a concentrated suspension before being diluted to obtain a final cell concentration of 5 × 105 CFU/mL.
Aliquots (100 μL) of the diluted bacterial suspension were transferred into the wells of 96-well microtiter plates, and equivalent volumes of the antibiotic solutions were added to achieve final concentrations of 0 (control), 0.750, 0.500, 0.375, 0.250, 0.188, 0.125, 0.094 μg/mL. LB broth was used as the negative control. To assess bacterial growth, the initial absorbance of each well at 600 nm was determined before incubation, and absorbance readings were again determined following incubation at 37°C for 24 h. The maximum non-inhibitory concentration of the antibiotics was defined as the highest concentration at which bacterial growth was not impeded compared with control.
LB agar plates supplemented with the maximum non-inhibitory concentrations of each of the antibiotics were then prepared. S. flexneri suspension was illuminated using the 405-nm LED for 90 min at 4°C, whereas the unilluminated samples were incubated at 11.9°C for the same period. After 90 min, aliquots of both the illuminated and unilluminated samples were collected, diluted, and plated onto LB agar plates with and without antibiotics. The plates were incubated for 18 h at 37°C for colony enumeration. Sites of cellular damage were determined by comparing the numbers of surviving cells on LB agar plates (non-selective medium) with those on LB agar plates supplemented with various concentrations of each antibiotic (selective medium). Bacterial metabolic inhibition (%) was then calculated using the following equation:

Examination of membrane integrity following LED illumination
A Shigella flexneri ATCC 12022 cell pellet was resuspended in PBS and diluted to 8.4 log CFU/mL. Samples in the treatment group were subjected to LED illumination at 4°C for 3 h, whereas samples in the control group were cultured in the dark at 11.9°C for the same period. After 90 and 180 min, aliquots were collected from each sample, centrifuged at 10,000 × g for 2 min at 4°C, and the resulting cell pellets resuspended in 0.85% (w/v) NaCl. Two fluorophores, SYTO 9 and propidium iodide, were mixed and then added to the bacterial suspensions. The samples were incubated at room temperature in the dark for 5 min (Stiefel et al., 2015). Droplets of the stained bacteria were then placed on glass slides, covered with coverslips, and observed using a confocal laser scanning microscope (A1; Nikon, Tokyo, Japan).
Morphological examination of S. flexneri cells following LED illumination
Scanning electron microscopy was performed to observe morphological changes in the S. flexneri cells following illumination as described by Zheng et al. (2020), with some modifications. A Shigella flexneri ATCC 12022 cell pellet was resuspended in PBS to the required concentration (∼6 × 107 CFU/mL). S. flexneri in the treatment groups were then illuminated using the 405-nm LED at 4°C for 6 h, whereas the unilluminated control samples were incubated at 11.9°C for the same time period. After 240 and 360 min, aliquots of two samples were harvested via centrifugation at 5000 × g for 5 min at 4°C and then rinsed twice with PBS.
The washed cells were then resuspended in a fixative solution (2.5% [v/v] glutaraldehyde) for 12 h at 4°C before being washed with PBS. The cells were resuspended in 2.5% (v/v) glutaraldehyde, incubated for 6 h at 4°C, and then washed with PBS followed by sterile water. The samples were dehydrated using an ethanol gradient (30% [v/v], 50%, 70%, 80%, 90%, and 100%) and then fixed on microscope support, sputter-coated with gold under a vacuum, and examined using a field emission scanning electron microscope (FESEM, S-4800; Hitachi, Tokyo, Japan) under 400 × magnification.
Determination of total cellular protein concentration
A Shigella flexneri ATCC 12022 cell pellet was resuspended in PBS to obtain a concentrated cell suspension (∼8.4 log CFU/mL). Samples in the treatment group were then illuminated using the 405-nm LED at 4°C for 0, 2, 4, and 6 h, whereas the unilluminated control samples were incubated at 11.9°C for the same period. After 0, 120, 240, and 360 min, samples were centrifuged at 10,000 × g for 5 min at 4°C and then washed with PBS. Total cellular protein was extracted from planktonic cells using a Bacterial Total Protein Extraction Kit (BB-3130; BestBio, Shanghai, China) as per the manufacturer's instructions.
