
Abstract
The objective of this study was to clarify the antibacterial activity and mechanism of Chrysanthemum buds' crude extract (CBCE) against Salmonella Typhimurium, and explore the potential application in cooked chicken. The zone of inhibition (ZI), minimum inhibitory concentration (MIC), and minimum bactericidal concentration (MBC) were used to assess the in vitro antibacterial activity of CBCE against Salmonella Typhimurium. The antibacterial mechanism was elucidated by revealing the changes in intracellular adenosine 5′-triphosphate (ATP) concentration, membrane potential, content of biomacromolecule, and cell morphology. Furthermore, the effect of CBCE on the counts of Salmonella Typhimurium and color of cooked chicken during storage was studied. The results showed that the ZI, MIC, and MBC of CBCE against Salmonella Typhimurium were 12.9 ± 0.53–13.6 ± 0.14 mm, 40, and 80 mg/mL, respectively. In the process of inhibiting Salmonella Typhimurium by CBCE, the reduction of intracellular ATP concentration, cell membrane depolarization, leakage of protein and nucleic acid, and destruction of cell morphology were observed. Moreover, after treatments with CBCE, the growth of Salmonella Typhimurium in cooked chicken was significantly inhibited (p < 0.05) compared with the control group. No significant differences (p > 0.05) in lightness (L*), redness (a*), and yellowness (b*) values of cooked chicken were found between untreated and treated samples, as well as the color of cooked chicken treated with CBCE did not change significantly (p > 0.05) during the six days of storage. Overall, our findings suggested that CBCE exhibited the antibacterial effect against Salmonella Typhimurium, and had the potential to be used as a natural food preservative for the control of Salmonella Typhimurium in chicken products.
Introduction
Salmonella Typhimurium is a rod-shaped, nonspore-producing, Gram-negative food-borne pathogen, which can contaminate a wide variety of foods, especially chicken products (Borch and Arinder, 2002). Importantly, Salmonella Typhimurium was considered to be one of the leading causes of bacterial food-borne diseases worldwide, and can cause intestinal fever, sepsis, and gastrointestinal diseases, seriously threatening the people's health (Wang et al., 2021). Although various efforts have been used to control the Salmonella Typhimurium contamination in food, food safety problems caused by this pathogen continue to occur (Mónica et al., 2003; Park et al., 2018).
Salmonella Typhimurium contamination in chicken products can lead to the shortened shelf life and epidemic outbreaks of food-borne diseases (Fearnley et al., 2011). To solve this problem, synthetic preservatives are often used to inhibit the colonization and growth of harmful microorganisms to extend the shelf life of food (Mishra et al., 2011). However, the potential toxicity of synthetic preservatives and adverse influence on the sensory quality and natural flavor of food have made consumers increasingly reluctant to accept them (Zhang et al., 2016). Therefore, the development of natural preservatives has attracted much attention due to their safety and good antibacterial properties.
Chrysanthemum morifolium, a well-known medicinal and edible homologous plant in China, has been reported to contain a variety of biologically active constituents, including polyphenols, essential oils, polysaccharides, which occupy an important position in traditional folk medicine because of its antioxidant, anticarcinogenic, and antibacterial activities (Zhang et al., 2008; Youssef et al., 2020). Chrysanthemum buds are made by drying the buds of C. morifolium, and are considered to be one of the highest grades of C. morifolium (Xing and Li, 2015). Modern pharmacological studies indicate that Chrysanthemum buds have the antibacterial, anti-inflammatory, and antioxidant effects (Lin and Harnly, 2010). However, the antibacterial effect of Chrysanthemum buds' crude extract (CBCE) against food-borne pathogens has rarely been reported.
Therefore, this study was conducted to evaluate the antibacterial effect of CBCE against Salmonella Typhimurium by measuring the zone of inhibition (ZI), minimum inhibitory concentration (MIC), and minimum bactericide concentration (MBC), revealing its antibacterial mechanisms of action in terms of the changes in the intracellular adenosine 5′-triphosphate (ATP) concentration, cell membrane potential, biological macromolecule concentration, and cell morphology. In addition, the antibacterial effect of CBCE was applied in cooked chicken against Salmonella Typhimurium.
Materials and Methods
CBCE material
The CBCE used in this study was in powder form, and was obtained from our team's previous report (Chang et al., 2021). The main chemical components of CBCE are shown in Supplementary Table S1. The images of Chrysanthemum buds and CBCE powders are displayed in Supplementary Figure S1.
Strains and culture conditions
The following bacterial strains were used in this study: Salmonella Typhimurium ATCC 14028 obtained from the American Type Culture Collection (ATCC, Manassas, VA) and six Salmonella Typhimurium isolates from commercial cooked chicken. All strains were used for the antibacterial activity test of CBCE against Salmonella Typhimurium, and Salmonella Typhimurium ATCC 14028 was used for the mechanism test. Before the experiments, 100 μL stock culture frozen at −80°C was inoculated into 10 mL Luria-Bertani (LB) broth and grown at 37°C for 20 h. Then, the culture was streaked on tryptone soya agar (TSA) and incubated at 37°C for 24 h to obtain isolated colonies. A single typical colony was transferred into LB broth and incubated at 37°C for 20 h for later experiments.
Determination of ZI
The ZIs of CBCE for six Salmonella Typhimurium isolates were determined using the Oxford cup method reported by Song et al. (2020), and Salmonella Typhimurium ATCC 14028 was used as the quality control strain. Briefly, 20 mL of TSA medium was poured onto the sterile Petri dish and allowed to solidify. Then, 100 μL of suspensions containing 107 colony-forming unit (CFU)/mL of Salmonella Typhimurium was uniformly distributed on TSA plates with a glass triangle coated rod. Three sterile oxford cups with 8 mm diameter were put on the surface of the above plates, and 200 μL of CBCE (100 mg/mL) was added in the cups. An equal amount of the sterilized normal saline (NS) was used as a negative control. After incubation at 37°C for 24 h, the ZIs were measured, and the mean of readings was calculated.
Determination of MIC and MBC
The MICs and MBCs of CBCE against Salmonella Typhimurium strains were determined according to the previous report (Fei et al., 2020). Briefly, the sterilized TSA medium was added into the 24-well plate, followed by the addition of CBCE at final concentrations of 0, 5, 10, 20, 40, 80, and 160 mg/mL, respectively, and 0.1 mg/mL ampicillin was used as the positive control. After solidifying, 2 μL of bacterial suspension (106 CFU/mL) was inoculated in the center of each well, and cultured at 37°C for 24 h. The MIC was interpreted as the lowest concentrations without any growth of bacteria. One hundred microliters of (106 CFU/mL) bacterial suspension was treated with 40, 80, and 160 mg/mL of CBCE for 30 min, then spread on the TSA plate followed by incubation at 37°C for 24 h. The MBC was determined as the lowest concentration at which no colonies can grow.
Measurement of intracellular ATP concentration
The intracellular ATP concentrations of Salmonella Typhimurium ATCC 14028 treated with different concentrations of CBCE were measured as described by Li et al. (2014). In brief, after treatments with 0 (control), 1 × MIC and 2 × MIC of CBCE for 30 min, 107 CFU/mL of Salmonella Typhimurium cells were centrifuged at 8000 × g for 5 min at 4°C to get the supernatants. Finally, the supernatants were used to measure the intracellular ATP concentration by an ATP Assay Kit (Beyotime Bioengineering Institute, Shanghai, China) under the guidance of the product instructions.