Protein concentrations were determined using a Bradford Protein Assay Reagent Kit (BestBio) as per the manufacturer's instructions using bovine serum albumin as the standard (Zhou et al., 2016). Briefly, 12.5 μL of different concentrations of the protein standard solution and each of the samples were added to the wells of 96-well microtiter plates. A 100 μL aliquot of working solution was added to each well, and plates were incubated for 30 min at 37°C. Following incubation, the absorbance of each well at 570 nm was determined, a standard curve was constructed, and the concentration of protein in each sample was calculated.
Statistical analyses
The mean values were calculated from independent triplicates, with duplicate sampling (n = 6). Statistical analyses were carried out using SPSS software (version 20.0; IBM Corp, Armonk, NY, USA). Data are presented as the mean ± standard deviation (n = 3). Differences between means were tested using a Student's t-test. Differences were defined as significant at p < 0.05 and extremely significant at p < 0.01.
Results
Temperatures of PBS, RIF, fresh-cut carrot slices, and stainless steel sheets during LED illumination
When PBS, RIF, and fresh-cut carrot slices were illuminated using the 405-nm LED (61.3 ± 0.2 mW/cm2) at the set temperature of 4°C for 150, 100, and 100 min, respectively, the average temperatures were increased to 11.9, 9.5, and 15.7°C, respectively (Supplementary Fig. S1A–C). The average temperature of the stainless steel sheets increased to 27.4°C when illuminated at 10°C for 80 min (Supplementary Fig. S1D). Thus, the unilluminated samples in PBS or RIF or on fresh-cut carrot slices or stainless steel sheets were incubated at 11.9, 9.5, 15.7, and 27.4°C, respectively, to eliminate the influence of the temperature rise of the sample caused by the LED irradiation.
Survival of S. flexneri in PBS following 405-nm LED treatment
The initial concentration of viable S. flexneri cells in PBS was ∼6.81 log CFU/mL. In the unilluminated samples, viable cell counts remained relatively stable over the 360 min experimental period. In contrast, while viable cell counts remained steady for the first 60 min in the 405-nm LED-treated samples, viable cell counts dramatically decreased between 60 and 360 min. At the 90- and 180-min time points, S. flexneri concentrations were 6.00 ± 0.09 and 5.10 ± 0.28 log CFU/mL, respectively, and decreased by 3.29 log CFU/mL at 360 min (Fig. 1A).
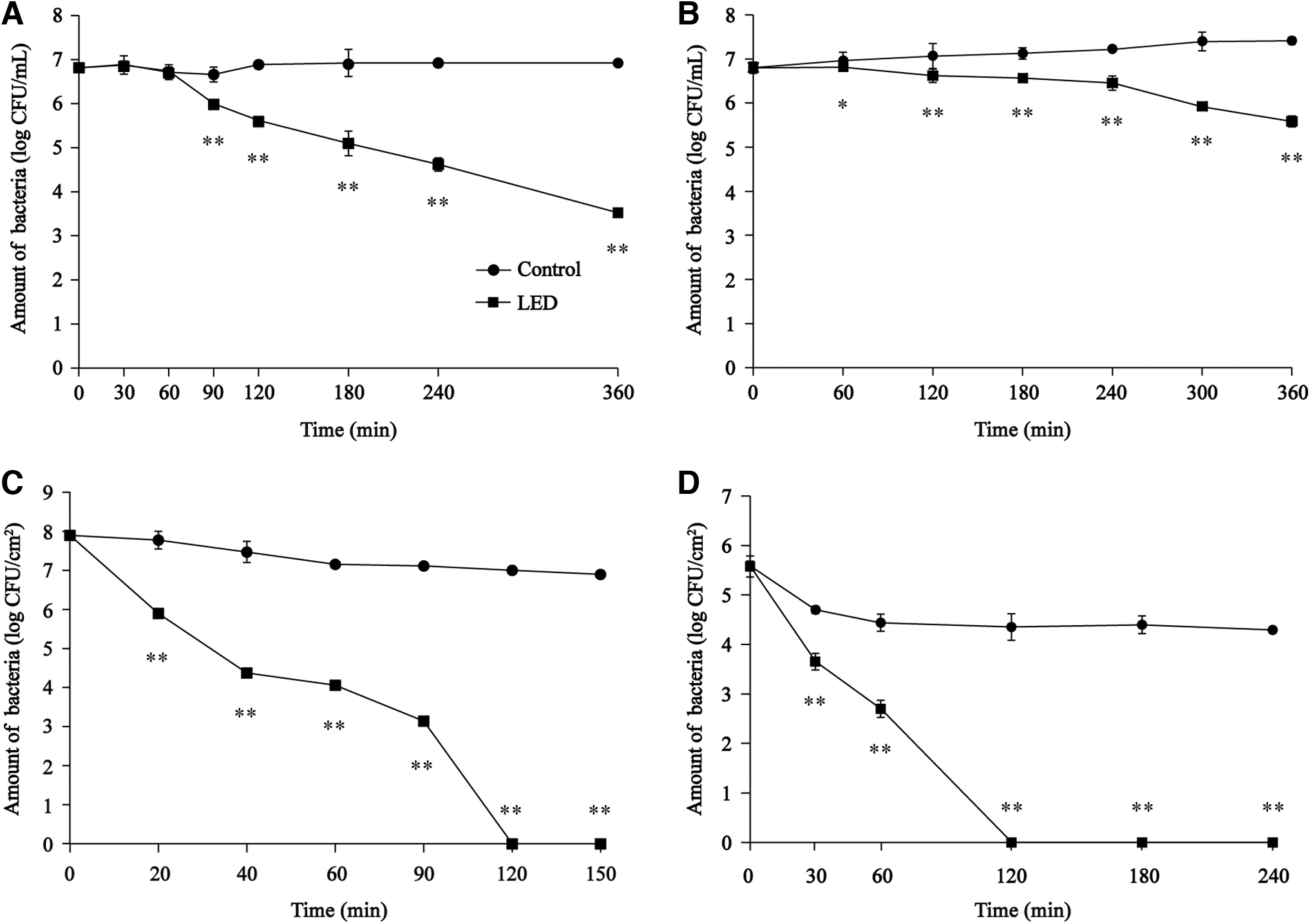
Effects of 405-nm LED illumination on the survival of Shigella flexneri in
Survival of S. flexneri in RIF following 405-nm LED treatment
The initial bacterial concentration in all RIF samples was ∼6.80 log CFU/mL. Over the course of the experiment, the number of viable S. flexneri cells in the unilluminated samples incubated at 4°C slowly increased to a maximum of 7.41 ± 0.09 log CFU/mL. When illuminated with the LED, the number of viable cells decreased by 0.35 log CFU/mL over the first 4 h, with a viable cell count after 360 min of illumination of 5.58 ± 0.12 log CFU/mL (Fig. 1B).
Survival of S. flexneri on fresh-cut carrot slices following 405-nm LED treatment
The initial concentration of S. flexneri on fresh-cut carrot slices was ∼7.90 log CFU/cm2. The viable cell count in the unilluminated samples decreased slowly over the course of the experiment, reaching 6.90 ± 0.09 log CFU/cm2 after 150 min. However, a dramatic decrease in the survival of illuminated S. flexneri was observed, with an ∼55% decrease (4.38 log CFU/cm2) in viable cell numbers after 40 min. After 120 min, viable cell counts in the illuminated samples had dropped below the limit of detection (20 CFU/cm2) (Fig. 1C).
Effects of 405-nm LED treatment on the survival of biofilm-associated S. flexneri cells
As shown in Figure 1D, a decrease in the number of viable S. flexneri cells in biofilms was observed following treatment with the 405-nm LED. The initial bacterial concentration in all samples was ∼5.58 log CFU/cm2. In the unilluminated samples, the number of viable cells decreased by about 1.15 log CFU/cm2 after 60 min but remained stable thereafter, with a final viable cell count of 4.29 ± 0.05 log CFU/cm2 recorded at the end of the experimental period (240 min). In comparison, a significant decrease in the number of viable biofilm-associated S. flexneri cells was observed following LED illumination. In this group, a decrease of ∼1.05 log CFU/cm2 was observed after 30 min, with viable cell counts then continuing to decrease before finally dropping below the limit of detection (1.5 CFU/cm2) after 120 min.