Membrane potential analysis
The effect of CBCE on the membrane potential of Salmonella Typhimurium ATCC 14028 was analyzed based on a previous report (Guo et al., 2020). First, 125 μL of 107 CFU/mL bacterial suspension was mixed with 0.5 μL of bis-(1,3-dibutylbarbituriis-(1,3-dibutylbarbituric acid) trimethine oxonol) (Beijing Solarbio Science and Technology Co. Ltd., Beijing, China) in the 96-well plate, followed by incubation at 37°C for 30 min. Then, CBCE was added to the mixture at final concentrations of 0 (control), 1 × MIC, and 2 × MIC. The fluorescence intensity in the cells was measured by a multifunctional microplate reader (Jinan Qiansi Biotechnology Co. Ltd., Shandong, China) according to following parameters: excitation wavelengths of 492 nm and emission wavelengths of 515 nm.
Measurement of biological macromolecule leakage
According to the report by Chang et al. (2021), 107 CFU/mL of Salmonella Typhimurium ATCC 14028 was treated by 0 (control), 1 × MIC, and 2 × MIC of CBCE at 37°C for 12 h. Every 2 h, the suspensions of the tested bacteria were prepared and the optical density (OD)260 values of suspensions were measured to the level of nucleic acid leakage. Meanwhile, every three hours, the protein content in supernatants was determined according to the Coomassie Bright Blue method.
Transmission electron microscopy
According to the report by Fei et al. (2019), the cell morphology of different treated Salmonella Typhimurium ATCC 14028 was observed using transmission electron microscopy (TEM).
Briefly, after treatments with 0 (control), 1 × MIC, and 2 × MIC of CBCE for 4 h, the cell pellets of Salmonella Typhimurium (107 CFU/mL) were collected after centrifugation at 3000 × g for 5 min, then fixed with 0.1 M sodium phosphate buffer containing 2.5% glutaraldehyde for 12 h, followed by washing three times with sterile NS. The cells were postfixed with 1% osmium tetroxide for 6 h at 25°C and washed three times with the same NS. Then, the cells were further dehydrated through various concentrations of alcohol (50%, 70%, 90%, 100%) for 10 min, embedded in Epon Lx-112 (Ladd Research, Williston, VT), and double stained with uranyl acetate and lead citrate. Eventually, cell morphology of all samples was observed using a TEM (Hitachi, Tokyo, Japan).
Application in cooked chicken samples
According to a previous report (Gong et al., 2021), the fresh chicken was cut into small pieces of about 1.5 g, then was treated at 121°C for 15 min to get the sterile cooked chicken. These small pieces were dipped into the CBCE solution with the concentrations of 1 × MIC and 2 × MIC for 10 s, then placed in an empty sterile culture dish for 30 min. The sample treated with sterile NS for 10 s was used as a control group. All samples were inoculated with Salmonella Typhimurium ATCC 14028 to a concentration of 103 CFU/g, then packed in sterile polythene bags and stored at 4°C. The viable counts of Salmonella Typhimurium strains in cooked chicken were counted on days 0, 3, and 6 by performing plate counts (Sun et al., 2018).
Color measurement
According to the previous studies (Ahn et al., 2007; Ozaki et al., 2020), the color of cooked chicken samples on days 0, 3, and 6 was measured using a CR-300 Chroma Meter (Minolta Co., Osaka, Japan), and reported with lightness (L*), redness (a*), yellowness (b*), and ΔE*, respectively. Moreover, ΔE* indicated the total color difference, and were calculated according to the formula as follows:
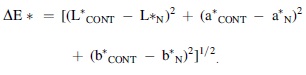
Statistical analysis
Each experiment was performed in triplicate. Data were reported as mean ± standard deviation and analyzed using the analysis of variance in the SPSS 20.0 software (SPSS, Inc., Chicago, IL). Values of p < 0.05 were considered to be statistically different.
Results
ZI, MIC, and MBC of CBCE against Salmonella Typhimurium strains
The ZIs of CBCE against all Salmonella Typhimurium strains were in the range of 12.9 ± 0.53–13.6 ± 0.14 mm and there were no significant differences (p > 0.05) among the different strains. Meanwhile, the MICs and MBCs of CBCE against all Salmonella Typhimurium strains were 40 and 80 mg/mL, respectively (Supplementary Table S2).
Changes in intracellular ATP
As shown in Figure 1, compared with untreated cells, the intracellular ATP concentrations of Salmonella Typhimurium cells treated with CBCE decreased significantly (p < 0.05), and the decrease became more significant with the increase of CBCE concentration (p < 0.05).
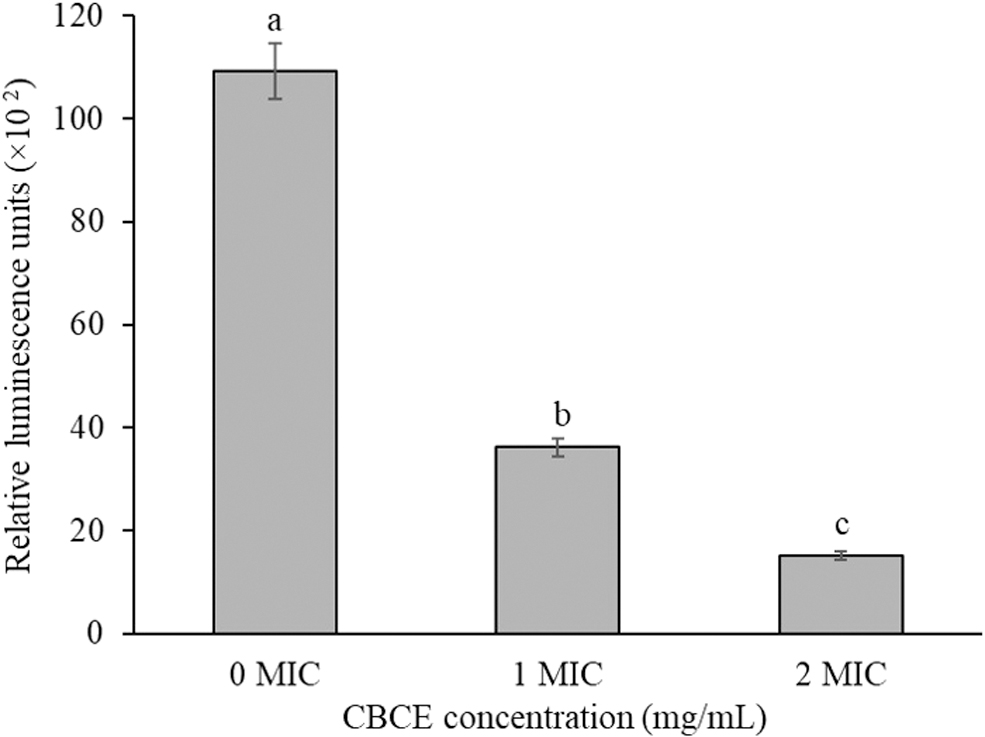
Influence of CBCE treatments at different concentrations on the intracellular ATP concentration of Salmonella Typhimurium ATCC 14028. Bars represent the standard deviation from triplicate experiments. Values with different letters are significantly different at p < 0.05. ATP, adenosine 5′-triphosphate; CBCE, Chrysanthemum buds' crude extract.
Changes in membrane potential
Figure 2 showed that after treatments with 1 × and 2 × MIC of CBCE, the fluorescence intensity of Salmonella Typhimurium ATCC 14028 was significantly increased compared with the control group (p < 0.05), which indicated that the membrane potential depolarization of Salmonella Typhimurium cell was presented under the action of CBCE.