Mechanism of inhibitory action of LED illumination against S. flexneri
Sites of cellular damage in S. flexneri caused by LED illumination
Preliminary experiments showed that the growth of S. flexneri was not inhibited by chloramphenicol at concentrations ≤8 μg/mL, indicating non-sensitivity. Therefore, chloramphenicol was not used in subsequent assays to determine the sites of cellular damage in S. flexneri resulting from LED illumination. The maximum non-inhibitory concentrations of ampicillin and nalidixic acid against S. flexneri were 0.125 and 0.5 μg/mL, respectively.
Metabolic inhibition caused by ampicillin and nalidixic acid in S. flexneri cultures in PBS at 4°C was determined as a percentage, and results are shown in Figure 2. The metabolic inhibition caused by the two antibiotics in non-illuminated cells was <0.6% in both cases. In contrast, LED illumination resulted in a significant (p < 0.01) increase in metabolic inhibition caused by ampicillin (2.2%) and nalidixic acid (7.9%).

Changes in metabolic inhibition of Shigella flexneri by Amp and Nal following LED illumination in phosphate-buffered saline at 4°C. **p < 0.01 versus the control. Amp, ampicillin; LED, light-emitting diode; Nal, nalidixic acid.
Changes in the membrane integrity of S. flexneri cells as a result of LED illumination
The effects of 405-nm LED treatment on the cell membrane integrity of S. flexneri were examined using fluorescence staining (Fig. 3). The majority of unilluminated S. flexneri cells showed green fluorescence at both time points (Fig. 3A, B), indicating intact cellular membranes. In comparison, some yellow and red fluorescence was observed in cultures subjected to LED illumination for 90 min (Fig. 3C), indicative of cell membrane damage, bacterial cells cannot survive. Following exposure to LED illumination for 180 min (Fig. 3D), a marked decrease in green fluorescence was observed, with an abundance of red fluorescence in the field of view. These results indicated that 405-nm LED treatment impacts membrane integrity in S. flexneri cells.

Confocal laser scanning microscope images of Shigella flexneri. Untreated S. flexneri incubated for 90 min
Morphology of S. flexneri cells following LED illumination
Changes in cell morphology were observed using FESEM. As shown in Figure 4, untreated cells appeared smooth (Fig. 4A, B), whereas cells illuminated for 4 h (Fig. 4C) or 6 h (Fig. 4D) were wrinkled and had a rough surface. With increases in incubation time, the degree of cell shrinkage increased. No cell rupture was observed at any of the time points.

Field emission scanning electron micrographs of untreated Shigella flexneri cells at 4 h
Effect of 405-nm LED treatment on total protein concentration
Figure 5 shows the effect of 405-nm LED on the total protein concentration of S. flexneri in PBS at 4°C. Over the course of the experiment, compared with the unilluminated samples, the protein concentration of the LED illuminated samples decreased by 0.14, 0.53, and 0.72 mg/mL, respectively, at 2, 4, and 6 h. The protein concentration of bacteria sample in two samples differed significantly (p < 0.05) at the 2, 4, and 6 h time points. The results showed that the total concentration of S. flexneri could be decreased under 405-nm LED illumination.