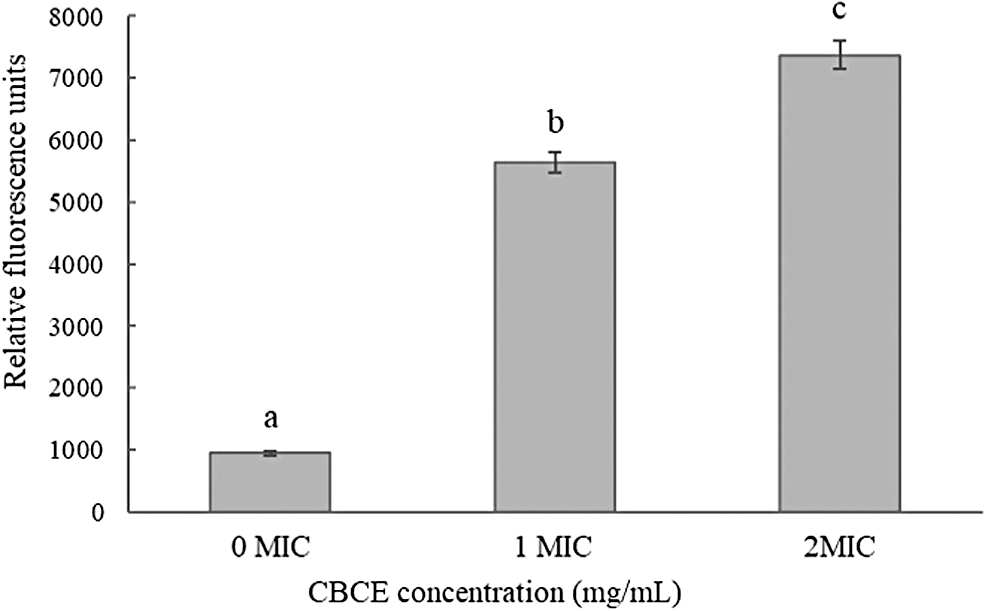
Influence of CBCE treatments at different concentrations on the membrane potential of Salmonella Typhimurium ATCC 14028. Bars represent the standard deviation from triplicate experiments. Values with different letters are significantly different at p < 0.05. CBCE, Chrysanthemum buds' crude extract.
Leakage of biological macromolecule
Figure 3A showed that nucleic acid substances in cells were obviously leaked after treatment with CBCE at different concentrations, and the leakage level increased with the increase of treatment time and concentration. Figure 3B showed that compared with the control group, the protein leakages from cells treated with CBCE for 3, 6, 9, and 12 h were significantly increased, and increased significantly with the increase of CBCE concentration (p < 0.05).
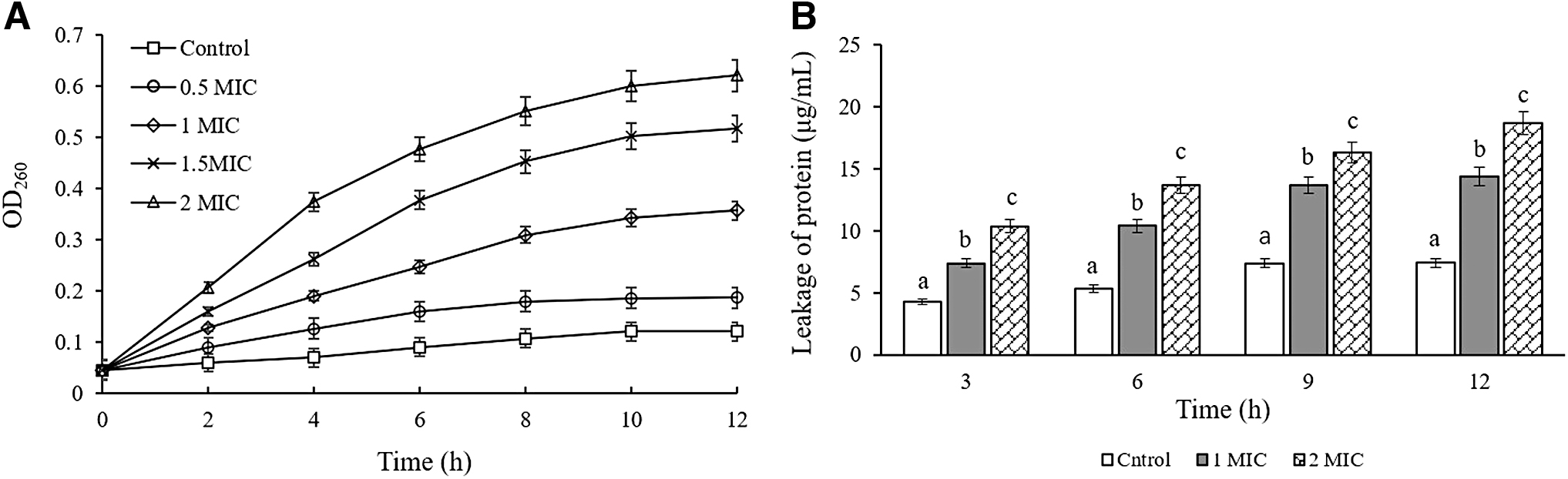
The leakage of nucleic acid
TEM observations
Figure 4A showed that the untreated cells exhibited typical rod-shaped morphology, smooth and intact surface, and full cytoplasm. After exposure to CBCE at 1 × MIC for 4 h, the cells underwent obvious morphological damage, which were characterized by sunken surface, severe deformations, separation of cell walls and membranes, and cytoplasmic leakage (Fig. 4B). Moreover, the cell treated by 2 × MIC of CBCE was more severely damaged (Fig. 4C).

Transmission electron microscopy images of Salmonella Typhimurium ATCC 14028 (40,000 × )
Application in cooked chicken
As shown in Figure 5, compared with the control group, the viable counts of Salmonella Typhimurium ATCC 14028 strains treated with CBCE in cooked chicken samples decreased significantly on the third and sixth days (p < 0.05), and the inhibitory effect was also significantly increased with the increase of CBCE concentration (p < 0.05).