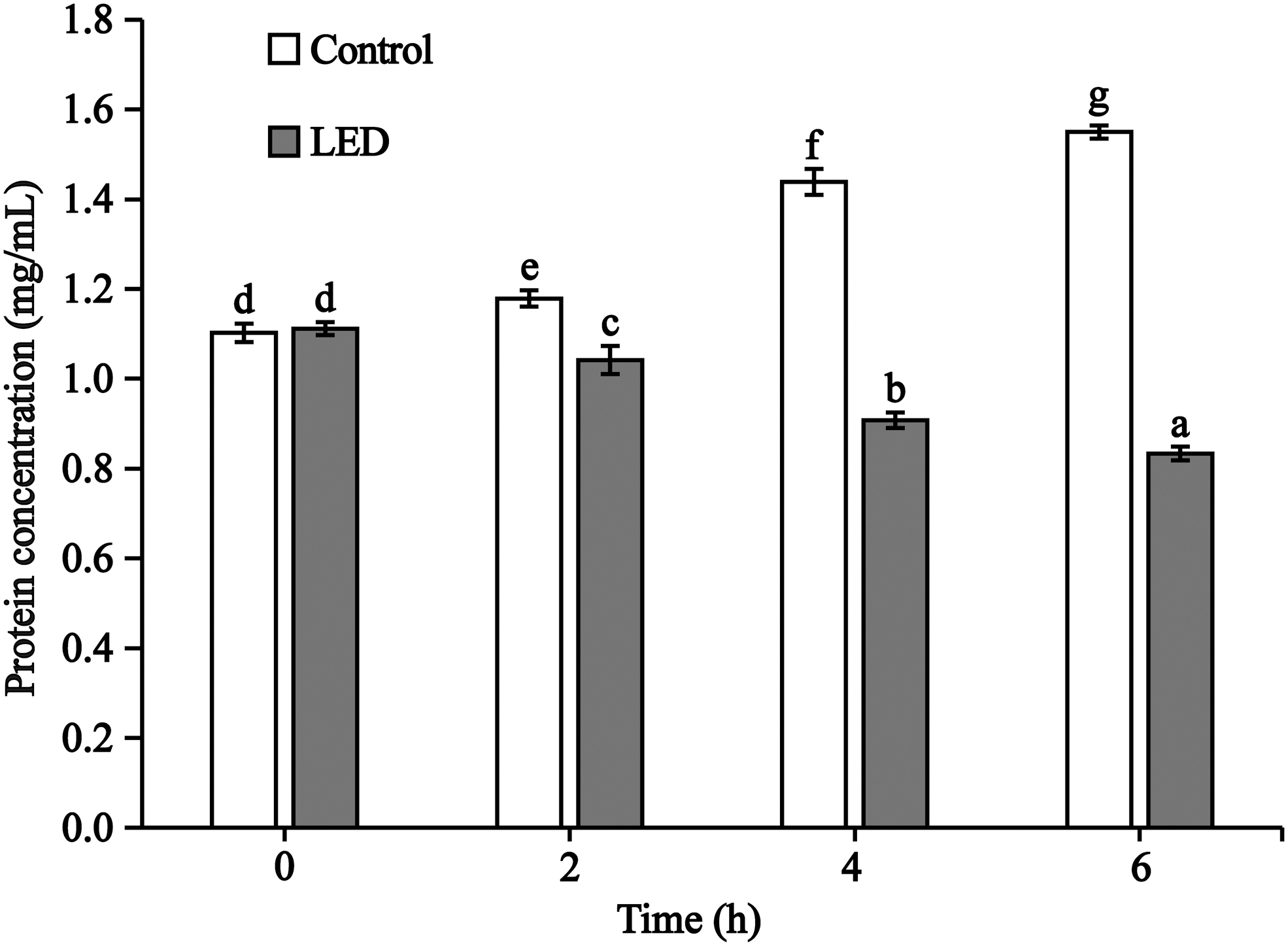
Changes of total protein content treated with LED illumination in phosphate-buffered saline at 4°C. Values having different letters (on the top of the columns) are significantly different from each other (p < 0.01). LED, light-emitting diode.
Discussion
Light inactivation is a promising method for controlling microbial contamination in medical environments (Kumar et al., 2016), leading us to explore the potential of this new technology in the field of food safety. Previous studies have demonstrated the antibacterial effects of 405-nm LED treatment on a variety of foodborne pathogens (Kim and Yuk, 2017; Hyun and Lee, 2020), yet nothing is known about the antibacterial effects, or the underlying mechanism, of LED illumination on S. flexneri. Here, we explored the antibacterial effects of 405-nm LED treatment on planktonic S. flexneri in laboratory media (PBS), RIF, and carrot slices and on biofilm-associated S. flexneri on stainless steel sheets in an attempt to clarify the possible mechanism of inhibition.
Our results showed that 405-nm LED (61.3 ± 0.2 mW/cm2) treatment had a good inhibitory effect on S. flexneri in PBS at 4°C, with the viable cell count reduced by 3.3 log after 360 min (Fig. 1A). This result agrees with the findings of a previous study showing that L. monocytogenes viable cell counts in PBS were reduced by 3.6 log following 405-nm LED illumination (18.9 ± 1.4 mW/cm2) for 180 min at 4°C (Kang et al., 2019). Similarly, Maclean et al. (2009) reported that the concentration of E. coli in PBS was reduced by 3.1 log after 300 min of exposure to a 405-nm LED (10 mW/cm2) at 27°C. While these findings suggest that LED treatment negatively impacts the survival of a variety of foodborne pathogens, including S. flexneri, the sterilization efficiency of LED treatment may be variable.
Treatment efficacy may be related to the bacterial species, with differences in sterilization device, radiation dose, reaction temperature, illumination time, and other factors also likely to play a role. In addition, we found that there was no obvious change in the viability of S. flexneri over the first 60 min of treatment (Fig. 1A). A similar study showed that the number of viable Salmonella enterica serovar Enteritidis and Mycobacterium tuberculosis cells was not significantly altered compared with the control following 405-nm LED illumination at a dose of 50 J/cm2 (Murdoch et al., 2012). However, the number of viable Salmonella Enteritidis cells was reduced by 3.5 log at a dose of 288 J/cm2, and viable M. tuberculosis cells were reduced by 4–5 log at an illumination dose of 144–288 J/cm2 (Murdoch et al., 2012).
We speculate that at lower irradiation doses, cell damage must be accumulated to a certain level before any death of S. flexneri cells is observed, after which the numbers of viable bacteria will continue to decline.
S. flexneri can be detected in RIF and has been implicated in a variety of illnesses and even deaths associated with ingestion of contaminated products (Wu et al., 2002). It is also commonly isolated from fresh-cut fruits and vegetables (Kang et al., 2018). Therefore, RIF and fresh carrot were used in this study to investigate the antibacterial effects of 405-nm LED treatment on S. flexneri. The results showed that illuminated samples had significantly decreased viable cell counts compared with unilluminated samples in RIF (Fig. 1B) and on fresh carrot slices (Fig. 1C). In addition, we found that bacterial cell counts on carrot slices decreased faster (Fig. 1C) than those in RIF and PBS (Fig. 1A).
This is possibly because the penetration capacity of 405-nm LED illumination limits the sterilization effects. In addition, RIF can provide bacteria with some protection against damage caused by reactive oxygen species (ROS). In a similar study, Ghate et al. (2019) used a 410-nm LED device to illuminate L. monocytogenes in LB broth and on paperboard coupons and found that bacterial cell counts on paperboard coupons decreased faster than those in LB broth.
S. flexneri adheres to and forms biofilms on solid surfaces throughout the food production chain, causing cleaning difficulties and increasing the risk of cross-contamination (Kang et al., 2018). To investigate the effects of 405-nm LED treatment on the survival of S. flexneri in mature biofilms, we used stainless steel sheets to simulate solid surfaces in food processing environments. Our results revealed a significant decrease in viable cell numbers in mature biofilms following LED treatment, with S. flexneri concentrations dropping below the limit of detection limit (1.5 CFU/cm2) after 120 min.
Similarly, Yang et al. (2020) showed that 405-nm LED treatment significantly reduced the number of viable Pseudomonas aeruginosa cells in biofilms on stainless steel coupons by about 4.41 log CFU/cm2 within a 4-h period, whereas McKenzie et al. (2013) found that a mixed biofilm of E. coli and S. aureus was successfully inactivated by 405-nm LED treatment, reducing the total viable cell population by ∼2.2 log CFU/mL within 30 min. Recently, photosensitizers have been shown to enhance the sterilization effects of LED-based illumination on planktonic bacteria (Ke et al., 2012). Thus, we plan on investigating the effects of LED illumination combined with photosensitizers or other ways on the viability of biofilm-associated S. flexneri and other foodborne pathogens.