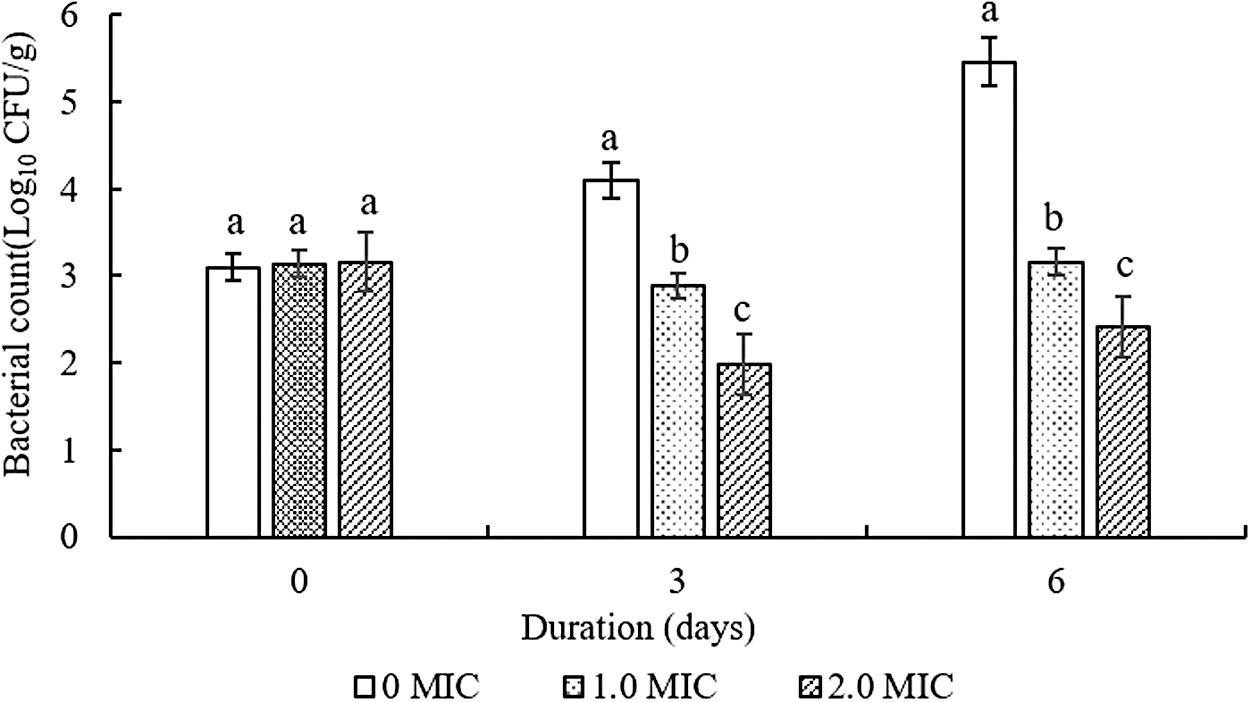
Inhibitory effect of CBCE on Salmonella Typhimurium ATCC 14028 in cooked chicken samples stored at 4°C for 6 days. Bars represent the standard deviation from triplicate experiments. Values with different letters are significantly different at p < 0.05. CBCE, Chrysanthemum buds' crude extract.
Color changes
As shown in Table 1, after treatments with CBCE for 0 and 3 days, no significant differences in L*, a*, b*, and ΔE* values were found between the treated and untreated groups (p > 0.05). After 6 days of storage, compared with the untreated group, the cooked chicken treated with CBCE showed higher L*and ΔE* values, and lower b* value (p < 0.05). No significant differences in a* values of samples with and without treatments with CBCE were detected during the entire storage period (p > 0.05). Besides, as the extension of storage time, the L* and a* values of untreated samples decreased significantly, and their b* value was significantly increased (p < 0.05), while, no significant changes in L*, a*, and b* values of cooked chicken treated with CBCE were found (p > 0.05).
The Color of Cooked Chicken Treated with Different Concentrations of Chrysanthemum Buds Crude Extract at 4°C for 0, 3, and 6 Days
Values represent means of three independent replicates ± standard deviation. Different lowercase superscripts (a–c) within the same row indicate significant differences between the values (p < 0.05). Different uppercase superscripts (A–C) within the same column indicate significant differences between the values (p < 0.05).
MIC, minimum inhibitory concentration.
Discussion
This study first revealed the antibacterial activity and mode of action of CBCE against Salmonella Typhimurium strains. Our results showed that CBCE exhibited a moderate antibacterial effect against Salmonella Typhimurium (MIC = 40 mg/mL) compared with aqueous garlic extract (48 mg/mL) and pepper extract (9.3 mg/mL) (Lee et al., 2015b; Kittisakulnam et al., 2017). In addition, in this study, the crude extract of Chrysanthemum buds was used to inhibit Salmonella Typhimurium, indicating that the purified Chrysanthemum buds' extract should have a better antibacterial effect.
In this study, the intracellular ATP concentration of Salmonella Typhimurium treated with CBCE decreased significantly, which is similar to the results as found in the process of Tetrapleura tetraptera stem extract against Salmonella Typhimurium (Agyemang et al., 2020). Meanwhile, some studies showed that the diminished ATP concentration may be related to the increased permeability of bacterial cell membranes after treatments with natural products, destruction of loss of K+ ions and inorganic phosphate, hydrolysis by the proton-pumping ATPase, and depletion of the intracellular ATP pool (Sánchez et al., 2010; Shi et al., 2016b; Khan et al., 2017).
Besides, our results indicated that after treatments with CBCE, the amounts of nucleic acid and protein leaking from Salmonella Typhimurium cells were increased significantly, which was similar to the previous studies (Guo et al., 2019; Li et al., 2019), and suggested that the membrane permeability of Salmonella Typhimurium increased after treatments with CBCE.