In this study, ampicillin and nalidixic acid were used to study the mechanism of metabolic inhibition of S. flexneri in PBS following 405-nm LED illumination at 4°C. Nalidixic acid targets bacterial DNA replication (Kim et al., 2017b), whereas ampicillin inhibits the synthesis of peptidoglycan, resulting in lysis and cell death (Raynor, 1997). If LED illumination affects DNA, RNA, protein, or cell wall synthesis, bacterial cells are likely to be incapable of multiplying on the selective medium containing the tested antibiotics (Kim et al., 2017b). Our results showed that in the presence of ampicillin or nalidixic acid, metabolic inhibition (%) was significantly increased in LED-treated samples compared with unilluminated samples, suggesting that DNA and the cell wall are damaged by LED illumination of S. flexneri.
Similarly, Kim et al. (2017b) found that LED illumination increased the metabolic inhibition of Salmonella Enteritidis by nalidixic acid, rifampicin, chloramphenicol, and ampicillin by 30.1%, 44.1%, 24.2%, and 32.5%, respectively. The authors therefore speculated that the sites of cellular damage might include DNA, RNA, proteins, and the cell wall. Another study found that the levels of 8-hydroxyde-oxyguanosine (8-OHdG), an oxidized derivative of guanine residues, in genomic DNA of illuminated Salmonella enterica serovar Agona and Salmonella enterica serovar Typhimurium cells were 1.7–1.8 times higher than those of non-illuminated cells. Therefore, the authors speculated that ROS produced by LED illumination might oxidize DNA (Kim et al., 2017a).
In the current study, morphological changes in S. flexneri cells caused by LED illumination were observed by FESEM (Fig. 4), whereas cell membrane damage was examined using confocal laser scanning microscope (Fig. 3). Kim and Yuk (2017) used flow cytometry to show that the permeability of Salmonella Enteritidis cells was increased following treatment with LED illumination. They speculated that internal ROS produced by LED illumination can attack proteins related to the function of efflux pumps, thereby destroying cell membrane function. Hyun and Lee (2020) treated L. monocytogenes with LED illumination for 4 d and found that intracellular components were released and some cells showed a fuzzy cell membrane.
Combined with propidium iodide absorption measurements, the authors confirmed that the destruction of the cell membrane was the main factor causing bacterial inactivation under 460- and 470-nm LED illumination. We hypothesize that ROS stimulated by LED treatment in S. flexneri destroy the integrity and function of the cell membrane, disrupting the flow of molecules into and out of the cell and causing changes in cell morphology.
Proteins account for more than 75% of the bacterial dry cell weight and are therefore an important part of the bacterial cell structure. Our study showed that the total protein content of S. flexneri cells was significantly reduced by 405-nm LED illumination. The generation of ROS plays an important role in causing cell damage by oxidizing intracellular biomolecules such as nucleic acids and proteins (Ahn et al., 2012). We hypothesize that the decrease in S. flexneri total protein content following LED illumination is the result of ROS production, destroying protein structure and/or affecting protein synthesis. LED illumination may also increase the permeability of cells, resulting in protein leakage.
Conclusions
This is the first study to investigate the antibacterial effects and possible mechanism of 405-nm LED illumination on S. flexneri. Our results demonstrated that 405-nm LED illumination effectively reduces S. flexneri viable cell counts in PBS, RIF, and fresh-cut carrot slices. Similarly, LED illumination had good bactericidal activity against biofilm-associated S. flexneri on stainless steel surfaces. Our results also confirmed that cellular damage caused by LED illumination may involve the bacterial cell wall and DNA, with LED treatment significantly increasing the permeability of the bacterial cell membrane, inducing morphological changes, and reducing the total intracellular protein concentration (Supplementary Fig. S3).
In light of these observed antibacterial properties, together with its superiorities for being safe, green, efficient, and low-cost, 405-nm LED illumination has significant potential for use in controlling S. flexneri in food processing, storage, and preparation situations.
Footnotes
Disclosure Statement
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this article.
Funding Information
This work was supported by the National Natural Science Foundation of China (31801659), the class General Financial Grant from the Shaanxi Postdoctoral Science Foundation (2018BSHEDZZ150), and the Key Research and Development Program of Shaanxi Province, China (2021NY-125).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.