Membrane potential was an important indicator to reflect the vitality of bacterial cells. In this study, after treatments with CBCE, Salmonella Typhimurium caused the cell membrane depolarization, which is consistent with the changes in membrane potential of Salmonella Typhimurium treated by olive oil polyphenol extract, Staphylococcus aureus treated by cranberry anthocyanin, and Cronobacter sakazakii treated by CBCE (Guo et al., 2019; Chang et al., 2021; Gong et al., 2021).
As a contrast, the membrane hyperpolarization was found in the tested strains after exposure to citral, syringic acid, and white sagebrush methanolic extracts (Sánchez et al., 2010; Shi et al., 2016a, 2016b). In addition, some studies have found that the depolarization of cell membrane was related to the reduction of intracellular negative charges due to Na+ migration, while the hyperpolarization of cell membrane was caused by the K+ leaks or intracellular pH changes (Bot and Prodan, 2009; Casanova et al., 2017).
TEM images displayed that Salmonella Typhimurium cells were damaged by the CBCE treatments, such as separation of cell wall and membrane, cell collapse or even fragmentation, and cytoplasmic leakage. A similar phenomenon has been found in the inhibitory actions of olive oil polyphenols and sugar beet molasses polyphenols against foodborne pathogens (Chen et al., 2017; Fei et al., 2018, 2019; Guo et al., 2019).
In addition, Chen et al. (2017) concluded that polyphenols increase the cell membrane permeability and thus destroy cell morphology, phenolic acid can change the hydrophobicity of cell membrane and make the cell membrane break or form small pore, flavonoids form compounds with cell walls that disrupt cell function. All of the above compounds can be found in CBCE (Supplementary Table S1), and the corresponding phenomenon is also consistent with our study.
In previous studies, the L*, a*, and b* values of meat products treated with natural extracts changed significantly during the entire storage period, such as cooked pork treated with Perilla frutescens var. acuta water extract, cooked pork and beef treated with cranberry anthocyanin, and cooked pork treated with Amaranthus tricolor crude extract (Lee et al., 2015a; Guo et al., 2020; Gong et al., 2021). In our study, 80 mg/mL of CBCE has not significantly changed the L*, a*, and b* values of cooked chicken sample, and maintains the original color of cooked chicken throughout the storage period, which are possibly due to the faint yellow of the CBCE, which is very close to the color of cooked chicken, meanwhile, CBCE can maintain the color of cooked chicken due to its antioxidant effect.
In addition, although cooked chicken is rich in nutrients and bacteria can easily grow and multiply on it, the treatment with 80 mg/mL of CBCE can reduce about 1 Log CFU/g of Salmonella Typhimurium, indicating that treatment with 80 mg/mL is sufficient and effective. Therefore, 80 mg/mL was optimal to be used as preservative both in terms of antibacterial effect and in maintaining the original color of cooked chicken.
Conclusion
This study emphasizes the antibacterial effect of CBCE against Salmonella Typhimurium and the potential application in cooked chicken. The antibacterial effect of CBCE is achieved by reducing the concentration of intracellular ATP concentration, cell membrane depolarization, leakage of protein and nucleic acid, and destruction of cell morphology. Moreover, CBCE can effectively inhibit the growth of Salmonella Typhimurium in cooked chicken, and the original color of cooked chicken is not significantly changed throughout the storage period, indicating that CBCE is expected to be a preservative for chicken products. In future studies, we will focus on the dose optimization, safety assessment, and determination and purification of main antibacterial ingredients of CBCE.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported by the Natural Science Foundation of Henan Province (212300410137), Key Scientific Research Projects of Institutions of Higher Learning of Henan Province (21A550007), the National Natural Science Foundation of China (31601450), Key Science and Technology Program of Henan (212102110077), and Young Teacher Funding Program of the Henan Higher School (2020GGJS072).
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.